油用牡丹‘鳳丹'種子內源抑制物研究
2019-10-23 07:51:52朱銘瑋鄒雨婷李永榮李淑嫻
西南林業大學學報 2019年6期
朱銘瑋 鄒雨婷 李永榮 李淑嫻
( 1. 南京林業大學南方現代林業協同創新中心,江蘇 南京 210037;2. 南京綠宙薄殼山核桃科技有限公司,江蘇 南京 210014)
牡丹(Paeonia suffruticosa)是毛茛科(Ranunculaceae)、芍藥屬(Paeonia)多年生落葉灌木,在我國分布范圍較廣。油用牡丹是牡丹組中結實力強,能夠生產種子及加工食用牡丹籽油的一個特殊類型。2011年3月以牡丹籽仁為原料提煉出的牡丹籽油被衛生部批準為新資源食品[1]。對牡丹籽油的成分進行分析后,人們發現其主要以亞麻酸、油酸、亞油酸、棕櫚酸和硬脂酸為主的37種成分組成,其中不飽和脂肪酸占總量83.42%[2],特別籽油中α-亞麻酸含量較高,是橄欖油的140倍[3],因此牡丹籽是一種適宜開發利用的良好油料資源,具有廣闊的發展前景。
生產上油用牡丹多用種子進行繁殖,但是該種子存在休眠習性,需要經過6個月以上的層積才能正常萌發,這給播種繁殖帶來了很多不便。以往的研究表明,造成鳳丹(P. ostill‘Fengdan')種子休眠的原因是種胚形態發育不完全以及種子內存在抑制物引起的生理休眠,是綜合休眠[4]。宋會興等[5]認為四川牡丹種子(P. decomposita)的種皮、胚乳中含有抑制白菜種子萌發及幼苗生長的物質,并且隨著該物質濃度的增加,抑制作用更為顯著,但是對造成其休眠的抑制物的種類卻未開展研究。本研究以油用牡丹的主栽品種群——鳳丹種子為材料,從內源抑制物質角度討論其休眠原因,通過系統溶劑分離法對鳳丹種皮和種胚的浸提液進行分離,并對各提取相進行生物測定,結合氣相色譜-質譜聯用(GC-MS)技術分析造成鳳丹種子休眠的內源抑制物的成分,探討內源抑制物與種子休眠的關系,以期為種子生物學及休眠機理研究提供參考。
1 材料與方法
1.1 實驗材料
油用牡丹‘鳳丹'種子,從安徽亳州采后置于5 ℃冰箱儲藏;市售白菜種子(矮腳黃),純度≥95.0%,發芽率≥85%。
1.2 實驗方法
1.2.1 鳳丹種子內源抑制物的提取
稱取飽滿鳳丹種子50 g,將其分為種皮、種胚2個部分,用粉碎機磨碎后,置于500 mL錐形瓶中,再加入400 mL的80%甲醇溶液,于4 ℃條件下密閉浸提,其間多次取出攪拌,使浸提充分。24 h后過濾浸提液,再向錐形瓶中加入400 mL的甲醇溶液,連續浸提3次,最后將3次浸提液合并。在60 ℃下減壓蒸除甲醇和部分水,最后將濃縮液定容至200 mL。
1.2.2 鳳丹種子內源抑制物的初步分離
采用系統溶劑分離法[6]對上述種子各部位的甲醇提取液進行初步分離,將得到的石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相浸提液,分別置于旋轉蒸發儀上進行濃縮蒸發,然后定容為200 mL,置于4 ℃冰箱中保存。
1.2.3 鳳丹種子分離相的生物活性測定
