西藏青稞冬春性鑒定及抽穗期多樣性與穩定性分析
2019-11-13 10:41:02倫珠朗杰李慧慧郭剛剛其美旺姆高麗云唐亞偉尼瑪扎西達瓦頓珠
作物學報 2019年12期
倫珠朗杰 李慧慧 郭剛剛 其美旺姆 高麗云 唐亞偉 尼瑪扎西 達瓦頓珠,* 卓 嘎
西藏青稞冬春性鑒定及抽穗期多樣性與穩定性分析
倫珠朗杰1,2,3,4,5李慧慧5郭剛剛5其美旺姆2,3高麗云2,3唐亞偉2,3尼瑪扎西2,4達瓦頓珠2,3,*卓 嘎1,*
1西藏農牧學院, 西藏林芝 860000;2省部共建青稞和牦牛種質資源與遺傳改良國家重點實驗室, 西藏拉薩 850002;3西藏自治區農牧科學院農業研究所, 西藏拉薩 850002;4西藏自治區農牧科學院, 西藏拉薩 850002;5中國農業科學院作物科學研究所, 北京 100081
為提高青稞種質資源的利用效率并篩選優異雜交親本, 分別在西藏林芝和拉薩兩地的春播和秋播環境下對1605份青稞種質資源進行冬春性鑒定和抽穗期多樣性分析。設拉薩春播2個播期, 即正常春播I和晚10 d春播II, 以2個播期的抽穗期變化作為冬春性區別依據。結果表明, 96.2%的西藏青稞地方品種為春性, 在西藏3個生態區均有分布, 冬性品種僅有3.8%, 且主要分布在以林芝為主的藏東南生態區; 抽穗期的Shannon-Wiener’s多樣性分析表明, 春播條件下抽穗期多樣性高于秋播, 拉薩春播條件下抽穗期多樣性最高, 林芝春播次之, 林芝秋播最低; 在相同環境下, 地方品種的多樣性高于育成品種; 在環境穩定性分析中, 368份春性地方品種和21份育成品種在不同環境中抽穗期較為穩定, 其中康青3號在兩地所有參試品種中抽穗期穩定性最高。本研究為全面理解西藏青稞資源的冬春性、抽穗期多樣性和環境穩定性提供了參考, 為廣適應性青稞品種培育篩選出環境穩定性佳的親本材料。
青稞; 生長習性; 抽穗期; 多樣性; 穩定性
抽穗期是作物重要的馴化和農藝性狀, 抽穗期長短與作物產量和品質密切相關[1-4]。作物從起源中心向外傳播和馴化選擇時, 只有在恰當的時間抽穗, 才能使其正常生存和繁衍后代[5-6]。大麥存在3個抽穗期信號通路, 一是低溫感知, 即春化敏感性, 將大麥分為冬性和春性; 二是光周期響應通路, 長日照促進開花; 三是早熟相關基因調控通路, 分為早熟型和晚熟型[7-10]。前2個通路受外界環境影響, 環境與基因互作調控開花, 后者主要由自身遺傳決定[11]。目前, 通過圖位克隆或同源克隆手段, 已經分離鑒定出抽穗期的關鍵基因及基因家族, 如春化基因和光周期基因以及等, 這極大促進了大麥種質資源的鑒定與利用[12-16]。
青稞(裸大麥)是青藏高原地區藏民主要的糧食作物, 在西藏青稞播種面積和產量分別約占糧食總播種面積和產量的60%和55%, 同時也是農區飼草的主要來源[17-20]。青稞具有較好的適應性和抗逆性, 在農業生產和生態環境惡劣的西藏邊境農區或海拔4700 m以上的高寒農區, 青稞是唯一能正常收獲的糧食作物[20]。自1952年以來, 農業科研人員對西藏農作物種質資源多次考察和收集, 迄今共收集西藏大麥青稞種質資源7000余份, 大麥科技工作者對青稞種質進行了變種分類、多樣性評價等系統研究, 經田間種植、性狀觀察和鑒定編目后的3602份保存于國家農作物種質資源長期庫[21-23]。為青稞資源的遺傳研究和利用奠定了基礎。
西藏平均海拔在4000 m以上, 地理氣候生態環境多樣, 包括秋播區、冬春兼作區和春播區3個主要生態區[21], 到目前為止, 尚不明確不同來源的青稞種質在西藏各生態區的生長特性和環境適應性, 限制了多樣性地方品種在提高青稞育成品種的生態適應性方面的有效利用[24]。為此, 本研究利用1605份西藏青稞地方品種、育成品種及青海、四川等周邊藏區代表性育成品種, 在多種環境鑒定冬春性和抽穗期, 研究不同環境中青稞抽穗期的變化規律, 為青稞種質資源的利用與廣適應性青稞品種的選育提供依據。
1 材料與方法
1.1 試驗材料
