絹蒿荒漠主要植物幼苗發生的數量動態及其影響因素
2019-10-22 02:28:06李娜娜劉福元魯為華陳乙實孫海榮靳省飛張凡凡
草業科學 2019年9期
關鍵詞:植物
李娜娜,劉福元,魯為華,陳乙實,孫海榮,靳省飛,張凡凡
(1.省部共建綿羊遺傳改良與健康養殖國家重點實驗室/ 新疆農墾科學院,新疆 石河子 832000;2.石河子大學動物科技學院,新疆 石河子 832003)
絹蒿(Seriphidium)荒漠廣泛分布在新疆的南北疆,其中北疆地區分布面積占總面積(114.25 hm2)的80%以上,是當地牧民主要的春秋放牧場[1]。絹蒿荒漠主要以各類絹蒿為建群種;以禾本科和莎草科為優勢種;以藜科、十字花科、百合科和牻牛兒苗科等多年生、一年生及短命和類短命植物為伴生種,物種多樣性較為豐富。但由于絹蒿荒漠生境的復雜多變,使荒漠植物幼苗在空間分布以及數量上形成不同的特征[2-3]。而幼苗階段作為植物生命周期中最脆弱、最敏感的階段之一,生境的多變必定會對植物幼苗的分布和數量造成一定的影響[4]。對絹蒿荒漠來講,其環境的多變主要表現在土壤和氣候上。土壤制約荒漠植物的生命過程,土壤因子分布變化是影響荒漠植物分布和群落數量特征的重要驅動力,已有研究表明,在干旱和半干旱生態系統中,土壤的性質、地形、水分對植物種類的空間分布尤為明顯,地形的不同在一定程度上控制了太陽輻射和降水的空間再分配,往往形成局部小氣候環境,從而影響土壤水分和養分的分布,使荒漠植物呈斑塊分布[5-7]。并且,由于放牧家畜糞便的沉積和放牧壓力的相互作用,加速了草地上糞便和枯落物的累積,導致土壤有機質的儲存,從而能夠直接增加土壤中營養元素的含量,加快了生態系統的營養循環率,使得植物幼苗的數量及分布發生了改變[8]。從氣候角度來講,溫度和降水量是荒漠生態系統中極為敏感的主要因素,降水量直接影響了土壤水分的儲存量,過高的溫度增大了荒漠區的蒸發量而間接導致了土壤水分的降低,荒漠區較少降水量與炎熱氣候的共同作用,使得土壤干旱愈來愈嚴重,從而導致荒漠植物多樣性及生產力發生巨大的改變。由此可見,土壤因子(pH、有機質、糞便、凋落物、水分)、地形、氣候及自然因素的耦合作用均是影響荒漠植物分布及幼苗數量的重要因素[9]。然而,絹蒿荒漠擁有特殊的地理位置和復雜多變的環境,其特點是干旱少雨、溫度、濕度和降水的時間變異大,土壤鹽堿化嚴重以及人為的干擾,這些特點共同決定了荒漠草地植物的分布格局和數量特征[10-11],絹蒿荒漠植物也在長期的進化歷程中形成了與極端環境相適應的生長對策[12]。迄今為止,關于絹蒿荒漠土壤環境、坡向及氣候環境對植物幼苗的發生類型和數量動態變化的研究比較缺乏,明確絹蒿荒漠植物幼苗分布、數量特征和環境因子之間的關系,對遏制荒漠化、促進植被恢復,維護荒漠穩定具有現實意義。
為此,以新疆絹蒿荒漠為研究區,通過測定其土壤環境因子、水熱條件以及年內幼苗發生類型及數量動態等參量,利用DCCA排序進行環境要素和幼苗發生數量特征的分析,并對不同時間幼苗數量動態變化規律與水熱條件進行回歸和相關分析,探究土壤因素、坡向、時間及氣候因素對幼苗發生類型和數量動態變化的影響,以期回答以下問題:1)絹蒿荒漠的幼苗發生類型可分為幾種?影響荒漠植物幼苗分發生類型的土壤環境因素有哪些?2)坡向和時間及其交互作用對幼苗的分布和數量產生怎樣的影響?3)主要植物幼苗數量動態變化特征與當地水熱氣候條件的關系如何?