以白菜種籽為材料,對上述各相提取的溶液進行生物測定,具體為:取各相浸提液5 mL,分別加入鋪有一層濾紙的培養皿中,待有機溶劑揮發后,加5 mL蒸餾水,對照加入相同體積的蒸餾水。隨機數取白菜籽50粒,3次重復,置于培養皿中,隨后將培養皿放在25 ℃的恒溫光照培養箱內進行發芽實驗,當胚根伸長至1 cm時即認為該粒種子已經發芽,每8 h統計1次,連續3 d種子發芽粒數不超過1%時即認為發芽結束,最后計算各處理的發芽率。
1.2.4 種皮、種胚分離相的GC-MS鑒定
分別取鳳丹種子的種皮、種胚各有機相濃縮液100 mL,在旋轉蒸發儀上減壓濃縮蒸干,得到濃縮的干物質,然后再用相應的有機溶劑溶解定容至3 mL,即得到各有機相提取物的濃縮樣品。上述濃縮樣品在清華大學分析中心質譜室進行鑒定,鑒定條件如下:氣質儀器:GC-MS QP2010 ultra(Shimadzu,Kyoto,Japan)。氣質分析方法:柱子 Rxi-5Sil MS(Restek,USA,30 m×0.25 mm×0.25 μm),進樣口300 ℃,分流進樣分流比20∶1,載氣為高純氦氣,流速為1 mL/min。升溫程序:50 ℃保持 5 min,以 5 ℃/min漲至 300 ℃保持30 min。質譜條件:離子源:EI;離子源溫度:200 ℃;接口溫度:250 ℃,溶劑切除時間:3 min;掃描45~500 m/z。將GC-MS鑒定出的各個成分的質譜圖與標準庫譜圖進行核對,確定各個物質的名稱、分子式、分子量和相對含量等。
2 結果與分析
2.1 種皮和種胚各分離相的生物測定
鳳丹種子的種皮、種胚各相提取液對白菜籽發芽的影響見表1。從表1可知:種子各部位、各相提取液對白菜籽發芽均有一定的抑制作用,且除石油醚相及種胚乙酸乙酯相外,其他各相均使白菜籽發芽顯著下降,但種子部位不同、提取相不同,其對白菜籽發芽率的抑制程度有所不同。種皮提取液中,乙醚相的抑制作用最強,處理后白菜籽幾乎不能發芽,發芽率由對照的86.7%下降到0.7%;其次是乙酸乙酯相,發芽率下降到6.7%,降低了80.0%;甲醇相、水相處理后,發芽率也有較大程度的下降,分別較對照下降了52.0%、37.4%。種胚部位,仍是乙醚相的抑制作用最強,使白菜籽發芽率顯著下降到3.3%,甲醇相、水相處理后,白菜籽發芽率分別顯著下降到44.7%、56.0%,乙酸乙酯相處理后白菜籽的發芽率為83.4%,較對照差異不顯著。同一提取相中,不同部位對白菜籽發芽的影響程度也不盡相同,種皮乙酸乙酯相和甲醇相的抑制作用顯著高于種胚,特別是乙酸乙酯相,種皮提取液處理后僅有6.7%的白菜籽能夠發芽,但種胚的提取液卻使白菜籽發芽率下降不顯著;乙醚相、水相中,種皮、種胚部位的抑制作用差異不顯著;石油醚相中則是種胚的抑制作用略強于種皮,但二者均較對照差異不顯著。總的來說,同一提取相,種皮部位的抑制作用略強于種胚。種子不同部位各相生物測定的發芽率結果,均是乙醚相的抑制作用最強,因此鳳丹種子的內源抑制物可能主要存在于乙醚相中。
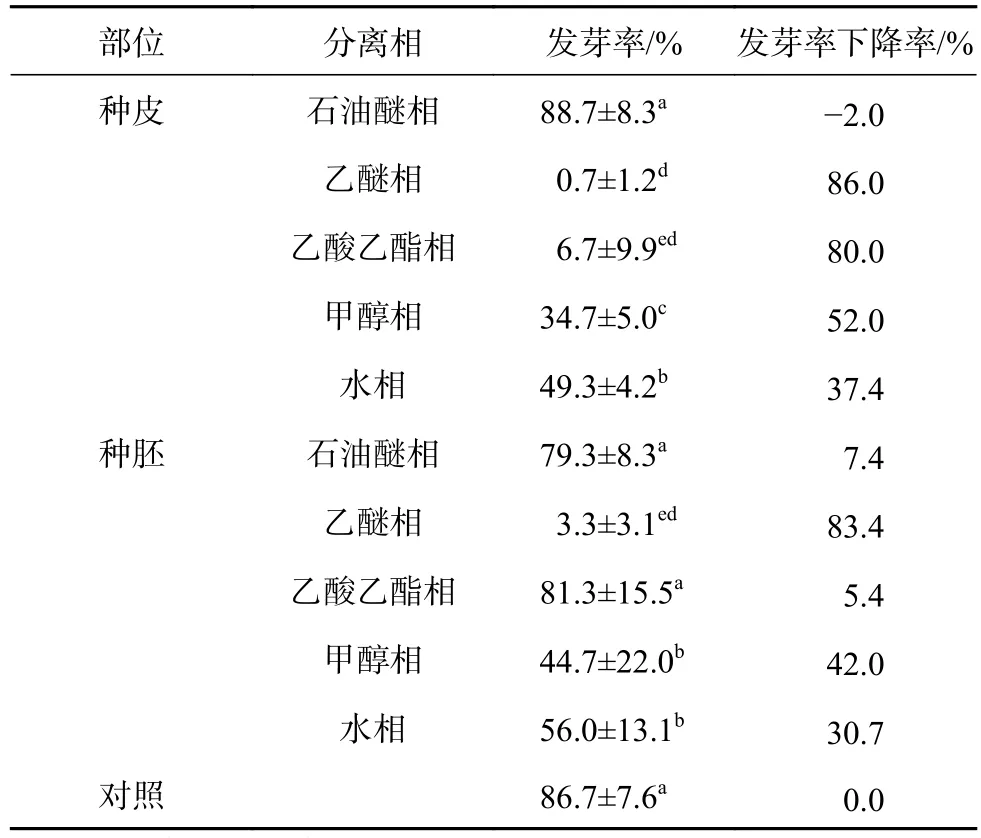
表1 鳳丹種子不同部位各相提取物對白菜籽發芽的影響Table 1 Effects of extracts of P . ostii ‘Fengdan' seeds to the germination of cabbage seeds
