血散薯內生真菌的分離鑒定及其抗菌活性研究
2019-10-21 08:00:50顏楨靈李國萍駱海玉陸保屹梁春霞鄧業成鄧志勇
河南農業科學 2019年10期
顏楨靈,李國萍,駱海玉,陸保屹,梁春霞,鄧業成,鄧志勇
(珍稀瀕危動植物生態與環境保護省部共建教育部重點實驗室/廣西珍稀瀕危動物生態學重點實驗室/廣西師范大學 生命科學學院,廣西 桂林 541006)
植物內生真菌指的是一類全部或部分生活史存在于植物健康組織或器官中,卻不引起宿主植物產生明顯感染病癥的微生物[1-2],其普遍存在于各種植物體中,種類繁多,分布廣泛。不同宿主植物不同器官的內生真菌種類差異較大[3-11]。植物內生真菌與宿主植物協同進化、互惠互利,其不僅對植物的生長、發育、進化及生態適應性產生重要影響,還可以提高宿主植物適應惡劣環境的能力[12]。
隨著傳統化學藥劑的長期大量使用,病蟲害的耐藥性日益加重,同時也引發水體污染、土壤污染、野生動物或人畜中毒等一系列生態問題,因此,尋找環境友好型的新農藥迫在眉睫[13]。而植物內生真菌長期協同進化,可產生種類豐富、活性顯著的代謝產物,尤其是來源于藥用植物的內生真菌,已成為挖掘新型天然活性物質的寶貴資源庫[14-15]。目前,已有許多研究報道藥用植物內生真菌可產生萜類及其皂苷、甾體、生物堿、肽類、芳香類等具有抗菌、抗蟲、抗氧化、抗腫瘤等多種生物活性的物質[2,16]。內生真菌中的活性物質,具有可持續利用、對自然資源破壞小等優點,可緩解自然資源短缺的問題,特別是能為保護珍稀瀕危藥用植物資源提供新思路,是近年來的一個研究熱點[14]。內生真菌產生的次生代謝產物在植物病蟲害生物防治、醫藥工業上的用途和范圍逐漸擴大[17]。
血散薯(StephaniadielsianaY.C.Wu),別名金不換、一滴血,為防己科千金藤屬植物,是我國南部及西南部地區重要的藥用植物,味苦、性寒,有清熱解毒、活血散瘀、止痛、抗炎、涼血、治療蛇毒咬傷等多種藥理功能[18-20]。此外,其甲醇粗提物及活性物質千金藤堿(Stephanine)和克斑寧(Crebanine)等具有顯著、廣譜的抗菌活性[21]。但血散薯僅生長于廣西、廣東、貴州南部及湖南南部等地的山谷、溪邊、林中及峭壁上,分布范圍窄,資源有限,加之人們缺乏對血散薯的保護意識,近年來過度采挖,已造成該資源的嚴重稀缺。基于共生理論,認為血散薯內生真菌可能具有多種生物活性。目前,對血散薯內生真菌的相關研究尚未見報道。因此,本研究從血散薯莖、葉中分離純化內生真菌,對其進行鑒定,并探究其次生代謝產物的抗菌活性,以期為血散薯植物資源的保護及其內生真菌的合理開發利用提供依據。