本研究的青稞種質資源共1605份, 包括西藏地方品種1505份及育成品種70份、周邊藏區育成品種30份(青海19份、四川甘孜5份、甘肅甘南5份, 云南迪慶1份), 育成品種中有93份春性品種和7份冬性品種。以西藏主推春青稞品種藏青2000、藏青320、喜瑪拉19號作為春性品種對照, 冬青稞品種冬青18號和果洛作為冬性品種對照。地方品種覆蓋西藏所有生態區, 育成品種包括近50年選育的春青稞和冬青稞代表性品種。
1.2 試驗方法
2017年10月和2018年3月, 于西藏農牧學院植物科學院試驗基地(西藏林芝, 海拔2900 m, 94°34′E, 26°67′N)秋播和春播。在西藏農牧科學院農業研究所試驗地(西藏拉薩, 海拔3660 m, 91°04′E, 29°64′N)進行冬春性鑒定分2個播期, 春播I (當地正常播期, 2018年4月12日)和春播II (比正常播期晚10 d, 2018年4月22日)。每份材料種1行, 行長1.5 m, 每行25粒, 行距為30 cm。以每行材料中1/2的穗子抽出旗葉鞘一半的時間記為抽穗期[25-26]。參照Hemming等[27]的方法, 春播出苗后100 d作為區分冬春性的抽穗期記載期限。拉薩2個播期均正常抽穗的記為春性, 春播I能夠拔節或者極晚抽穗, 春播II不能抽穗, 甚至不能拔節的作為半冬性, 2個春播均不能抽穗的記為冬性[24,28](圖1)。

圖1 拉薩冬春性鑒定表型
1.3 數據分析
采用Microsoft Excel 2016和R語言整理和分析數據, 采用QTL IciMapping軟件中表型數據ANOVA功能分析遺傳力和相關性。利用Shannon-Wiener’s遺傳多樣性指數(Shannon-Wiener diversity index,)進行遺傳多樣性評價[29]。通過計算參試材料的平均值()和標準差(), 把抽穗期劃分為5等級, 早抽穗I (<–1.5)、偏早抽穗II (–1.5<<–0.5)、中間型III (–0.5<<+0.5)、偏晚抽穗IV (+0.5<<+1.5)和晚抽穗V (>+1.5),為每份材料的抽穗期。根據種質類型和生長習性, 將品種劃分為春性地方品種、春性育成品種、冬性地方品種和冬性育成品種4個類型。
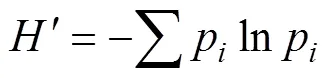
式中,為Shannon-Wiener’s遺傳多樣性指數,p為第級中的材料份數占該類型品種總份數的百分比。采用R語言multcomp包Tukey方法進行多重比較。
對不同環境和播期的抽穗期進行標準化, 以便不同等級之間的比較。按(–)/標準化, 式中為不同播期某一個參試材料的抽穗期,為播期內參試材料的平均值,為標準差。
2 結果與分析
2.1 青稞種質資源冬春性鑒定與地理分布
絕大多數青稞地方品種和育成品種均為春性類型(表1)。青稞地方品種中春性共有1448份, 冬性共有57份, 分別占地方品種的96.2%和3.8%; 春性育成品種共有93份與記載吻合; 冬性育成品種有7份, 在本研究中均表現為半冬性, 冬性育成品種中未發現強冬性類型。
春性是青稞資源的主要特征, 分布在西藏所有生態區, 冬性地方品種主要分布在林芝, 共30份; 其次在山南、昌都和日喀則, 分別有12、6和6份; 在拉薩只有3份材料。以牧業為主的阿里和那曲地區, 收集資源僅有41份, 未發現冬性類型(圖2)。目前生產上推廣的7份半冬性品種主要種植在拉薩、山南和林芝等海拔3700 m以下農區。
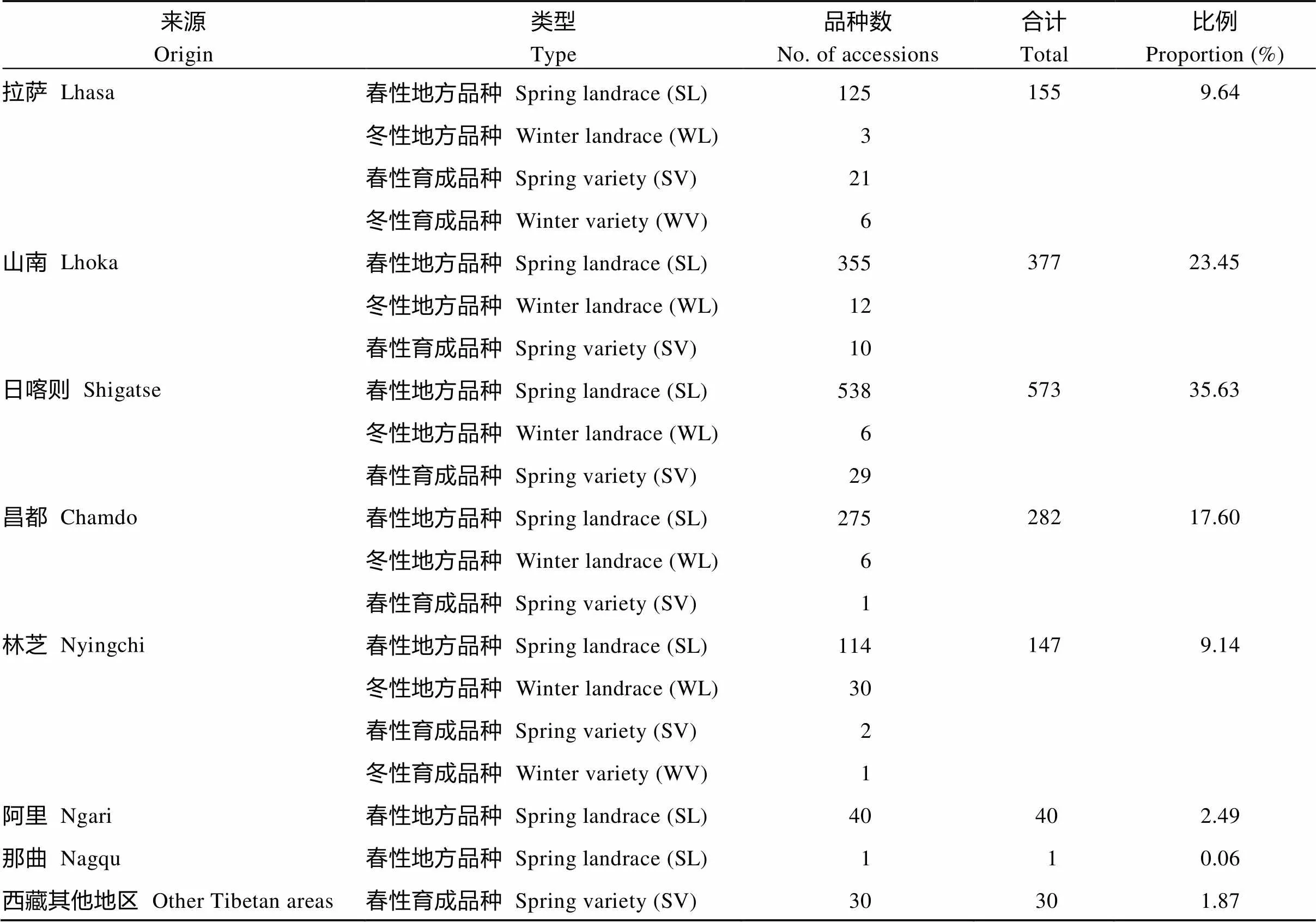
表1 材料來源及品種類型
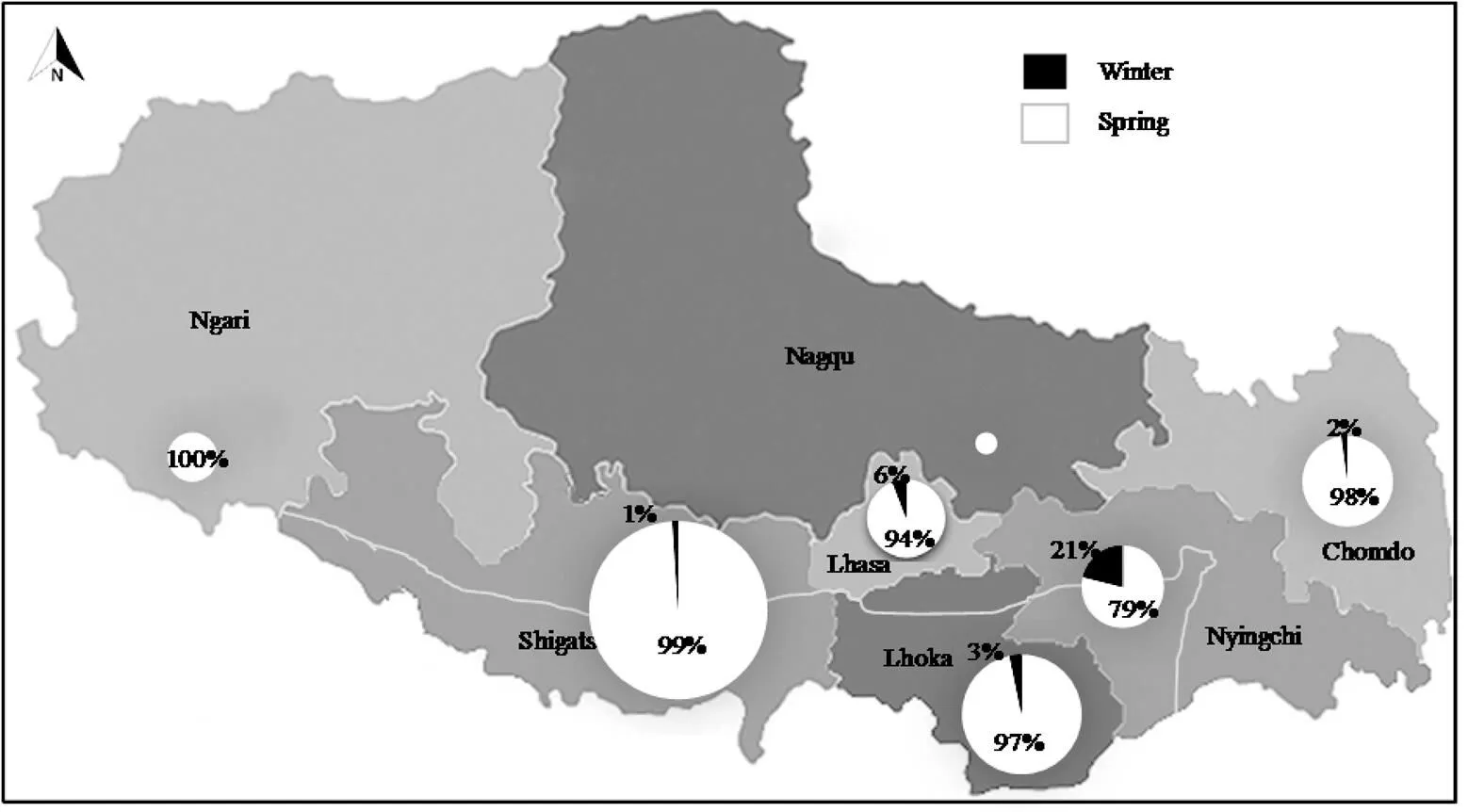
圖2 青稞冬春性資源的地理分布
灰色線條表示雅魯藏布江流域, 餅圖大小表示在總材料中所占的比重。
Gray line indicates the Yarlung Zangbo River basin, and the pie chart size represents the proportion to the entire material.