1 材料與方法
1.1 研究區概況
試驗區位于天山北坡中段低山帶山前丘陵及平原荒漠區 (84°58′-86°24′ E,43°26′-45°20′ N),是北疆的絹蒿荒漠集中分布區,屬于溫帶荒漠干旱氣候。地勢相對平坦,無特別大的起伏或斷層,海拔900-1 300 m。年平均溫度5~7 ℃,月平均最高溫度集中在7月,為26.6 ℃,最低溫度集中在1月,為-18.5 ℃。年降水集中在6月-8月,年平均降水量在150~210 mm,蒸發量是降水量的4~5倍或以上。土壤為淡栗鈣土。植被組成以伊犁絹蒿(Seriphidium transiliense)為建群種,另廣泛伴生有藜科、十字花科、百合科、禾本科和牻牛兒苗科等多年生、一年生及短命和類短命植物,植被結構組成復雜,且存在季節動態變化。在生產利用上為春秋放牧場,春季放牧時間為4月-6月,秋季放牧時間為9月-11月。
1.2 數據獲取
于2016年5月7號,在典型的絹蒿荒漠草地分布區,使用GPS定位,選取具有代表性的樣地,分別于陽坡、陰坡、半陽坡、半陰坡、平地取樣,每個方位平行取10個1 m×1 m的樣方框,對各樣方內的植物種類及其幼苗進行詳細計數,共計獲得50個樣方數據,形成樣方×物種的二維數據表。
在進行樣方調查時,用土壤緊實度儀對每個樣方進行土壤緊實度測定,并采集每個樣方內10 cm深土壤,用于土壤有機質、pH測定。收集地表枯落物、家畜糞便裝袋,帶回實驗室風干稱重。通過上述工作獲得每一樣方包括土壤緊實度、土壤有機質和pH、枯落物量、家畜糞便量5個環境指標,形成樣方×環境要素的二維數據表。
為闡明主要植物幼苗數量隨時間推移而產生的數量動態變化與當地氣候條件之間的關系。于2016年4月-11月,分別按陽坡、陰坡、半陽坡、半陰坡、平地5個方位布置固定監測樣方,每個方位布置3個重復固定樣方,每月中旬進行主要植物幼苗的數量統計,得到不同時間點的幼苗數量數據。同時從中國氣象數據共享網上下載2016年1月-12月本地的降水、平均氣溫、相對濕度等氣象數據。
1.3 數據分析及統計
使用Excel 2007軟件進行所有數據的初步整理,不同區域幼苗數量比較的方差分析,不同時間幼苗數量動態變化規律與氣候因素關系的回歸和相關分析利用DPS7.5完成。利用Canoco4.5軟件的DCCA模塊進行環境要素和幼苗發生數量特征的排序分析,以闡明幼苗發生類型和環境要素之間的關系。