1 材料和方法
1.1 血散薯
2016年8月于廣西來賓市金秀瑤族自治縣(E109°59′34″,N24°16′10″),采集血散薯新鮮健康莖、葉組織,標本保存于廣西師范大學珍稀瀕危動植物生態與環境保護省部共建教育部重點實驗室化學生態實驗室。
1.2 供試病原菌
供試植物病原真菌:選取9種常見農業植物病原真菌作為供試菌株,包括貢柑鏈格孢菌(Alternariacitri)、金橘砂皮病菌(Diaporthecitri)、辣椒炭疽病菌(Colletotrichumcapsici)、茶輪斑病菌(Pestalotiopsistheae)、甘藍黑斑病菌(Alternariaoleracea)、甘蔗鳳梨病菌(Ceratocystisparadoxa)、水稻胡麻葉斑病菌(Cochliobolusmiyabeanus)、煙草黑脛病菌(Phytophthoraparasiticavar.Nicotianae)、玉米大斑病菌(Exserohilumturcicum)。以上9種植物病原真菌除金橘砂皮病菌和貢柑鏈格孢菌由廣西特色作物研究所提供以外,其余7種均由廣西大學農學院植物病理室提供。
供試動物病原細菌:選取10種供試動物病原細菌作為供試菌株,包括大腸桿菌(Escherichiacoli)、炭疽桿菌(Bacillusanthraci)、溶壁微球菌(Micrococcuslysodeikticus)、藤黃微球菌(Micrococcusluteus)、銅綠假單胞菌(Pseudomonasaeruginosa)、金黃色葡萄球菌(Staphyloccocusaureus)、普通變形桿菌(Proteusvulgaris)、巨大芽孢桿菌(Bacillusmegaterium)、蠟狀芽孢桿菌(Bacilluscereus)、枯草芽孢桿菌(Bacillussubtilis)。以上10種動物病原菌均由桂林醫學院提供,保存于4 ℃備用。
1.3 培養基
內生真菌分離培養基:馬鈴薯200 g/L、葡萄糖20 g/L、氯霉素200 mg/L、瓊脂15 g/L、蒸餾水1 L。
植物病原真菌以及內生真菌純化培養基為馬鈴薯葡萄糖瓊脂培養基(PDA):馬鈴薯200 g/L、葡萄糖20 g/L、瓊脂20 g/L、蒸餾水1 L。
動物病原細菌培養基為牛肉膏蛋白胨培養基:牛肉膏3 g/L、蛋白胨10 g/L、氯化鈉5 g/L、瓊脂20 g/L、蒸餾水1 L。
種子液體培養基為馬鈴薯葡萄糖液體培養基(PDB):馬鈴薯200 g/L、葡萄糖20 g/L、氯霉素200 mg/L、蒸餾水1 L。
