準(zhǔn)噶爾東南緣梭梭幼苗生境生態(tài)因子及其生長
2019-10-21 11:25:00羅艷麗張玲衛(wèi)仝伯凱
新疆農(nóng)業(yè)科學(xué) 2019年8期
粟 玲,王 澤,羅艷麗,麻 浩,3,任 財,張玲衛(wèi),仝伯凱
(1.新疆農(nóng)業(yè)大學(xué)干旱區(qū)荒漠研究所,烏魯木齊 830052;2.新疆農(nóng)業(yè)大學(xué)草業(yè)與環(huán)境科學(xué)學(xué)院/新疆土壤與植物生態(tài)過程實(shí)驗室,烏魯木齊 830052;3.南京農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,南京 210095)
0 引 言
【研究意義】梭梭(Haloxylonammodendron)是藜科(Chenopodiaceae)落葉小喬木或灌木,是西北荒漠半荒漠地區(qū)的珍貴植物資源[1]。梭梭分布面積最大的準(zhǔn)噶爾盆地,自然條件惡劣,生態(tài)環(huán)境脆弱,以梭梭為優(yōu)勢種的植物群落為近200種荒漠植物提供了繁衍和棲息生境,是荒漠地區(qū)生物多樣性保護(hù)的重要場所,亦是當(dāng)?shù)亟?jīng)濟(jì)與社會發(fā)展的綠色屏障[2-5]。研究梭梭幼苗對生境條件變化的適應(yīng)狀況,對當(dāng)?shù)鼗哪鷳B(tài)系統(tǒng)的穩(wěn)定與維持意義重大。【前人研究進(jìn)展】前人對梭梭生理生態(tài)學(xué)特性、群落結(jié)構(gòu)和生物學(xué)等方面進(jìn)行了大量的研究,魯燕等[8]研究表明,低濃度的NaCl處理(≤50 mmol/L)對梭梭生長具有一定的促進(jìn)效應(yīng),梭梭具有一定適應(yīng)鹽漬土壤環(huán)境的能力;YANG等[9]、許皓等[10]研究發(fā)現(xiàn)梭梭為非深根型植物,主要水源是降水形成的淺層土壤水,其用水策略是根據(jù)水分條件有效調(diào)節(jié)根系和冠層生長,從而維持正常的光合作用;王春玲等[11]研究發(fā)現(xiàn)準(zhǔn)噶爾盆地東南緣不同生境梭梭群落天然更新幼苗幼樹的分布格局均為聚集分布;蘇培璽等[12]研究發(fā)現(xiàn)梭梭同化枝具有花環(huán)結(jié)構(gòu),肉細(xì)胞呈柵欄狀,且有巨大的含晶細(xì)胞,具有適應(yīng)高溫、強(qiáng)光荒漠環(huán)境的光合作用結(jié)構(gòu)。光照強(qiáng)度是指光照的強(qiáng)弱,以單位面積上所接受可見光的能量來量度[15]。土壤溫度作為地表主要物理參量之一,是土壤作用于植物的重要指標(biāo)[16]。【本研究切入點(diǎn)】針對荒漠不同梭梭幼苗生境區(qū)的生態(tài)因子與幼苗生長適應(yīng)性對比方面的研究較少。幼苗階段是梭梭個體生長最為脆弱、對環(huán)境變化最為敏感的時期,由于幼苗生存環(huán)境的異質(zhì)性,受到外界生態(tài)因素如光照、溫度和水分等綜合影響,使得個體數(shù)量發(fā)生變化,進(jìn)而影響種群更新。研究幼苗生境區(qū)的生態(tài)因子及幼苗個體生長生理特征,及時掌握特定區(qū)域幼苗的適應(yīng)性有助于預(yù)測梭梭種群的演替變化。【擬解決的關(guān)鍵問題】研究準(zhǔn)噶爾東南緣不同梭梭幼苗生境區(qū)光照強(qiáng)度、土壤溫度和土壤水分等生態(tài)因子的動態(tài)變化規(guī)律,分析生態(tài)因子變化對幼苗個體生長生理的影響,研究幼苗對荒漠逆境條件的響應(yīng)機(jī)制,為梭梭抗逆機(jī)理和種群生態(tài)保育提供科學(xué)依據(jù)。
1 材料與方法
1.1 材 料