2 結果與分析
2.1 荒漠植物幼苗發生類型形成的環境解釋
環境因子基本統計量以及與幼苗數量的相關性如表1和表2所列。50個樣方的環境因子基本統計參數結果表明,各環境因子的變異較大,其中糞便量的變異系數達到1.216 5,凋落物重量變異系數為0.797 4,土壤有機質變異為0.440 0(表1)。
從環境因子和各類主要植物幼苗總數量的相關分析可知,凋落物重量與幼苗數量呈顯著負相關關系(P<0.05)。糞便量、土壤有機質及土壤緊實度與幼苗總數存在極顯著正相關關系(P<0.01),土壤pH值和幼苗數量之間相關性不顯著(P>0.05)(表2)。
由于大部分環境因子和幼苗總數量存在相關性,進一步對50個調查幼苗的樣方采取DCCA進行分類排序,結果如圖1所示。DCCA排序軸第一軸基本反映了枯落物、土壤pH、土壤有機質和糞便量的變化,排序軸自左向右,土壤pH逐漸減小,而枯落物、土壤有機質和糞便量逐漸增大;第二軸基本反映了土壤緊實度的數量變化趨勢,排序軸自上而下,土壤緊實度逐漸增加。
從50個樣方在二維排序圖上的分布來看,各樣方在排序圖上有規律地分布。可根據樣方在DCCA排序圖上的位置及其與各環境軸之間的位置關系將其劃分為可劃分為3種類型,坐標軸右下方的11、12、13、14、16、19、20、41~50等17個樣方幼苗總數量和糞便重量密切相關,這些樣方的幼苗數量主要采集于陰坡和平地,主要是由羊茅(Festuca ovina)、蚤綴 (Arenaria serpyllifolia)、角果毛茛(Ceratocephalus orthoceras)、絹蒿、串珠老鸛草(Geranium transversale)、頂冰花(Gagea lutea)幼苗組成,可以認為這些物種為糞便主導影響的幼苗發生類型;而15、17、31~ 38等11個樣方內幼苗主要和枯落物重量及土壤有機質密切相關,這些樣方的幼苗數量主要采集于半陰坡和陰坡,主要由絹蒿、羊茅和庭薺(Alyssum desertorum)幼苗組成;其余 1 ~ 10、21 ~ 30、39、40等 22 個樣方位置在二維排序圖的左上部,這些樣方的幼苗數量主要采集于陽坡和半陽坡,主要由絹蒿、庭薺和針茅(Stipa capillata)組成,角果藜和叉毛蓬數量較少,這些物種幼苗發生主要受土壤緊實度和pH的影響。