2.2 鳳丹種皮各分離相GC-MS鑒定結果
以抑制作用最強的乙醚相為例,經GC-MS分析得到的總離子流色譜圖見圖1,通過質譜系統檢索并與標準譜圖進行核對,篩選總離子流色譜圖中峰面積較大、相似度大于80%的有機化合物,將質譜圖結果與計算機檢索譜庫圖比對后,得到了吻合性較好的14種物質,見表2。由表2可知,種皮乙醚相中14種有機物主要為酚類物質3種(雙酚F、苯酚、2,6-二叔丁基對甲酚),相對含量為12.3%;醇類物質2種(beta-谷甾醇、羽扇豆醇),相對含量為10.1%;酸類物質2種,棕櫚酸、反油酸,相對含量為6.4%;酰胺類物質1種,芥酸酰胺,含量為1.4%,其他物質2種,未知物質4種。
圖1 鳳丹種皮中乙醚相提取物的離子流程Fig. 1 The ion flow of extract with aether solution in seed coat of P. ostii ‘Fengdan'
按照相同的方法,篩選出種皮其他相中的有機物,見表3。由表3可知,種皮乙酸乙酯相中共鑒定出17種有機化合物,主要為酚類物質,其總相對含量為20.3%,主要包括雙酚F、苯酚、鄰苯二酚、間苯三酚,另外還有有機酸類、酰胺類物質,如棕櫚酸、苯甲酸、反油酸、芥酸酰胺、油酸酰胺等,還含有少量的6,7-二羥基香豆素[7](1.1%)。種皮甲醇相也鑒定出17種有機物,主要為酚類和有機酸物質,其相對含量較高的為苯甲酸(13.2%)、鄰苯二酚(10.5%)、苯酚(9.4%)、芥酸酰胺(2.6%)等。種皮石油醚相共鑒定出24種物質,主要是烷類物質其總相對含量為20.4%,還含有棕櫚酸(3.7%)、芥酸酰胺(3.3%)等物質。
2.3 種胚分離相的GC-MS鑒定結果
本次篩選出種胚各有機相中的化合物見表4。種胚乙醚相、甲醇相、乙酸乙酯相和石油醚相分別鑒定出了34、26、24、26種有機化合物,各相鑒定出的物質種類數比種皮部位稍多些。種胚乙醚相中鑒定出的有機化合物相對含量較高的主要是有機酸類(19.9%)及其酯類化合物(18.5%),有機酸類主要為苯甲酸、油酸、棕櫚酸、硬脂酸等,酯類物質可能是由于提取及測試過程中某些酸類物質發生了酯化作用造成的,因此有機酸的實際相對含量可能還要高些;另外,該有機相內雖然含有2,6-二叔丁基對甲酚、芥酸酰胺、苯酚等物質,但是相對含量較低。甲醇相主要為酚類和有機酸類物質,酚類物質主要為苯酚,其相對含量為8.3%,苯甲酸及其酯化物的總含量為8.8%。乙酸乙酯相中鑒定出的主要為酯類物質,存在于該相的酚類物質為苯酚、2,6-二叔丁基對甲酚,有機酸類為反油酸、硬脂酸、苯甲酸和棕櫚酸,酰胺類為芥酸酰胺和油酸酰胺,但酚類、有機酸類和酰胺類物質含量都只在1%左右。石油醚相中鑒定出的有機化合物主要為酯類(20.1%)、烷類(19.1%)和醇類(13.8%)物質,該有機相中也存在甲基苯、酰胺類以及有機酸類物質,但是其含量很低。

表2 鳳丹種子種皮乙醚相中的化合物及相對含量Table 2 Compounds and their relative contents in aether extract of P . ostii ‘Fengdan' seed coat