2.2 青稞種質資源抽穗期的多樣性
2.2.1 2017年林芝秋播抽穗期 林芝秋播條件下, 除116份春性地方品種和2份冬性地方品種不能正常越冬未獲得數據, 其余1487份材料均能正常抽穗。春性地方品種共有1332份, 平均抽穗期為174.54± 9.33 d, 其中40份(日喀則21份、山南10份、拉薩5份、昌都4份)抽穗期最早, 為160 d, 來源于日喀則的3份(2份謝通門縣、1份定結縣)抽穗期最晚, 為204 d。冬性地方品種共有55份, 平均抽穗期值為182.86± 10.04 d, 其中抽穗最早的有3份(林芝2份、日喀則1份), 抽穗期為161 d, 而來自西藏林芝的品種乃那(黑青稞)抽穗期最晚, 為202 d。
春性育成品種共有93份, 抽穗期平均值為174.40±6.68 d, 其中青海的2份(北青3號、北青4號)最早抽穗, 抽穗期為160 d, 林芝的紫芒抽穗期最晚, 為202 d。半冬性育成品種共有7份, 平均抽穗期為185.33±8.36 d, 冬青8號抽穗最早, 為180 d, 果洛抽穗期最晚, 為202 d。
2.2.2 2018年林芝春播 1605份青稞資源中除6份冬性地方品種、1份春性育成品種不能正常抽穗外,其余1598份材料在林芝春播均能抽穗。春性地方品種共有1448份, 抽穗期平均值為73.16±6.33 d, 其中來源西藏日喀則的2份抽穗期最早, 為59 d, 來源林芝的扎卓骨和山南隆子縣的變芒紫六棱抽穗期最晚,為94 d。冬性地方品種共有52份, 平均抽穗期為84.72±6.39 d, 其中來源拉薩的果洛抽穗期最早, 為64 d, 來源山南的勾芒白青稞的抽穗期最晚, 為97 d。
春性育成品種共有92份, 抽穗期平均值為75.78±5.50 d, 其中甘南的甘青1號抽穗期最早, 為62 d, 拉薩的拉薩勾芒、日喀則的喜瑪拉8號、喜瑪拉9號和喜瑪拉10號4個品種的抽穗期最晚, 為85 d。半冬性育成品種共有6份, 平均抽穗期值80.6±7.09 d,其中冬青1號抽穗期最早, 為71 d, 林芝的果洛抽穗期最晚, 為88 d。
2.2.3 2018年拉薩春播 1605份青稞資源中, 除56份冬性品種外, 其他均能正常抽穗。冬性品種中7份半冬性品種主莖能正常抽穗并記錄抽穗期。綜合兩個播期的抽穗期計算其廣義遺傳力(2), 為0.82。
1448份春性地方品種的抽穗期平均值為68.19± 5.62 d, 其中, 最早抽穗的來源于西藏日喀則的吉隆青稞, 抽穗期為53 d, 而來自西藏山南的左線那果(黑青稞)抽穗期最晚, 為85 d。冬性地方品種共有4份能夠正常抽穗, 抽穗期平均值為79.00±9.17 d, 其中來源西藏日喀則薩迦縣的紫青稞最早抽穗, 抽穗期為69 d, 而來自西藏山南市錯那縣的白青稞抽穗期最晚, 為87 d。春性育成品種共有93份, 抽穗期平均值為70.18±4.19d, 變幅為59~81 d; 其中, 北青7號抽穗期最早, 為59 d, 藏青7239號抽穗期最晚, 為81 d。4份半冬性育成品種冬青1號、冬青8號、冬青11號、冬青19號能夠記載抽穗期, 其平均值為73.75±3.28 d, 變幅為71~78 d。