內生真菌發酵培養基為大米固體發酵培養基:大米∶水約為1∶1(m/V)。
1.4 內生真菌分離與純化
參考LUO等[22]、王利娟等[23]的方法,對內生真菌進行分離純化。首先選取新鮮的血散薯莖、葉組織,用清水洗凈表面,吸干表面殘留水分,再按以下步驟進行內生真菌的分離:(1)在超凈工作臺內進行無菌操作,將血散薯莖、葉組織切片(段)。(2)將切片(段)后的血散薯組織進行表面消毒。先用75%乙醇漂洗30 s,轉入無菌水中,沖洗3~4次后,放置在無菌濾紙上將多余水分吸干。再用2%次氯酸鈉漂洗90 s,轉入無菌水中,沖洗3~4次后,放置在無菌濾紙上將多余水分吸干。最后用0.1%氯化汞漂洗3 s,無菌水沖洗3~4次后,放置在無菌濾紙上將多余水分吸干。(3)在每個用PDA培養基制作的平板上放入3~4塊已進行過表面消毒的組織片(段)。設置2組對照:一組取最后一次漂洗的無菌水100 μL加至新鮮無菌PDA平板上,涂布,逐日觀察平板上是否有菌長出;另一組采用按壓法,將消毒后的組織塊放在新鮮無菌PDA平板上,稍加按壓,放置20 min后移去組織塊。每組處理設置3個平行,以驗證血散薯表面消毒是否徹底,若無菌長出,則可以斷定分離出的真菌來自血散薯莖、葉組織內部。(4)分離組織在恒溫培養箱內,(28±1)℃暗培養1~25 d,逐日觀察。待分離組織邊緣長出菌絲后及時挑取,將其轉入另一PDA平板培養,重復操作,直至長出的菌落形態完全一致。
1.5 內生真菌鑒定
主要采用形態學和分子生物學的方法進行鑒定。
形態學鑒定:參考文獻[24]以及廣西師范大學珍稀瀕危動植物生態與環境保護省部共建教育部重點實驗室化學生態實驗室已鑒定至屬種的多種內生真菌菌株等的鑒定方法,采用不同培養基(PDA、CMA、NGA)對分離得到的內生真菌進行培養鑒定。鑒定特征包括菌落大小、形態、生長速率、菌落及培養基基質顏色、菌落邊緣特征,以及菌落在顯微鏡下的特征(如菌絲有無分隔、分支,分生孢子形態、大小,產孢結構特征等)。
分子生物學鑒定:利用真核生物在rDNA的ITS區段的特性,即具有保守性和特異性序列的特性,通過內生真菌的rDNA-ITS序列對其進行鑒定。以真菌通用引物ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)擴增菌株ITS堿基序列,委托北京六合華大基因科技有限公司武漢分公司進行檢測純化及測序。將所獲得的測序結果與NBCI數據庫中的序列進行BLAST比對,選擇相似度較高的菌株,進行同源性比對,并用MEGA Ⅹ軟件,對ITS堿基序列進行聚類分析,利用Bootstrap進行自展次數為1 000的置信度檢測,根據系統發育樹中的組群關系對菌株進行分類。