從DCCA排序圖中可以看出,3種幼苗發生類型中的兩種是和糞便、枯落物以及土壤有機質有關,這兩類樣方數量占總樣方數量的56%,說明糞便、枯落物可為幼苗的發生提供良好的微環境條件,保證了種子落粒后萌發過程中所需要的水分條件,增加了土壤環境條件的異質性。同時糞便內所攜帶的經過家畜消化道作用后而排放出的大量種子,也增加了種子的輸入數量,從而使幼苗總數量增加。而豐富的土壤有機質則有利于幼苗萌發和后續存活生長。以上環境要素不僅為幼苗輸入創造條件,而且也增加了植物群落特征的異質性,這對草地小尺度上豐富格局的形成提供了環境和物質基礎。
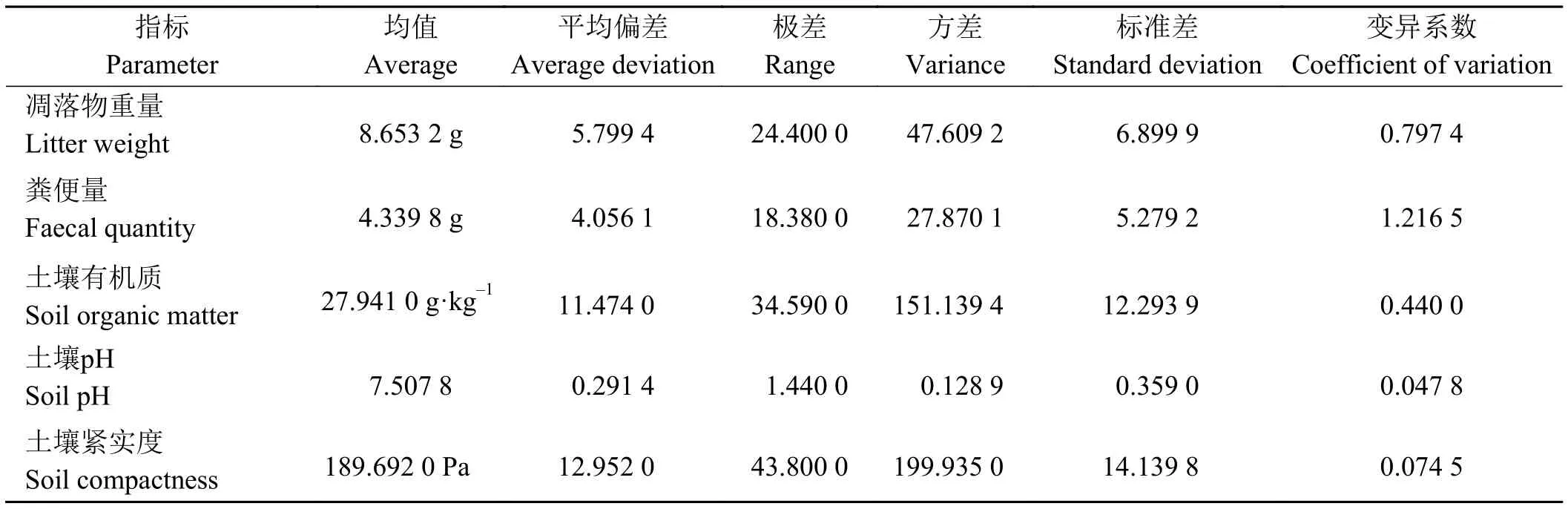
表1 不同區域50個樣方各環境因子基本統計參數Table 1 Fundamental statistical parameters of environmental factors

表2 主要植物幼苗總數量與各環境因子之間的相關性分析Table 2 Correlation analysis of seedling numbers and associated environmental factors
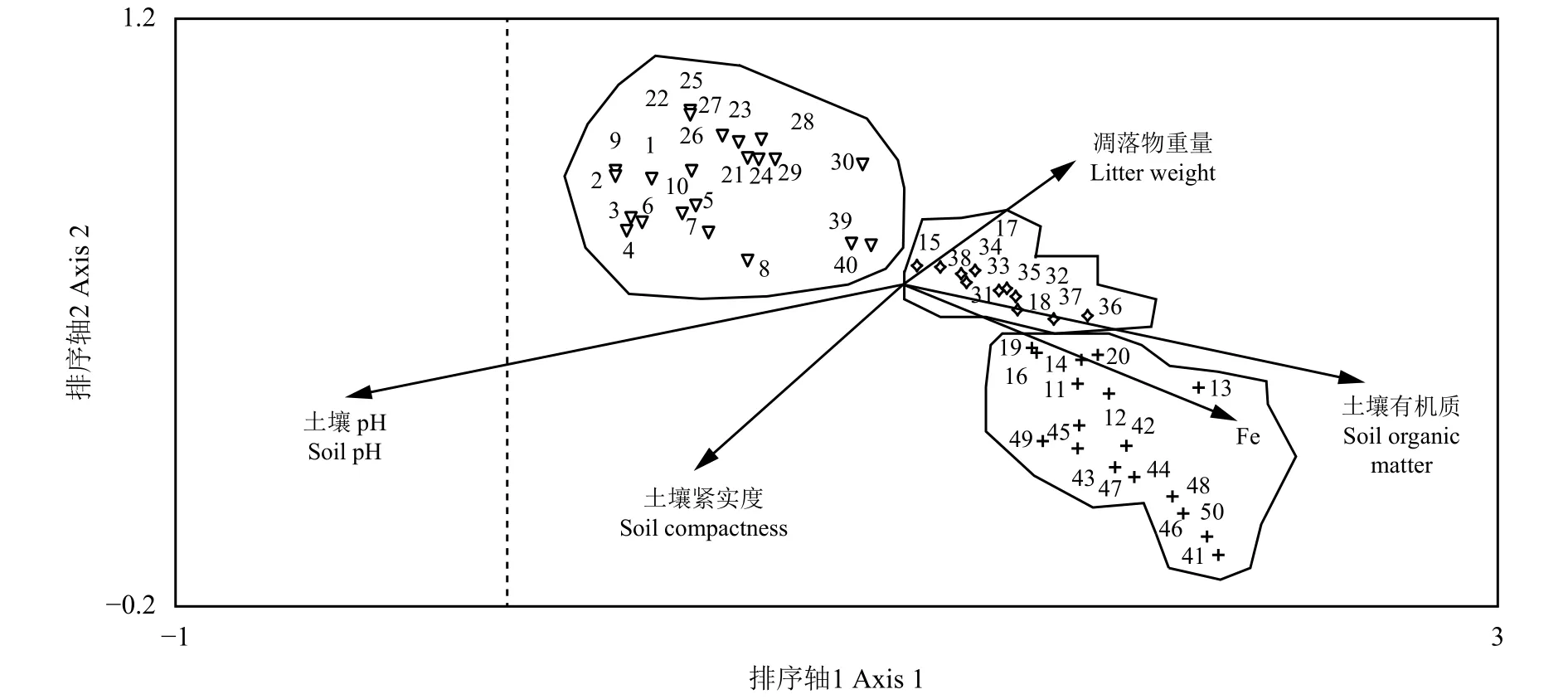
圖1 50個樣方的DCCA二維排序圖Figure 1 Two-dimensional DCCA ordination diagram of 50 quadrats
2.2 主要植物幼苗數量動態變化特征及成因分析
2.2.1 主要植物幼苗數量差異及動態變化特征
對絹蒿荒漠區4種主要植物在不同時間和坡向的幼苗數量進行二因素方差分析(表3),結果表明,不同坡向上絹蒿幼苗、羊茅幼苗、庭薺幼苗、角果毛茛幼苗的數量差異均達到極顯著水平(P<0.01),不同時間段,除了角果毛茛外,其他3種植物幼苗的數量與時間的差異均極顯著(P<0.01)。此外,時間和坡向的交互作用對4種植物幼苗數量均有極顯著的影響(P<0.01)。由此說明,時間和坡向均是影響這4種植物幼苗數量的重要因素。
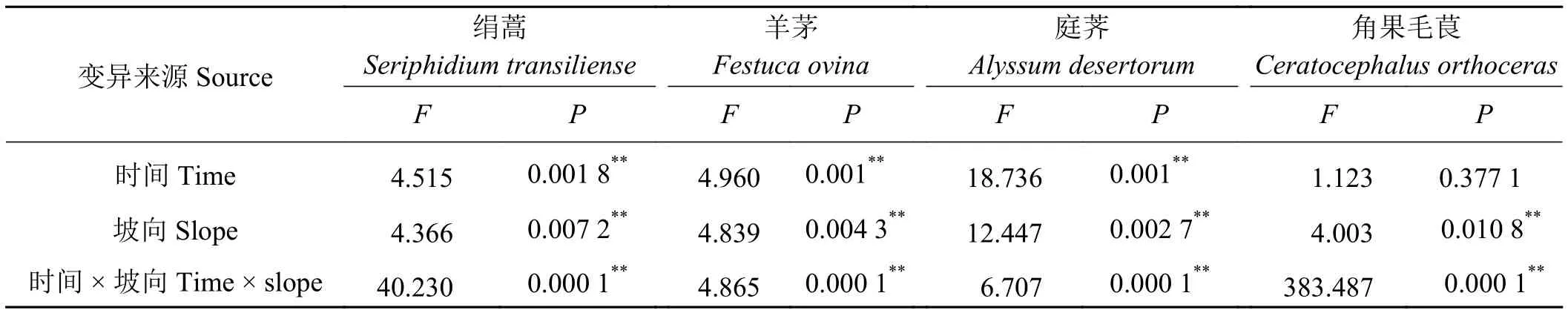
表3 4種主要植物幼苗數量特征二因素方差分析Table 3 Two-way ANOVA test showing seedling characteristics of the four main plant species