在不同環境和播期條件下, 四種類型品種的抽穗期符合正態分布(圖3), 并且具有較高的多樣性。總體而言, 地方品種的多樣性高于育成品種, 多樣性最高的是在拉薩春播環境, 其次是林芝春播環境, 而林芝秋播環境下多樣性最低, 且春播環境的變異系數大于秋播環境(表2)。
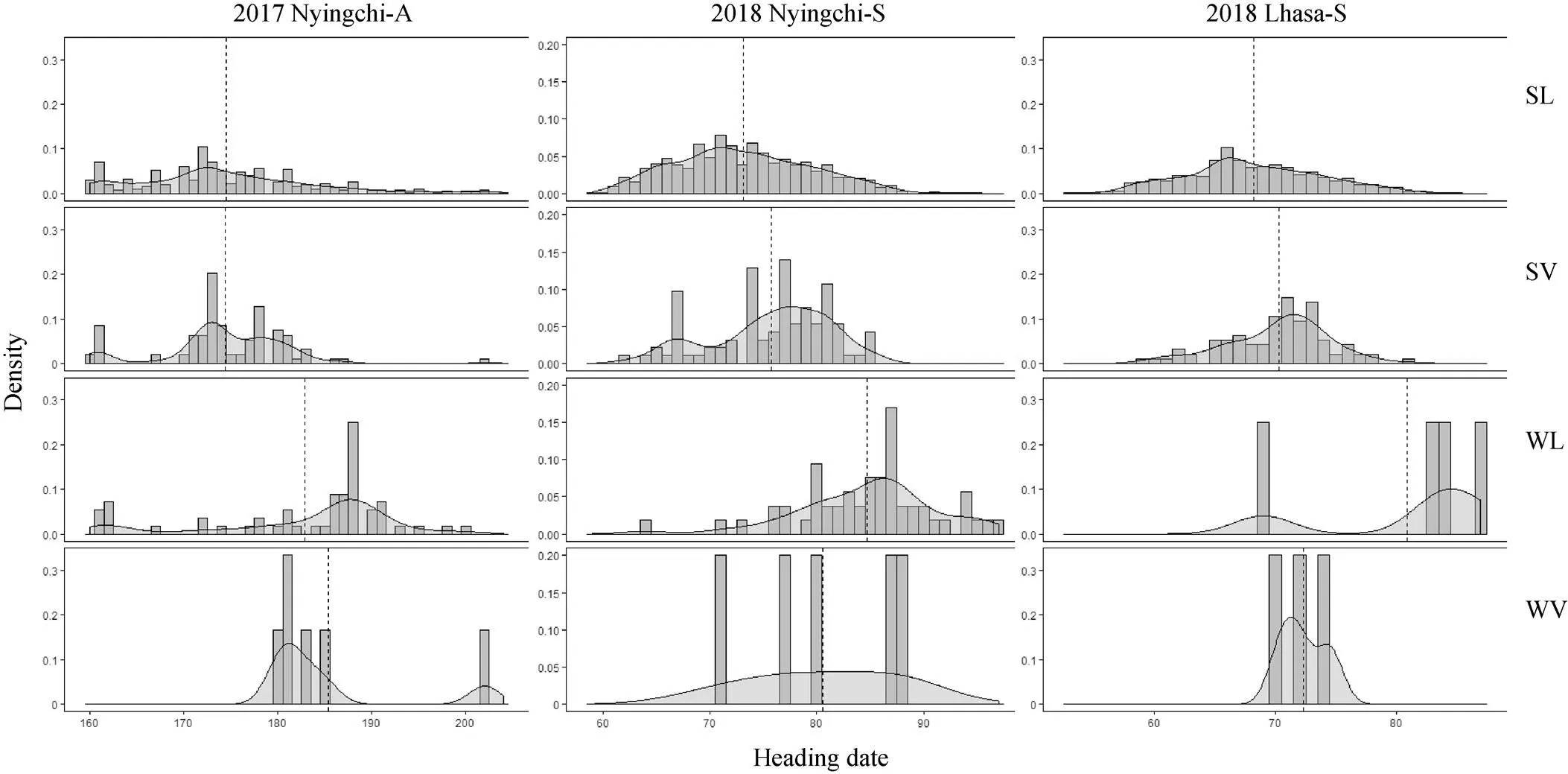
圖3 青稞資源在3個環境的抽穗期分布
黑色虛線表示抽穗期平均值, SL: 春性地方品種; SV: 春性育成品種; WL: 冬性地方品種; WV:冬性育成品種。2017年林芝秋播, 2018年林芝春播, 2018年拉薩春播。
Gray dotted line indicates the averaged value. SL:spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
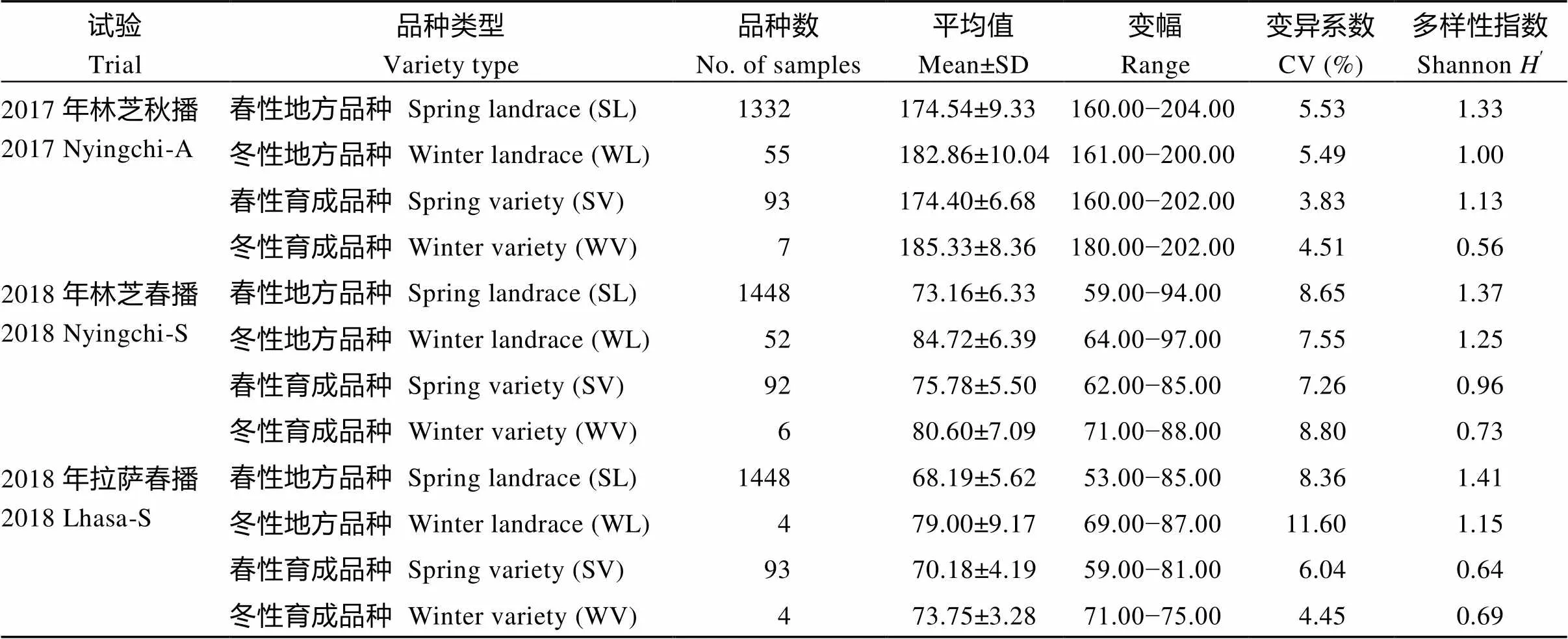
表2 青稞資源的抽穗期性狀變化及分布特征
2.3 青稞種質資源抽穗期的比較分析
2.3.1 相同播種條件下不同類型青稞資源抽穗期比較 在林芝秋播條件下, 春性品種較冬性品種早抽穗, 且達到極顯著水平(<0.01), 地方品種和育成品種中具有相同的趨勢。但生長習性相同的地方品種和育成品種之間無顯著差異。在林芝春播條件下, 春性地方品種最早抽穗, 且與其他3個類型品種的差異達到極顯著水平(<0.01), 其次是春性育成品種, 與冬性地方品種的差異達到顯著水平(<0.01), 與冬性育成品種無顯著差異, 冬性地方品種與冬性育成品種也沒有顯著差異。在拉薩春播條件下, 抽穗期由早到晚依次是春性地方品種、春性育成品種、冬性地方品種, 兩兩之間差異均達到極顯著水平(<0.01), 而冬性育成品種與其他三類品種之間沒有顯著差異(圖4)。
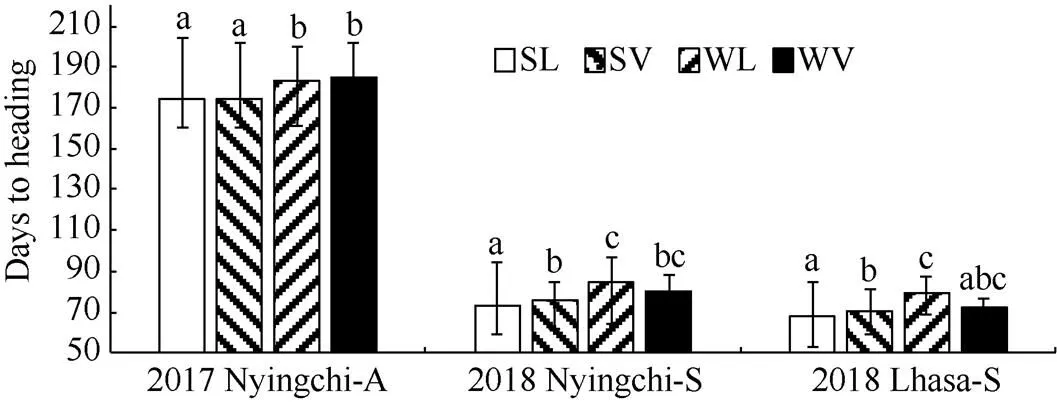
圖4 不同品種在秋播和春播條件下抽穗期的比較
SL: 春性地方品種; SV: 春性育成品種; WL: 冬性地方品種; WV: 冬性育成品種。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉薩春播。不同字母表示處理間差異顯著水平(< 0.01)。
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown. Significant difference (< 0.01).