1.6 內生真菌發酵培養及粗提物制備
在超凈工作臺上,用直徑為4 mm的打孔器在已經活化好的菌落邊緣處打孔,取3塊菌餅接種至含有馬鈴薯葡萄糖液體培養基(150 mL)的錐形瓶(250 mL)中,置于恒溫搖床上,(27±1)℃、130 r/min振蕩培養。培養5 d后,即可得到發酵種子液。取5 mL種子液接種于事先準備好的大米培養基,放置培養箱中,28 ℃培養2個月。
將發酵產物進行干燥、粉碎處理后,用適量的乙酸乙酯分別浸泡提取3次,合并3次的提取濾液,用旋轉蒸發儀濃縮蒸干,即得到次生代謝產物粗提物。
1.7 抗植物病原真菌活性測定
采用菌絲生長速率法[21]測定內生真菌發酵產物粗提物抗植物病原真菌活性。具體操作:樣品用丙酮配成2.5 mg/mL藥液,將藥液(對照用丙酮代替)與溫度為50~55 ℃的PDA培養基按1∶9混合均勻后,倒入直徑為9 cm的培養皿內,冷凝制成帶毒培養基后,每皿分別接種3塊已活化好的直徑為4 mm的供試病原菌菌餅(甘蔗鳳梨病菌每皿接1塊),帶菌絲面朝下貼至培養基表面。每處理3次重復,于28 ℃恒溫培養箱中培養72 h,采用十字交叉法測量菌落生長直徑,取平均值,計算抑菌率:抑菌率=(對照菌落直徑-處理菌落直徑)/(對照菌落直徑-0.4 cm)×100%。測定對植物病原菌菌絲體的有效中濃度(EC50)時,根據初篩時的抑菌率,配制系列不同濃度的藥液,測定其對供試病原真菌的菌絲毒力,用最小二乘法求出毒力回歸方程、EC50等。
1.8 抗動物病原細菌活性測定
先采用濾紙片法進行抗細菌活性初篩,再以帶毒平板法測定活性樣品的最低抑制濃度(MIC)。
濾紙片法:參考林洋等[25]的濾紙片法進行抗細菌活性初篩,具體操作:將熱熔的牛肉膏蛋白胨培養基倒入直徑為9 cm的玻璃培養皿中,冷凝備用。用西林瓶制備供試病原細菌懸液,從培養24 h的細菌培養基中用接種環刮取適量細菌與無菌水進行充分混勻,再用移液槍吸取0.1 mL菌液加至培養基表面,用涂布棒涂布均勻,制成表面含菌培養基。樣品用丙酮溶解并配成10.0 mg/mL藥液,將直徑為6 mm完好的濾紙片浸入藥液(對照以丙酮代替)5 min,取出晾干。將濾紙片貼入平板內適當位置,小心用鑷子壓平,使整個圓形濾紙片與培養基表面貼合,放入37 ℃細菌恒溫培養箱中培養24 h,用十字交叉法測抑菌圈直徑。根據抑菌圈大小,將抑菌效果分為如下幾類:抑菌圈直徑為0 mm者為不敏感;小于10 mm者為輕度敏感;10~15 mm者為中度敏感;大于15 mm者為高度敏感[26]。