對不同坡向的4種植物幼苗4月-11月的發生數量進行統計并作數量動態變化擬合曲線,結果表明,在生長季內,絹蒿幼苗數量在不同坡向上存在顯著差別,但其數量動態變化模式是一致的,其變化過程為:5月15日至6月之間,除了半陽坡外,其余坡向絹蒿幼苗數量呈現出峰值,隨后開始下降,9月份開始,不同坡向的幼苗數量逐漸增加,約在10月15日再次呈現出高峰,隨后降低,呈現出明顯的雙峰變化,且陽坡和半陽坡有利于絹蒿幼苗的生長(圖2)。陰坡有利于羊茅幼苗的生長,幼苗在陽坡、半陰坡和平地的數量變化模式一致,均符合線性模型,而陰坡和半陽坡數量變化分別符合高斯模型和指數模型。角果毛茛幼苗主要生長在平地區域,發生數量在生長季呈現“雙峰”型。庭薺幼苗發生數量較少,主要生長在陽坡和半陽坡,平地次之。其中陽坡數量呈現春季和秋季數量多,而夏季數量少的趨勢,半陽坡幼苗數量呈現“單峰”狀態。
2.2.2 主要植物幼苗數量動態形成原因分析

圖2 4個主要物種的幼苗數量變化Figure 2 Changes in quantity of the four plant seedlings at different times of year
對試驗區3月-11月份月平均溫度和月降水量進行分析發現(圖3),溫度隨時間的推移表現為先增加后降低的趨勢,其中6、7月份溫度達到最高,但降水量隨時間的推移表現為先增加后降低再增加的趨勢,在4、5月份降水量偏多,加之有春季的積雪融水,為各類植物幼苗發生提供了充足的水分條件,從而出現“爆發性出苗”的現象。從6月份開始降水逐漸減少,至9月份降水量最低,氣候極為干旱,從而導致幼苗出現 “驟然性死亡”現象,此后,溫度降低,降水量又逐漸增加,溫度和水分的改善勢必有利于部分幼苗的第2次發育和新生幼苗發生。
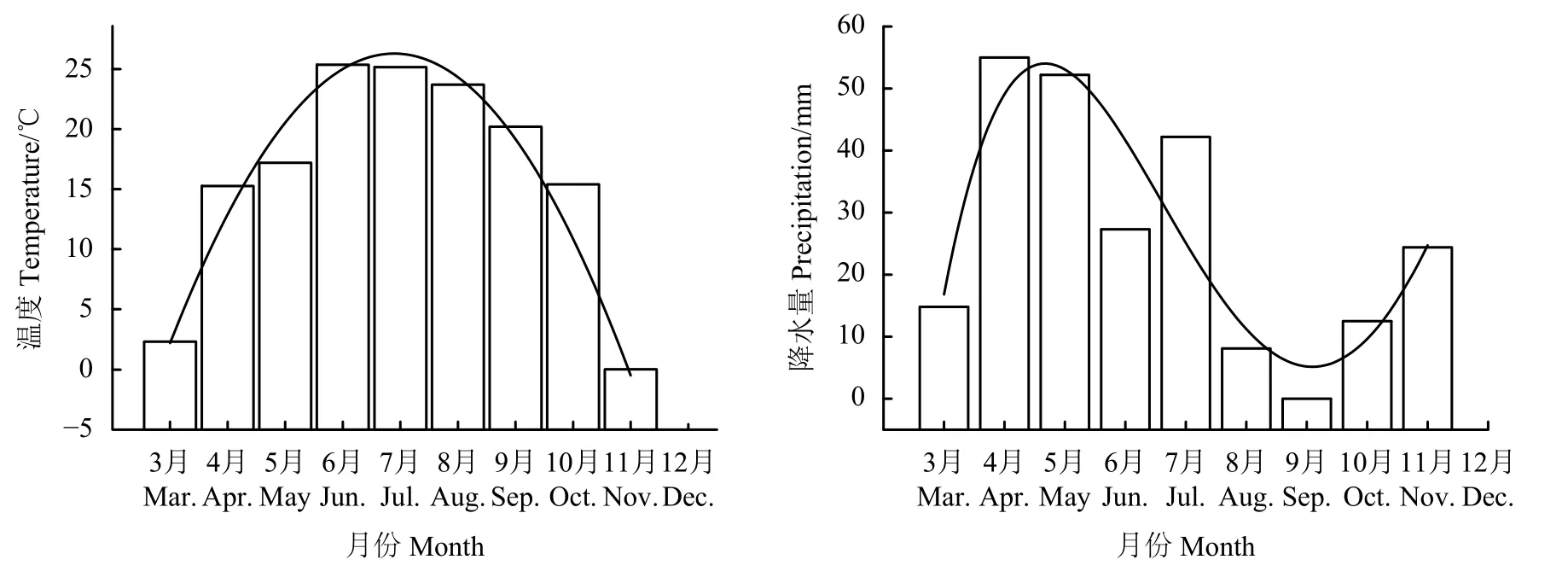
圖3 2016年3月-11月的月平均溫度和月平均降水量Figure 3 The average temperature and precipitation levels during March to November of 2016