2.3.2 不同環境條件下相同類型青稞資源抽穗期比較 在2個春播條件下, 地方品種在拉薩較在林芝更早抽穗, 且達到極顯著水平(<0.01), 春性品種和冬性品種存在相同的規律, 但是育成品種在2個春播條件下沒有顯著差異(圖5)。
2.4 抽穗期在不同環境中的相關性分析
3個環境抽穗期兩兩之間存在極顯著的正相關。其中, 2018年林芝和2018年拉薩2個春播之間相關系數為0.79, 明顯高于兩者與2017年林芝秋播之間的相關系數; 但2個春播與2017年秋播之間的相關系數很相近, 分別為0.40和0.43 (圖6)。
2.5 品種在抽穗期各等級中的分布
抽穗期主要集中在II~IV等級內, 即偏早抽穗、中間型和偏晚抽穗類型, 占2017年林芝秋播89.87%、2018年林芝春播87.13%和2018年拉薩春播的86.13%, I和V兩等級內所包含的品種量比較少, 只占10.13%、12.87%、13.87% (圖7)。
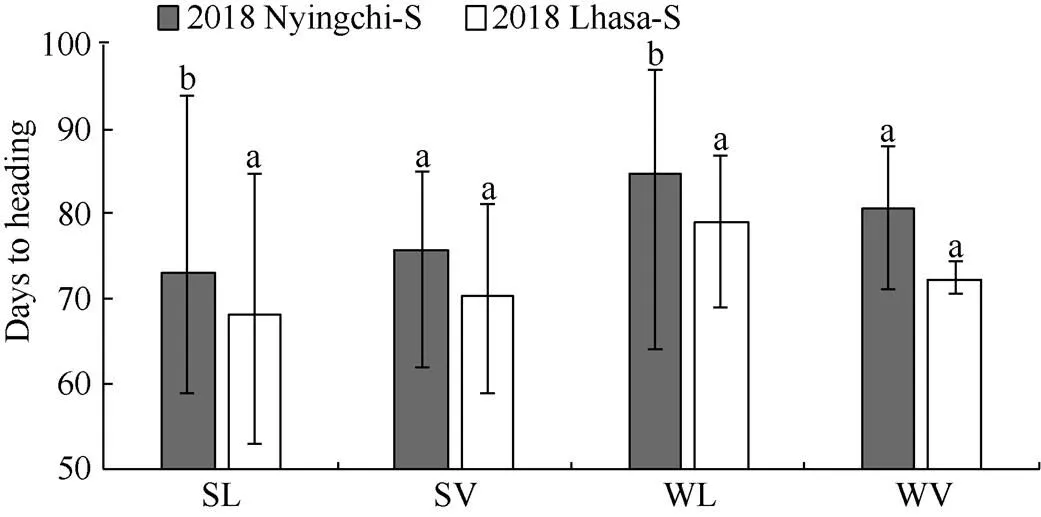
圖5 春播條件下不同環境間抽穗期的比較
SL: 春性地方品種; SV: 春性育成品種; WL: 冬性地方品種; WV:冬性育成品種。2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉薩春播。不同字母表示處理間差異顯著水平(<0.01)。
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown. Significant difference (<0.01).
2.6 青稞種質資源不同環境之間的穩定性分析
2.6.1 秋播和春播抽穗期穩定性分析 青稞資源在所有條件下同一個等級內共享或穩定性較高的品種共有390份品種, 分別為I(3)、II(123)、III(199)、IV(55)、V(10); 其中, 春性地方品種中穩定性較高的品種共有368份品種, 分別為I(3)、II(122)、III(185)、IV(48)、V(10); 春性育成品種中穩定性較高的品種共有21份, 分別為II(1)、III(14)、IV(6); 冬性地方品種和冬性育成品種中只有冬青11號在所有環境中穩定在IV, 冬性地方品種無早抽穗類型(I), 冬性育成品種中未發現早抽穗類型(I)和偏早抽穗類型(II) (圖8)。
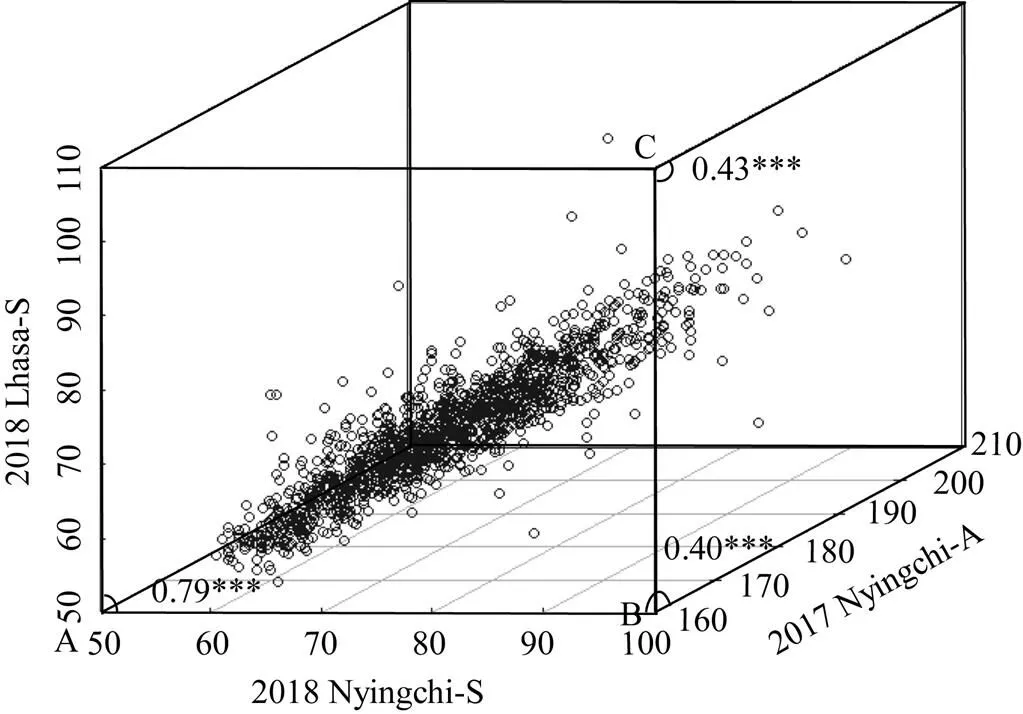
圖6 不同播期和環境間抽穗期的相關性
A、B、C表示播期間或環境間的相關系數。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉薩春播。
A, B, C represents the correlation coefficient of sowing time or environments. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
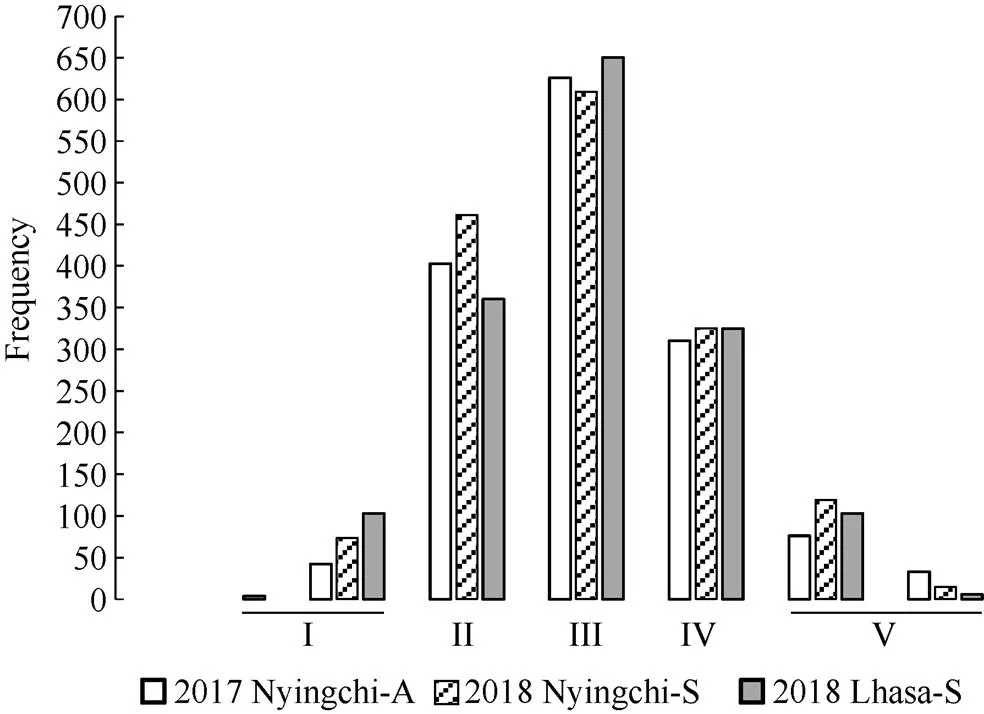
圖7 不同播期抽穗期值標準化的五個等級的變化頻率
I: 早抽穗; II: 偏早抽穗; III:中間型; IV: 偏晚抽穗; V: 晚抽穗。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉薩春播。
I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.