MIC值測定:參考DENG等[21]的方法測定樣品MIC值。用丙酮將樣品溶解并配成系列濃度藥液,將藥液(對照用丙酮代替)與溫度為50~55 ℃的熱溶牛肉膏蛋白胨培養基按1∶9混合均勻后,倒入直徑為6 cm的培養皿(5 mL/皿),待凝固后,吸取50 μL菌懸液至培養基表面,用涂布棒涂布均勻。每處理重復3次,于(37±1)℃恒溫倒置培養24 h,觀察細菌的生長情況。
1.9 數據處理
2 結果與分析
2.1 血散薯內生真菌的分離與鑒定
從血散薯莖、葉組織共分離得到38株內生真菌,其中31株基于ITS序列的BLAST比對結果(表1),結合形態學特征,將其歸為14個屬:鏈格孢屬(Alternaria)、炭團菌屬 (Hypoxylon)、枝孢屬(Cladosporium)、莖點霉屬(Phoma)、鐮刀屬(Fusarium)、Stagonosporopsis、Ascomycete、毛殼菌屬(Chaetomium)、擬盤多毛孢屬(Pestalotiopsis)、彎孢霉屬(Curvularia)、短梗霉屬(Aureobasidium)、Annulohypoxylon、小光殼屬(Leptosphaerulina)、刺盤孢屬(Colletotrichum)。鑒定結果表明,31株內生真菌中鏈格孢屬和莖點霉屬為優勢屬,在莖、葉組織中均有分布。其中,鏈格孢屬13株,莖點霉屬4株,表明其可能是血散薯莖、葉中穩定存在的內生真菌。其次為彎孢霉屬和鐮刀屬,各2株,其余10個屬的內生真菌均僅分離得到1株。
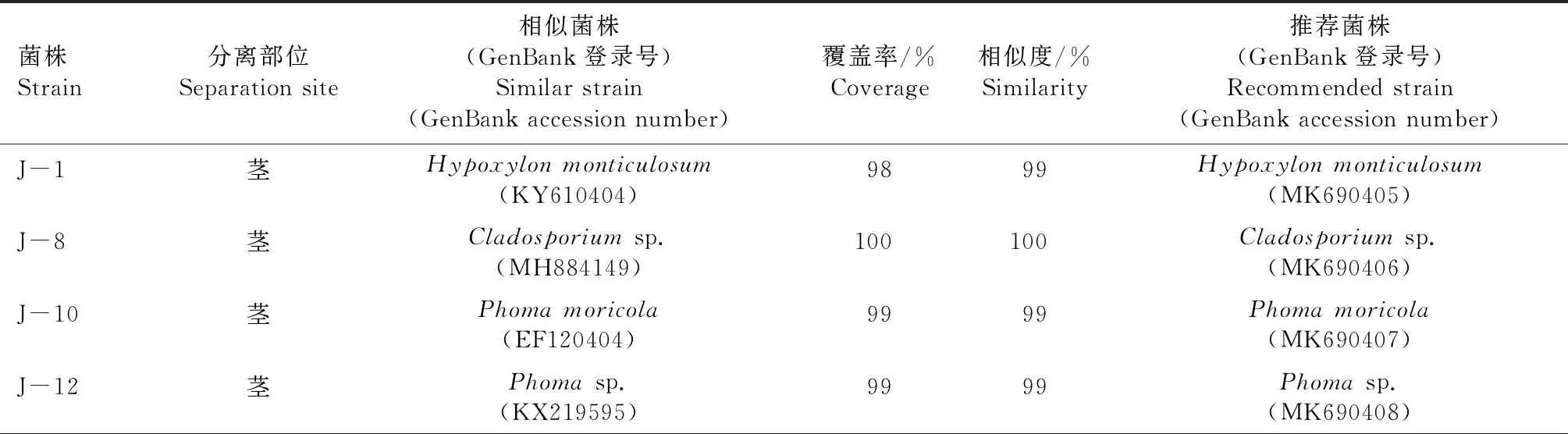
表1 31株血散薯內生真菌ITS序列BLAST比對結果Tab.1 ITS sequences BLAST alignment results of 31 endophytic fungi of Stephania dielsiana
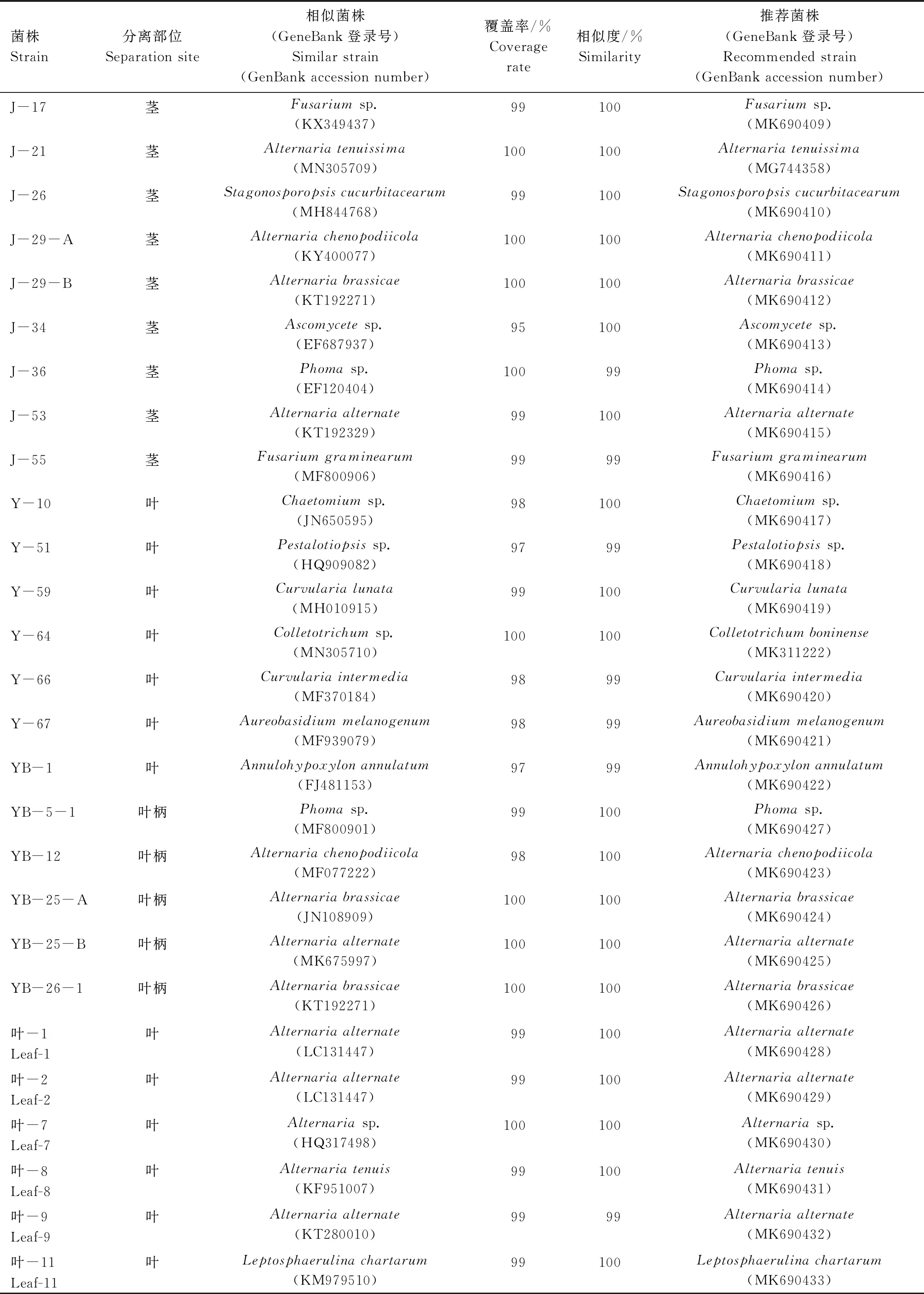
續表1 31株血散薯內生真菌ITS序列BLAST比對結果Tab.1(Continued) ITS sequences BLAST alignment results of 31 endophytic fungi of Stephania dielsiana Y.C.Wu
2.2 血散薯內生真菌對9種植物病原真菌的抗菌活性