為了探討月平均溫度和降水量對植物幼苗數量的影響,進行了相關性分析(表4),月平均降水量與4種幼苗的數量之間存在極顯著正相關(P<0.01),月平均溫度與絹蒿幼苗數量存在極顯著正相關(P<0.01),與羊茅、角果毛茛和庭薺幼苗數量的相關性不顯著(P>0.05)。由此可見,年內降水是影響各類植物幼苗數量動態的主要因素,溫度作為附加條件對幼苗發生也有一定影響。進一步對幼苗數量降水、溫度作回歸分析,4種植物幼苗數量和兩個氣候因子之間的關系均符合線性回歸模型。

表4 降水與溫度對主要植物幼苗數量動態變化特征的影響相關性分析Table 4 Correlation analysis of the effects of precipitation and temperature on the quantity of plant seedlings
3 討論
3.1 土壤環境條件影響絹蒿荒漠植物幼苗發生類型
幼苗是植物生活史中最為關鍵的階段,是連接植物代際間的紐帶,幼苗數量不僅能反映親代數量和質量特征,而且更為重要的是可預測未來種群動態和演變趨勢[13]。本研究利用DCCA排序分析方法,將絹蒿荒漠50個幼苗樣方劃分為3種發生類型,其中兩種類型與糞便、枯落物以及土壤有機質密切相關。盡管幼苗發生數量和土壤pH相關性未達到顯著水平(表2),但當考慮發生類型時發現,有1種幼苗發生類型與土壤pH和緊實度相關。眾多研究表明,土壤的理化性質對植物的幼苗發生乃至群落結構類型起著決定性的作用。對植物群落或者幼苗發生類型產生顯著影響的生境因子主要有土壤電導率、黏粒含量、有機質含量、凋落物、家畜糞便、土壤含水量和pH等[14]。并且,由于區域內環境條件的異質性會導致幼苗或成年植物群落在空間內產生分異,從而產生幼苗數量和結構方面的不同類型。且不同發生類型的幼苗群落在結構組成和數量特征上存在本質差別。趙鵬對民勤綠洲荒漠過渡帶的研究發現,不同類型植被群落空間分布的主導環境因子不盡相同[15],這與本研究結果一致。
絹蒿荒漠土壤有機質含量較低,是幼苗輸入并成功定居的限制因子之一,而荒漠區有機質的主要來源是凋落物和糞便的分解,植物幼苗對土壤有機質含量的變化反應非常敏感,從而會產生有機質主導的幼苗發生類型。有機質作為土壤主要的養分,對促進植物幼苗的生長發育起著關鍵的作用[9,16]。糞便不僅可以增加土壤有機質、氮、鉀、磷等元素和土壤水分,而且糞便內含有經過家畜消化道作用后的大量種子,增加了種子的輸入數量,同時也為植物幼苗的生長與存活提供良好的環境[17]。