圖8 不同品種在不同播期和環境條件下抽穗期的穩定性分析
SL: 春性地方品種; SV: 春性育成品種; WL: 冬性地方品種; WV: 冬性育成品種。I: 早抽穗; II: 偏早抽穗; III: 中間型; IV: 偏晚抽穗; V: 晚抽穗。a: 2017年林芝秋播; b: 2018年林芝春播; c: 2018年拉薩春播。紅色數字表示3個環境內共有的品種數。
SL: spring landrace; SV: spring variety; WL: winter landrace; WV: winter variety. I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. a: 2017 Nyingchi-Autumn sown; b: 2018 Nyingchi-Spring sown; c: 2018 Lhasa-Spring sown. Red number indicates samples shared by the three environments.
2.6.2 青稞育成品種的穩定性評價 為評價育成品種在不同生態環境的穩定性, 選擇主推青稞品種藏青320、藏青2000、喜瑪拉19號、喜瑪拉22號和早期青稞育種的骨干親本昆侖1號等代表性青稞育成品種進行環境穩定性分析。藏青320和康青3號穩定性最高, 3個環境下穩定在III等級中, 其次是藏青2000、高原早1號、喜瑪拉19號、喜瑪拉22號、昆侖1號等品種。所有育成品種中喜瑪拉9號、白玉紫芒和藏青148三個品種的環境穩定性最差(圖9)。
3 討論
國家農作物種質庫保存編目的3602份青稞地方品種中, 日喀則、山南、昌都、林芝和拉薩的資源占98%, 阿里和那曲的資源僅占2%[21]。本研究中, 地理來源和基礎信息相似的材料只選代表性品種, 共選取1505份地方品種。育成品種包含自1966年開展青稞育種以來, 每個階段具有一定推廣面積和利用時間的代表性品種。用于本研究的實驗材料不僅覆蓋西藏所有生態區, 四省區藏區的育成品種也選自曾經或者近年多次用來改良西藏青稞的骨干品種,能夠全面反映青稞種質資源的整體情況。
鑒定結果表明, 青稞地方品種絕大多數是春性類型, 這與青稞種植實際相吻合。在西藏, 生產上一直以春青稞為主, 冬青稞只在邊境地區和低海拔極小面積的盆地種植, 年總面積只有267~333 hm2 [17-18]。近幾年隨著西藏種植業結構調整和冬青稞復種飼草的巨大優勢, 冬青稞面積逐漸擴大, 年播種面積穩定在3333 hm2以上, 但是現有的冬青稞品種很難在海拔3700 m以上的高寒農區推廣[21]。一般情況下, 品種冬性與苗期抗寒性正相關[8]。本研究發現生產上主推的冬青稞育成品種均為半冬性類型, 這是限制現有冬青稞品種在高海拔地區大面積推廣的主要因素之一。除西藏昌都和林芝有較少的冬青稞資源外, 在其他生態區冬青稞資源極為稀少。因此, 除有效利用西藏當地的強冬性資源外, 引進優異抗寒冬大麥種質以豐富和拓寬冬青稞遺傳基礎是未來冬青稞育種的有效途徑。
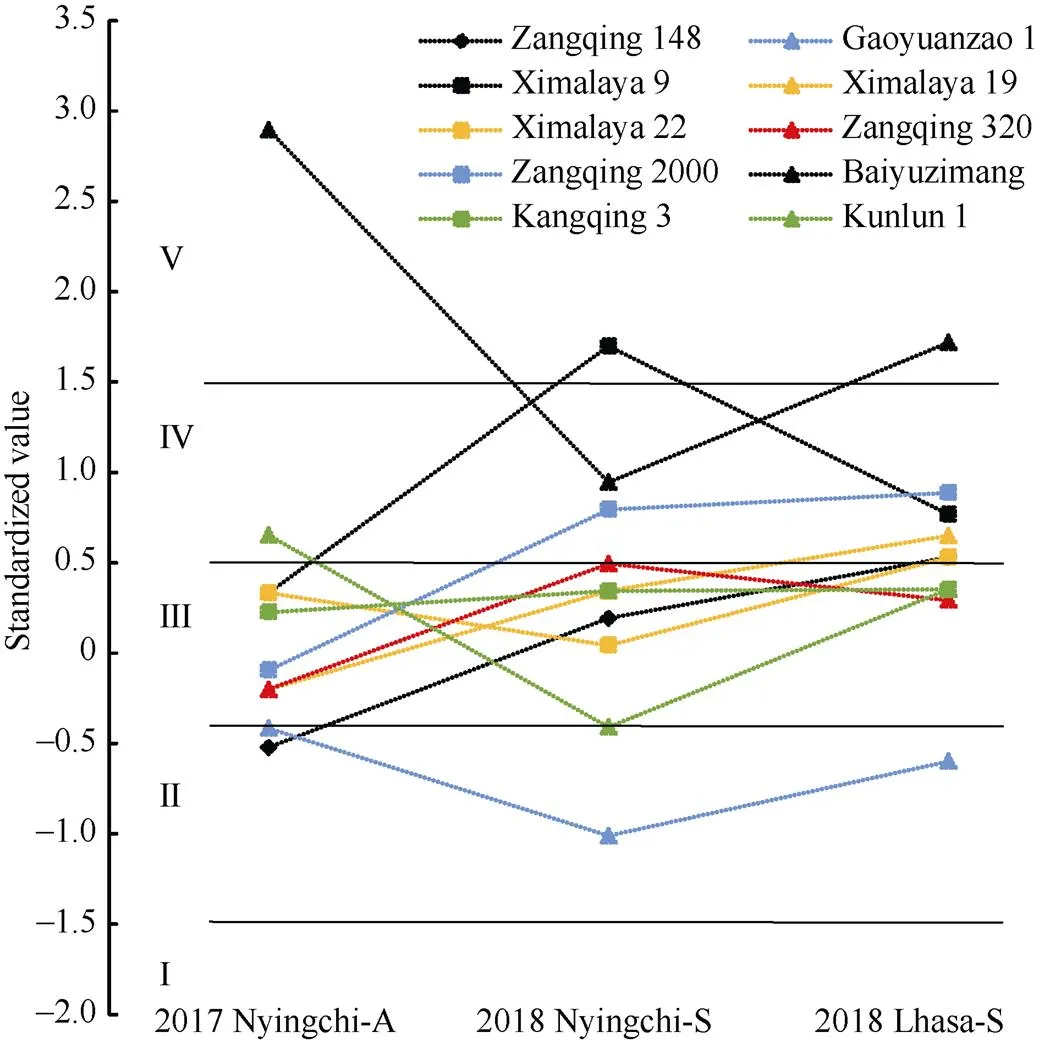
圖9 青稞主要育成品種在不同播期和環境之間抽穗期的穩定性分析
I: 早抽穗; II: 偏早抽穗; III: 中間型; IV: 偏晚抽穗; V: 晚抽穗。2017 Nyingchi-A: 2017年林芝秋播; 2018 Nyingchi-S: 2018年林芝春播; 2018 Lhasa-S: 2018年拉薩春播。
I: early heading; II: partial early heading; III: middle heading; IV: partial late heading; V: late heading. 2017 Nyingchi-A: 2017 Nyingchi-Autumn sown; 2018 Nyingchi-S: 2018 Nyingchi-Spring sown; 2018 Lhasa-S: 2018 Lhasa-Spring sown.