基于對峙拮抗預試驗結果,從上述38株內生真菌中選取表1中的13株及未測序的YB-20,共14株真菌進行發酵培養,并進一步測定發酵產物抗植物病原真菌活性,結果如表2所示。在藥液質量濃度為2.5 mg/mL時,14株內生真菌發酵產物的粗提物對9種植物病原真菌具有不同程度的抑制作用。除內生真菌J-26、Y-64外,其余內生真菌的粗提物均至少對1種供試植物病原真菌的抑菌率達到50%以上。其中,Y-66對9種植物病原真菌的抑菌率均在55%以上,抑菌率介于56.81%~100.00%,表現出較為廣譜的抗菌活性。內生真菌葉-8和J-12對除了茶輪斑病菌以外的8種供試植物病原真菌的抑菌率均在50%以上。其中,葉-8對金橘砂皮病菌抑菌率為88.56%,J-12對煙草黑脛病菌抑菌率則達到100.00%。YB-12對除了辣椒炭疽病菌和茶輪斑病菌外的7株供試植物病原真菌的抑菌率均在60%以上,對甘蔗鳳梨病菌、水稻胡麻葉病菌及煙草黑脛病菌的抑制作用最佳,抑菌率均為100.00%。內生真菌J-34、J-36和J-55均對其中5株供試植物病原真菌的抑菌率在60%以上。上述結果表明,血散薯內生真菌具有較好的抗植物病原真菌活性,其中7株內生真菌(葉-8、Y-66、YB-12、J-12、J-34、J-36、J-55)的抗菌活性較為明顯和廣譜。
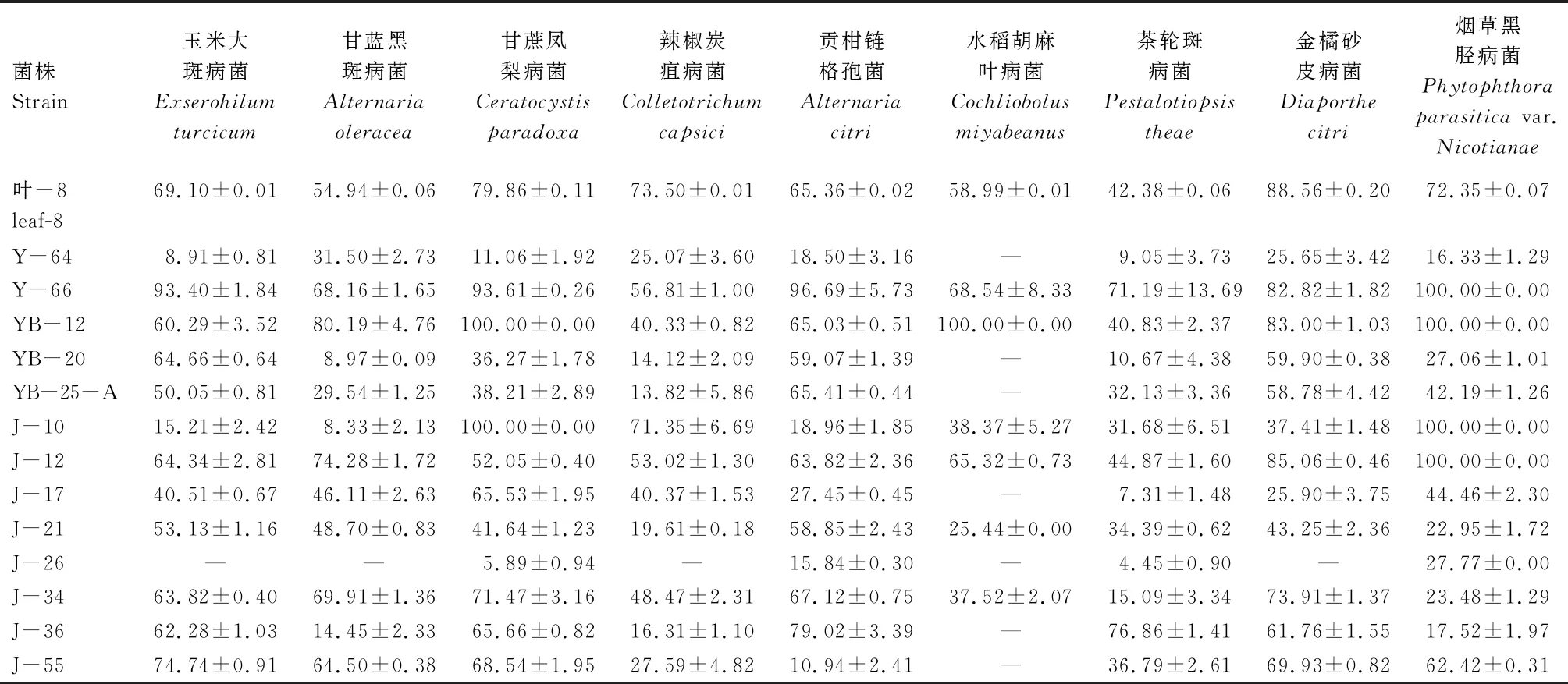
表2 14株血散薯內生真菌發酵產物粗提物對9種植物病原真菌的抑制作用Tab.2 Antifungal activity of the extracts of 14 endophytic fungi of Stephania dielsiana Y.C.Wu against 9 plant pathogenic fungi %
2.3 血散薯內生真菌對9種植物病原真菌菌絲的毒力
為確定以上7株活性菌株(葉-8、Y-66、YB-12、J-12、J-34、J-36、J-55)的抗真菌活性,結合以上抗菌活性測定結果,選取該7株活性內生真菌發酵產物粗提物對植物病原真菌抑菌率達到50%以上的相應病原菌作為供試菌株,進一步測定其對供試菌株菌絲的毒力,結果見表3。葉-8粗提物對玉米大斑病菌、甘蔗鳳梨病菌、貢柑鏈格孢菌、金橘沙皮病菌、煙草黑脛病菌具有較好的毒力,EC50值介于0.048 0~0.363 5 mg/mL,均小于0.5 mg/mL,其中,對玉米大斑病菌、甘蔗鳳梨病菌、金橘砂皮病菌具有較好的抗菌活性,其EC50值分別為0.063 1、0.059 5、0.048 0 mg/mL,均小于0.1 mg/mL。Y-66粗提物對其中6株供試植物病原真菌的EC50值介于0.133 3~0.679 0 mg/mL,其中,對甘蔗鳳梨病菌具有最好的抗菌活性,EC50值為0.133 3 mg/mL。