牛鈺杰等[18]通過放牧作用下高寒草甸群落物種分布與土壤因子關系的研究認為,放牧作用下土壤緊實度是影響群落物種分布格局的最關鍵因子,對荒漠區植物尤其是草本植物群落的發生和生態功能的發揮十分重要[18-19]。土壤緊實度的增加會造成立地條件的變化,進而影響到種子的萌發和幼苗的定居。本研究中pH主導下的幼苗發生類型樣方內主要以耐鹽堿的物種為主,隨著pH的增加,耐鹽堿物種的幼苗增加而其他物種數量減少。種子萌發環境中的pH會對其萌發行為產生顯著的影響,這在很多有關種子萌發試驗的文獻中都有報道[20-22],在絹蒿荒漠中,陽坡及半陽坡的土壤pH要高于其他區域,在該區域所發生的植被類型則適應于高pH土壤環境,而幼苗的發生類型分類也明顯的體現出這一點,樣方中的絕大多數都分布在陽坡和半陽坡。有學者提出在堿性土壤中植物物種多樣性分布與土壤pH不存在顯著相關性[23],這與本研究結果相似,可能是由于植物長期生長在鹽堿化的荒漠地區中,已經形成了良好的適應對策。
3.2 地形、水熱條件是絹蒿荒漠幼苗數量動態變化的主導因素
植物在長期的進化發展過程中,逐漸形成了獨特的繁殖策略,以增強其對異質生境的適應性[24]。在絹蒿荒漠這樣特殊的生境中,地形和氣候因子的變化會導致植物幼苗發生動態變化。本研究發現坡向與不同植物幼苗數量均達到極顯著水平,絹蒿幼苗在半陽坡生長最多,陽坡次之;羊茅幼苗在陰坡數量最多;角果毛茛整體上主要生長在平地上;庭薺數量雖在絹蒿荒漠區較少,但其主要生長在陽坡、半陽坡區域,充分說明不同的地形影響植物的組成與生長發育。然而地形是固定不變的因素,影響植物生長發育的可能是由于不同坡向的環境(土壤水分、土壤養分、土壤質地、風速、溫度等)不同,不同坡向決定了表層土壤接受太陽輻射的量,造成土壤水分蒸發量的差異性,從而導致土壤水熱再分配,進而營造出不同的微氣候環境,故在不同坡向表現出不同的植物分布的變化規律[25-28];也可能是因為4種植物對生態位的要求不同,植物面對不同生境所選擇不同的生長策略。已有大量研究表明,地形主要是通過影響光照、水分進而影響植物生存條件的選擇[29-30]。