本研究發現, 冬春性是影響青稞抽穗期的最主要因素, 在所有環境中, 地方品種和育成品種的冬性類型成熟均明顯晚于春性類型(圖4)。藏東南的生態區普遍存在春性品種秋播的現象, 這些區域的資源也往往當作冬性類型, 這種混淆大幅降低了種質資源的利用效率。因此, 冬春性的精準鑒定是有效利用青稞資源的前提。本研究鑒定的冬春性與馬得泉[21]的研究結果基本一致, 但由于20世紀80年代青稞資源收集入庫編目時以種植區域和種植習慣作為判斷冬春性的主要依據, 即秋播方式的春性材料記為冬性, 因此, 本研究也是對青稞資源目錄性狀的補充鑒定。在所有環境中, 冬性育成品種和冬性地方品種之間抽穗期無明顯差異, 且綜合農藝性狀與冬青稞骨干親本“果洛”類似, 可能與冬青稞地方品種數目相對少且均源于西藏東南部有關, 但是對于這些冬青稞種質資源的遺傳相似性有待進一步研究。
根據參試品種在不同播期或環境條件下抽穗期的變化規律, 有些品種在環境之間的波動較小, 而有些卻很大。對于波動較小的品種, 環境的改變不會引起表型上大的變化, 其說這些品種對環境的改變不太敏感, 它們的環境穩定性較高。而對于波動較大的品種, 環境的改變會引起表型上較大的變化, 它們對環境的改變比較敏感, 其環境穩定性較低。因此, 根據多個品種在多種環境下的表型數據, 就能對品種的環境敏感性進行評價, 又稱為穩定性分析(stability analysis)[30]。本研究通過抽穗期的標準差倍數對資源分類, 共分I~V 5個等級, 發現絕大多數地方品種和育成品種分布在偏早抽穗類型(II)、中間型(III)和偏晚抽穗類型中(IV) (圖8)。品種穩定性分析發現藏青320在西藏育成品種中穩定性最高(圖9),在青稞生產實踐中藏青320具有廣適應性和穩定性而種植了近40年。林芝波密縣白玉村的紫芒, 因當地表現良好而被選為骨干親本, 早期推廣到其他生態區, 配置了大量雜交組合。但是, 在其他地區并沒有達到預期的效果, 未能成為優勢品種。另外, 90年代審定的藏青148示范數年后因綜合表現不穩定而未能大面積推廣。本研究品種穩定性分析也顯示, 白玉紫芒和藏青148穩定最差(圖9)。因此, 品種穩定性能夠較為準確地反映品種在實際生產中的推廣情況和利用潛力。
作物品種的抽穗期是最重要的農藝性狀之一, 基于抽穗期的穩定性與育成品種的生長情況完全吻合, 即穩定性與推廣面積和年限正相關。因此, 廣適應性青稞品種的選育應注重參考品種抽穗期的穩定性。I~V 5個等級地方品種中均存在抽穗期穩定性高的材料, 來源青海的昆侖1號是早期(1974—1986年)西藏青稞育種的一個骨干親本[31]。本研究中來源于四川甘孜的康青3號是所有參試材料中穩定性最好的品種, 其穩定性顯著高于昆侖1號, 且綜合性狀優良, 可作為今后青稞改良的候選骨干親本。
4 結論
生長習性和抽穗期是決定青稞生態適應性的重要因素。青稞資源的抽穗期具有豐富的變異, 多樣性分析和不同環境穩定性評價為高穩定性的青稞種的篩選及高產、廣適應性春冬青稞品種的選育提供了理論依據。
致謝:感謝中國農業科學院作物科學研究所小宗作物種質資源創新研究者張京研究員提供青稞種質, 數量遺傳創新研究組王建康研究員指導數據分析。
[1] Pourkheirandish M, Komatsuda T. The importance of barley genetics and domestication in a global perspective., 2007, 100: 999–1008.
[2] Turner A, Beales J, Faure S, Dunford R P, Laurie D A. The pseudo-response regulatorprovides adaptation to photoperiod in barley., 2005, 310: 1031–1034.
[3] Jones H, Leigh F J, Mackay I, Bower M A, Smith L M, Charles M P, Jones G, Jones M K, Brown T A, Powell W. Population based resequencing reveals that the flowering time adaptation of cultivated barley originated east of the Fertile Crescent., 2008, 25: 2211–2219.
[4] Ibrahim A, Harrison M, Meinke H, Fan Y, Johnson P, Zhou M. A regulator of early flowering in barley (L.)., 2018, 13: e0200722.
[5] Doebley J F, Gaut B S, Smith B D. The molecular genetics of crop domestication., 2006, 127: 1309–1321.
[6] Sang T. Genes and mutations underlying domestication transitions in grasses., 2009, 149: 63–70.
[7] Trevaskis B, Bagnall D J, Ellis M H, Peacock W J, Dennis E S. MADS box genes control vernalization-induced flowering in cereals., 2003, 100: 13099–13104.
[8] Saisho D, Ishii M, Hori K, Sato K. Natural variation of barley vernalization requirements: implication of quantitative variation of winter growth habit as an adaptive trait in East Asia., 2011, 52: 775–784.
[9] Nishida H, Yoshida T, Kawakami K, Fujita M, Long B, Akashi Y, Laurie D A, Kato K. Structural variation in the 5′ upstream region of photoperiod-insensitive allelesa anda identified in hexaploid wheat (L.), and their effect on heading time., 2012, 31: 27–37.
[10] Comadran J, Kilian B, Russell J, Ramsay L, Stein N, Ganal M, Shaw P, Bayer M, Thomas W, Marshall D, Hedley P, Tondelli A, Pecchioni N, Francia E, Korzun V, Walther A, Waugh R. Natural variation in a homolog of Antirrhinumcontributed to spring growth habit and environmental adaptation in cultivated barley., 2012, 44: 1388–1392.