J-12粗提物對其中5株供試植物病原真菌的EC50值介于0.006 2~0.700 3 mg/mL,其中,對金橘砂皮病菌具有最好的抗菌活性,EC50值為0.006 2 mg/mL。YB-12粗提物對其中6種供試植物病原真菌的EC50值小于1.0 mg/mL,介于0.115 7~0.615 4 mg/mL,其中,對甘蔗鳳梨病菌具有最好的抗菌活性,EC50值為0.115 7 mg/mL。J-34粗提物對供試的5株植物病原真菌的EC50值介于0.004 2~0.522 6 mg/mL,其中,對甘藍黑斑病菌、甘蔗鳳梨病菌、貢柑鏈格孢菌均具有極好的抗菌活性,EC50值分別為0.018 1、0.004 2、0.084 6 mg/mL。J-36粗提物對玉米大斑病菌、貢柑鏈格孢菌的抗菌活性較好,EC50值分別為0.964 9、0.709 0 mg/mL。J-55粗提物對其中3種植物病原菌的EC50值小于1.0 mg/mL,其中,對甘藍黑斑病菌具有最好的抗菌活性,EC50值為0.280 8 mg/mL。

表3 7株血散薯內生真菌發酵產物粗提物對植物病原真菌菌絲的毒力Tab.3 Toxicity of the extracts of 7 endophytic fungi of Stephania dielsiana Y.C.Wu to mycelium of plant pathogenic fungi
2.4 血散薯內生真菌對2種動物病原細菌的抗菌活性
采用濾紙片法對2.2中14株內生真菌發酵產物粗提物的抗動物病原細菌活性進行初篩,發現該14株內生真菌具有一定抗菌活性。為進一步確定其抗菌作用,采用帶毒培養基法測定其抗菌活性。當藥液質量濃度為10.0 mg/mL時,有10株內生真菌對至少1種動物病原細菌具有抗菌活性(表4)。進一步測定其MIC值,發現10株內生真菌對金黃色葡萄球菌的MIC值介于0.500~10.000 mg/mL,其中,YB-25對金黃色葡萄球菌的抗菌活性最好,MIC值為0.500 mg/mL。僅有4株內生真菌(葉-8、YB-20、J-12、J-17)對大腸桿菌具有抗菌活性,MIC值介于0.156~4.000 mg/mL,其中,J-12對大腸桿菌的抗菌活性最佳,MIC值為0.156 mg/mL。葉-8、YB-20、J-12、J-17對2種供試動物病原細菌具有較好抑制作用。總體來看,內生真菌J-12發酵產物粗提物對10種動物病原細菌中的2種(金黃色葡萄球菌、大腸桿菌)抑制作用相對較好(MIC值分別為1.250、0.156 mg/mL),明顯高于已報道的血散薯甲醇提取物對2種供試動物病原菌的抗菌活性(MIC值分別為3.75、7.50 g/L),并高于Stephanine和Crebanine對大腸桿菌的抗菌活性(MIC值均大于5 g/L)[21]。