表3 鳳丹種皮各分離相的化合物及相對含量Table 3 Compounds and their relative contents in seed coat extracts of P . ostii ‘Fengdan'

表4 鳳丹種胚各分離相的化合物分類及相對含量Table 4 Compounds and their relative contents in decoated seeds extracts of P . ostii ‘Fengdan'
3 結論與討論
鳳丹種子萌發需要較長時間,這與其種子具有深休眠特性有關[8]。本研究從抑制物角度,探討引起鳳丹種子生理休眠的原因。以往的研究表明,內源抑制物的存在是導致許多種子休眠的主要原因之一,如黃精(Polygonatum sibiricum)[9]、烏桕(Sapium sebiferum)[10]、南方紅豆杉(Taxus chinensisvar.mairei)[11]等種子中都含有內源抑制物。對鳳丹種子各有機相的生物測定結果同樣表明,種子中內源抑制物質的存在可能是導致其休眠的原因之一。
要確定內源抑制物的存在,通常采用系統溶劑法對種子的甲醇相提取液進行分離,并結合各有機相對白菜籽等非休眠植物種子的發芽實驗進行生物測定。尚旭嵐等[12]采用系統溶劑法對青錢柳(Cyclocarya paliurus)種子的浸提液進行分離,通過生物測定發現抑制物質主要存在于乙醚相中;史鋒厚等[13]對南京椴(Tiliam iqueliana)種子的研究發現,果皮中的抑制物質主要存在于乙醚相和甲醇相中,而種子中的抑制物質主要在甲醇分離相中;張艷杰[14]對南方紅豆杉種子的研究中發現,抑制物質主要存在于種子的乙醚相和甲醇相中。本研究中,對各有機相生物測定結果比較后發現,種皮和種胚的乙醚相的抑制作用最強,這和上述研究結果相似,因此該種子乙醚相鑒定出的物質可能與其生理休眠有密切關系。
以往的研究表明,酚類、有機酸類是植物種子中常見的抑制物質,它們通過阻礙種子吸水進程、抑制呼吸作用、抑制生物酶的活性以及阻礙胚根生長等方式,對種子萌發產生抑制作用[15-17]。近年來,發芽抑制物質的分離與鑒定已成為種子休眠機理研究的重要內容。
Eiuhellin[18]認為,酚類物質能夠抑制種子萌發所需的關鍵酶的代謝,最終對種子的萌發有影響。酚類化合物對植物生長也具有較強的抑制作用,其對水葫蘆的抑制作用主要表現在使水葫蘆葉片失水干枯,根毛變黑,易脫落[19]。錢存夢[20]對烏桕種子休眠的研究中證實了2,6-二叔丁基對甲酚是重要的內源抑制物;韓東等[21]發現鄰苯二酚對西洋參(Panax quinquefolius)種子萌發具有重要的抑制作用,劉序等[22]在香榧種子抑制其發芽和生長的內源物篩選研究中,發現鄰苯二酚對白菜種子的發芽率和幼苗生長的抑制作用最強。本次GC-MS檢測結果表明,對白菜籽發芽有顯著影響的種皮乙醚相、甲醇相以及種胚的甲醇相中,酚類物質的相對含量較高,分別為12.3%、22.6%、9.4%,并且這幾個相中同樣含有苯酚、2,6-二叔丁基對甲酚和鄰苯二酚等酚類內源抑制物質,因此酚類物質可能與該種子休眠有關。另外,這幾個相中雙酚F的相對含量也比較高,該物質在以往的研究中被證明對生物會有毒害作用,如潘芳[23]、任文娟等[24]發現雙酚F對斑馬魚幼苗具有一定的毒害作用,但其對種子休眠是否有影響還需要進一步研究。