[11] Laurie D A, Pratchett N, Bezant J H, Snape J W. RFLP mapping of five major genes and eight quantitative trait loci controlling flowering time in a winter × spring barley (L.) cross., 1995, 38: 575–585.
[12] Yan L L, Loukoianov A, Tranquilli G, Helguera M, Fahima T, Dubcovsky J. Positional cloning of the wheat vernalization gene., 2003, 100: 6263–6268.
[13] Yan L L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W, Sanmiguel P, Bennetzen J L, Echenique V, Dubcovsky J. The wheatgene is a flowering repressor down-regulated by vernalization., 2004, 303: 1640–1644.
[14] Yan L L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A, Valarik M, Yasuda S, Dubcovsky J. The wheat and barley vernalization geneis an orthologue of., 2006, 103: 19581–19586.
[15] Faure S, Higgins J, Turner A, Laurie D A. The FLOWERING LOCUS T-like gene family in barley ()., 2007, 176: 599–609.
[16] Distelfeld A, Li C, Dubcovsky J. Regulation of flowering in temperate cereals., 2009, 12: 178–184.
[17] 盧良恕. 中國大麥學. 北京: 中國農業出版社, 1996. Lu L S. Chinese Barley. Beijing: China Science and Technology Press, 1996. pp 3–7 (in Chinese).
[18] 胡頌杰. 西藏農業概論. 成都: 四川科學技術出版社, 1995. Hu S J. An Introduction of Tibet Agriculture. Chengdu: Sichuan Science and Technology Press, 1995 (in Chinese)
[19] Tashi N. Food preparation from hull-less barley in Tibet. In: Food Barley—Importance, Uses and Local Knowledge: Proc. International Workshop on Food Barley Improvement, 2002. Grando S, Macpherson H G, eds. ICARDA, Aleppo, Syria, 2005.
[20] 朱睦元, 張京. 大麥(青稞)營養分析及其食品加工. 杭州: 浙江大學出版社, 2015. Zhu M Y, Zhang J. Barley Nutritional Analysis and Food Processing. Hangzhou: Zhejiang University Press, 2015 (in Chinese).
[21] 馬得泉. 中國西藏大麥遺傳資源. 北京: 中國農業出版社, 2000. Ma D Q. Genetic Resources of Tibetan Barley in China. Beijing: China Science and Technology Press, 2000 (in Chinese).
[22] 張京, 曹永生. 我國大麥基因庫的群體結構和表型多樣性研究. 中國農業科學, 1998, 32(4): 20–26. Zhang J, Cao Y S. Population Structure and Phenotypic Diversity of Barley Gene Bank in China., 1998 (in Chinese with English abstract).
[23] 馮宗云. 徐廷文大麥學術文集. 成都四川科學技術出版社, 2006. Feng Z Y. Xu Tingwen,s Work for Barley Science. Chengdu: Sichuan Science and Technology Press, 2006 (in Chinese)
[24] 達瓦頓珠. 中國大麥低溫春化和光周期基因單倍型及表型關聯分析. 中國農業科學院研究生院博士學位論文, 北京, 2015. pp 25–28.Dawadondrop. Haplotypes and Phenotypic Association Analsis of Vernalization and Photoperiod Genes in Chinese Barley in china. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2015. pp 25–28 (in Chinese with English abstract).
[25] Alqudah A M, Sharma R, Pasam R K, Graner A, Kilian B, Schnurbusch T. Genetic dissection of photoperiod response based on GWAS of pre-anthesis phase duration in spring barley., 2014, 9: e113120.
[26] Alqudah A M, Schnurbusch T. Heading date is not flowering time in spring barley., 2017, 8: 896.
[27] Hemming M N, Fieg S, Peacock W J, Dennis E S, Trevaskis B. Regions associated with repression of the barley ()gene are not required for cold induction., 2009, 282: 107–117.
[28] Dondup D, Dong G Q, Xu D D, Zhang L S, Zha S, Yuan X M, Tashi N, Zhang J, Guo G G. Allelic variation and geographic distribution of vernalization genesandin Chinese barley germplasm., 2016, 36: 11
[29] Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinons Press, 1949.
[30] 王建康. 數量遺傳學. 北京: 科學出版社, 2017. Wang J K. Quantitative Genetics. Beijing: Science Press, 2017 (in Chinese).
[31] 強小林. 西藏青稞品種系譜及其分析. 中國作物學會大麥專業委員會. 中國大麥文集. 北京: 中國農業科技出版社, 1986. pp 189–195. Qiang X L. The pedigree and its analysis of barley varieties in Tibet. In: Barley Professional Committee of China Crop Society. Barley Science in China. Beijing: China Agricultural Science and Technology Press, 1986. pp 189–195 (in Chinese).
Growth habit identification and diversity and stability analysis of heading date in Tibetan barley (L)
Lhundrupnamgyal1,2,3,4,5, LI Hui-Hui5, GUO Gang-Gang5, Chemiwangmo2,3, GAO Li-Yun2,3, TANG Ya-Wei2,3, Nyematashi2,4, Dawadondrup2,3,*, and Dolkar1,*
1Tibet Agriculture and Animal Husbandry College, Linzhi, Tibet 860000, China;2State Key Laboratory of Hulless Barley and Yak Germplasm Resources and Genetic Improvement, Lhasa, Tibet 850002, China;3Institute of Agricultural Sciences, Tibet Academy of Agricultural and Animal Husbandry Science, Lhasa, Tibet 850002, China;4Tibet Academy of Agricultural and Animal Husbandry Science, Lhasa, Tibet 850002, China;5Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
To improve the utilization of Tibetan barley germplasm and select appropriate parents in hybridization breeding, evaluated the growth habit and heading date diversity of 1605 Tibetan barley germplasm under the spring and autumn sowing conditions in Nyingchi and Lhasa, Tibet. The growth habit was determined by the difference in heading date observed in two spring sowing dates in Lhasa, i.e. normal spring sowing I and 10 days later of spring sowing II, showing that 96.2% of the landraces were spring barley distributed in all three ecological regions; 3.8% of the landraces were the winter barley accessions originated mainly from the southeastern ecological region of Tibet. Shannon-Wiener’s analysis showed that the heading date diversity was higher under spring-sown environment than under autumn-sown environment. The heading date diversity was highest in Lhasa region with spring sowing, moderate in Nyingchi spring-sown environment, and lowest in Nyingchi autumn-sown environment. In the same environment, landraces had higher diversity than modern varieties. A total of 368 spring-type landraces and 21 varieties were stable in heading date diversity across environments, and the cultivar Kangqing 3 had the highest stability among all tested materials. This study helps to further understand the growth habits, heading dates diversity and environmental stability of barley resources in Tibet, and provides useful information for the development of broad-adapted barley varieties.
barley; growth habit; heading date diversity; stability
本研究由國家自然科學基金項目(31660299), 西藏自治區重大專項(XZ201801NA01)和西藏農牧學院研究生創新計劃資助項目YJS2017-07(502000114)資助。
This study was supported by the National Natural Science Foundation of China (31660299), the Tibet Department of Major Projects (XZ201801NA01), and the Tibet Agricultural and Animal Husbandry College Graduate Innovation Program funded project YJS2017-07 (502000114).
達瓦頓珠, E-mail: dwdunzhu@126.com; 卓嘎, E-mail: slzhuoga@163.com
E-mail: lhundrupnamgyal@163.com
2019-03-20;
2019-06-22;
2019-07-15.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20190712.1520.004.html
10.3724/SP.J.1006.2019.91025