表4 10株血散薯內生真菌發酵產物粗提物抗動物病原細菌的活性Tab.4 Antibacterial activities of the extracts of 10 endophytic fungi of Stephania dielsiana Y.C.Wu against animal pathogenic bacteria
注:---表示未長菌;+++表示長菌;/表示無抗菌活性。
Notes: --- indicates no bacteria; +++ indicates expressing bacteria; /indicates no antibacterial activities.
3 結論與討論
本研究首次報道了血散薯內生真菌的分離鑒定及其抗菌活性。從血散薯莖葉中共分離得到38株內生真菌,其中31株已通過形態學結合分子生物學的方法鑒定,將其歸為14個屬。其中,優勢屬為鏈格孢屬和莖點霉屬,在莖、葉組織中均有分布,表明其可能是血散薯莖葉中穩定存在的內生真菌。鏈格孢屬和莖點霉屬在已報道的許多植物內生真菌中也均為優勢屬[22,27-28],可見鏈格孢屬和莖點霉屬是植物中普遍存在的內生真菌。
經內生真菌發酵產物粗提物抗菌活性研究發現,分離得到的大部分血散薯內生真菌具有良好而廣譜的抗菌活性。其中,抗植物病原真菌活性的研究結果表明,7株內生真菌(葉-8、Y-66、YB-12、J-12、J-34、J-36和J-55)均具有較為明顯和廣譜的抗植物病原真菌活性。而在抗動物病原細菌方面的研究結果表明,10株供試內生真菌的發酵產物粗提物對金黃色葡萄球菌均具有抗菌活性,但僅有4株內生真菌對大腸桿菌表現出抗菌活性。以上結果與前人[21]報道的血散薯植物抗菌活性相似,即血散薯甲醇粗提物及其活性成分Stephanine和Crebanine對植物病原真菌具有明顯而廣譜的抗菌活性,對金黃色葡萄球菌均具有不同程度的抑制作用,而對大腸桿菌的抗菌活性較差。
本研究結果表明,血散薯莖、葉組織中存在豐富的內生真菌資源,且其次生代謝產物含有廣譜抗菌活性物質。后期可對其活性物質進行分離和鑒定,以尋找活性化合物等,為生物源殺菌劑的創制提供更多優秀的先導化合物,同時也將為活性天然產物提供重要的資源,并對其在農業及醫藥領域的應用具有重要意義。