有機酸類物質也是已知的內源抑制物質,王艷華[25]認為苯甲酸、棕櫚酸、硬脂酸等是大山櫻(Prunus sargentii)種子休眠的重要內源抑制物質。李龍秀等[26]研究發現苯甲酸對蘿卜種子萌發及幼苗生長有明顯的抑制作用,表現為高濃度時抑制,低濃度時抑制作用減弱的特點。本次種皮的乙醚相、甲醇相以及種胚的乙醚相、甲醇相中,有機酸相對含量也比較高,分別為6.4%、16.9%、19.9%、4.1%,而且有機酸在實驗過程中容易酯化,因此其實際相對含量可能還要高些。本次抑制作用較強的種胚乙醚相中,苯甲酸的相對含量高達17.4%,同時還含有一定量的棕櫚酸和硬脂酸,這些酸類物質可能是該相對白菜籽發芽抑制較強的重要原因。
徐敬東[27]在對紫藤(Wisteria sinensis)的研究中發現,提取物芥酸酰胺對真菌活性具有一定的抑制作用,蔡夢可[28]對花梨木(Peterocarpesspp.)類木材的耐腐性機理的研究中也發現了芥酸酰胺,認為其對真菌的活性有一定抑制作用。酰胺類物質中的油酸酰胺也已被證實能夠誘導動物產生生理性睡眠[2]。本次各有機相中,除了酚類和有機酸類物質,還含有一定量的酰胺類物質,因此酰胺類物質對牡丹種子萌發的影響需要深入研究。
GC-MS結果分析時還發現,鳳丹種皮乙酸乙酯相中相對含量最多的為酚類,占20.7%,而種胚則以酯類物質為多,酚類物質的相對含量只有5.7%;而且種皮中有機酸類和酰胺類物質的相對含量也高于種胚。表1中種皮乙酸乙酯相對白菜籽發芽的影響顯著強于種胚(二者的發芽率分別是6.7%,81.3%),造成這一顯著差異的原因可能也是由于種皮中擁有較多的酚類、有機酸類、酰胺類物質。
本實驗在種皮、種胚石油醚相中鑒定出的有機化合物主要為烷類、酯類、醇類等物質,結合表1的生物測定結果,這些物質可能不是造成鳳丹種子休眠的重要內源抑制物質。此外,種皮石油醚相中有機酸的相對含量達7.0%,但其生物測定的抑制性不高,該相相對含量較高的是棕櫚酸,因此酚類、有機酸類和酰胺類物質中,哪些物質與鳳丹種子休眠的有關還需進一步研究。表1生物活性測定結果中,水相對白菜籽發芽也具有一定的抑制作用,但由于儀器的限制,水相中具體的抑制物質并未能確定出來。
綜上所述,內源抑制物的存在是鳳丹種子生理休眠的原因之一,初步分析發現,其內源抑制物的主要類型可能是酚類、有機酸類及酰胺類物質。本研究中乙醚相的抑制作用最強,其可能含有影響種子休眠的主要抑制物質。種皮抑制物的主要成分是酚類物質,可能是雙酚F、苯酚、鄰苯二酚、2,6-二叔丁基對甲酚等;種胚的主要抑制物質是有機酸類物質,如苯甲酸、油酸、棕櫚酸、硬脂酸等;一些酰胺類物質可能也具有抑制作用。本實驗被鑒定出的物質中,有些物質的抑制作用已被證實,但其他物質是否也能抑制種子萌發以及和鳳丹種子休眠的關系有待進一步證實。其中酚類、有機酸類和酰胺類物質,哪一類物質對鳳丹種子生理休眠起主要作用,以及相互之間對種子生理休眠的作用關系也有待進一步研究。