在荒漠區,不同時間段內降水和溫度等氣候條件波動劇烈,從而造成幼苗數量的顯著差異[31-32]。強烈的同胞競爭和波動劇烈的環境條件是絹蒿荒漠幼苗輸入效率低下的主要原因[33],但這種篩選對于荒漠區植物群落穩定性維持十分重要[34]。因為荒漠區可利用資源的有限性對植物種群數量的擴展有嚴格限制。本研究中4種植物幼苗的發生規律總體上呈現春季“爆發性出苗”,夏季“驟然性死亡”的特點,同時在秋季又會以二次發育或者秋萌的形式形成一個幼年個體輸入的峰值,這種幼年個體的輸入方式是長期適應荒漠氣候的結果。已有研究表明,無論是在亞熱帶地區內還是在暖溫帶地區內,溫度和降水都是影響植物幼苗生長的顯著因子[35]。De Almeida等[36]通過光照和溫度對刺芒幼苗萌發和生長的研究發現,適宜的溫度會加速幼苗的萌發與生長,過高的溫度會對幼苗的生長造成不利的影響。Shan等[37]研究了西北地區降水量及頻率對幼苗出苗及生長的影響,研究發現降水量及頻率的增加顯著地增加了幼苗的出苗率和生長率。光照和溫度的變化在一定程度上也決定了土壤的含水量[38]。絹蒿荒漠區早春氣溫回升快,積雪融化,降水量較多,土壤水分相對充足;秋季降水量逐漸升高。春秋兩季水分和溫度條件的適宜使植物幼苗集中發生,同時,在秋季還會因水熱條件良好而產生秋萌現象。然而,夏季降水量較少,溫度較高,導致幼苗地上部“驟然性死亡”,地上部生長停滯。由此可見,氣溫和水分對植被生產力的影響有顯著的時空異質性,絹蒿荒漠典型的氣候波動變化使其幼苗生長規律更為明顯[39-40]。
4 結論
1)絹蒿荒漠幼苗的3種發生類型中有2種是與家畜糞便量、土壤有機質、凋落物密切相關,1種與土壤緊實度和pH有關,上述這些因素均是影響幼苗發生類型的主要驅動力。2)不同坡向的主要植物幼苗發生數量存在極顯著差異,同一坡向不同時間的幼苗發生數量也存在顯著差異,且地形和時間對于幼苗數量的發生有顯著的交互作用。各類植物幼苗由于植物本身的生物學特性和環境條件時空異質性,在發生類型和數量上也會存明顯的差異。3)年內生長季的降水量和氣溫變化決定了植物幼苗數量的動態變化,夏季的高溫和低水分條件對各類植物幼苗有強烈的環境篩選作用,各類植物幼苗為了保證存活定居與荒漠區的溫度、降水節律保持高度的同步性,幼苗數量動態與溫度和降水有顯著的相關性。因此,絹蒿荒漠幼苗在長期波動的氣候條件下形成了鮮明的環境適應。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