研究區(qū)位于新疆準(zhǔn)噶爾盆地東南緣古爾班通古特沙漠吉木薩爾縣林業(yè)管護(hù)站(E88°49′46.97″,N 44°32′59.88″)周邊的荒漠區(qū),地勢南高北低,海拔500~700 m,屬中溫帶大陸干旱氣候,年平均氣溫6.7℃,年平均降水量165~180 mm,年蒸發(fā)量2 100~2 300 mm,年平均相對濕度60%左右,≥10℃的積溫3 500~3 550℃,全年日照時數(shù)3 100~3 200 h,無霜期150~170 d[13]。冬春兩季有降雪,積雪深度達(dá)10~35 cm,積雪時間長達(dá)4個月。植物群落以梭梭為優(yōu)勢種,伴生有白梭梭(H.persicum)、多枝檉柳(Tamarixramosissima)、白皮沙拐棗(Calligonumleucocladum)、角果藜(Ceratocarpusarenarium)、對節(jié)刺(Horaninoviaulicina)等[14]。
試驗前在林業(yè)管護(hù)站周邊分布廣泛的平緩沙地和平緩低洼地生境類型的梭梭群落內(nèi),選取具有代表性的幼苗生境區(qū)A和B,面積均為500 m2。
1.2 方 法
1.2.1 試驗設(shè)計
每個試驗區(qū)設(shè)置10個5 m × 6 m的調(diào)查樣方,在每個樣方內(nèi)隨機(jī)選取1~3年生長勢相對一致、無病蟲害、生長發(fā)育正常的梭梭幼苗10株掛牌標(biāo)記,作為試材。平緩沙地,地形起伏2~5 m,土壤為風(fēng)沙土,質(zhì)地以細(xì)沙為主,土壤pH 8.1,土壤總鹽含量為1.95 g/kg,有機(jī)質(zhì)含量為4.68 g/kg。丘間低地,地形起伏1~2 m,土壤為灰棕色荒漠土,壤質(zhì),土壤表層有腐殖質(zhì)殘留,土壤pH 8.2,土壤總鹽含量為2.03 g/kg,有機(jī)質(zhì)含量為4.95 g/kg,兩種生境類型的梭梭群落生存所需的水源均來自降水或地表徑流。
1.2.2 測定指標(biāo)
分別于2017年4月21日、7月18日、8月30日和11月12日,監(jiān)測每個試驗區(qū)各調(diào)查樣方的光照強(qiáng)度、土壤溫度和土壤水分含量等生態(tài)因子指標(biāo),其中:采用LA-1017數(shù)字照度計測定地表的光照強(qiáng)度,采用CENTER-300便攜式測溫儀測定0 cm、0~3 cm,3~6 cm,6~9 cm層的土壤溫度,采用TDR-100便攜式土壤水分測定儀測定0 cm、0~3 cm,3~6 cm,6~9 cm層的土壤水分含量。于相同時期動態(tài)監(jiān)測試驗區(qū)各調(diào)查樣方內(nèi)標(biāo)記梭梭幼苗個體的生長指標(biāo)(株高、冠幅、地徑和梢長等),采用游標(biāo)卡尺測定幼苗地徑,用卷尺測定幼苗株高、冠幅和梢長,利用SPAD-502葉綠素儀測定標(biāo)記幼苗同化枝葉綠素含量的SPAD值。
1.3 數(shù)據(jù)處理
采用SPSS 17.0和Excel2007軟件進(jìn)行統(tǒng)計分析,采用新復(fù)極差法(SSR)在P<0.05水平上進(jìn)行差異顯著性檢驗并完成制圖。
2 結(jié)果與分析
2.1 不同梭梭幼苗生境區(qū)的生態(tài)因子
2.1.1 不同梭梭幼苗生境區(qū)的地表光照強(qiáng)度變化
隨著生長期的推進(jìn),梭梭幼苗生境區(qū)A與B的地表光照強(qiáng)度均呈先升高后降低的變化趨勢,最大值均出現(xiàn)在7月,二者的地表光照強(qiáng)度值分別為78 500和82 000 lx;最小值均出現(xiàn)在11月,其光照強(qiáng)度值分別為19 800和23 800 lx。各生長期幼苗生境區(qū)A與B的地表光照強(qiáng)度相比,生境區(qū)A的地表光照強(qiáng)度均低于生境區(qū)B,4、7、8和11月各生長期生境區(qū)A的地表光照強(qiáng)度較生境區(qū)B分別低2 400、3 500、6 500和4 000 lx,差異顯著(P<0.05)。整個生長期,不同生長期幼苗生境區(qū)地表光照強(qiáng)度變化特征為:7月>8月>4月>11月。圖1
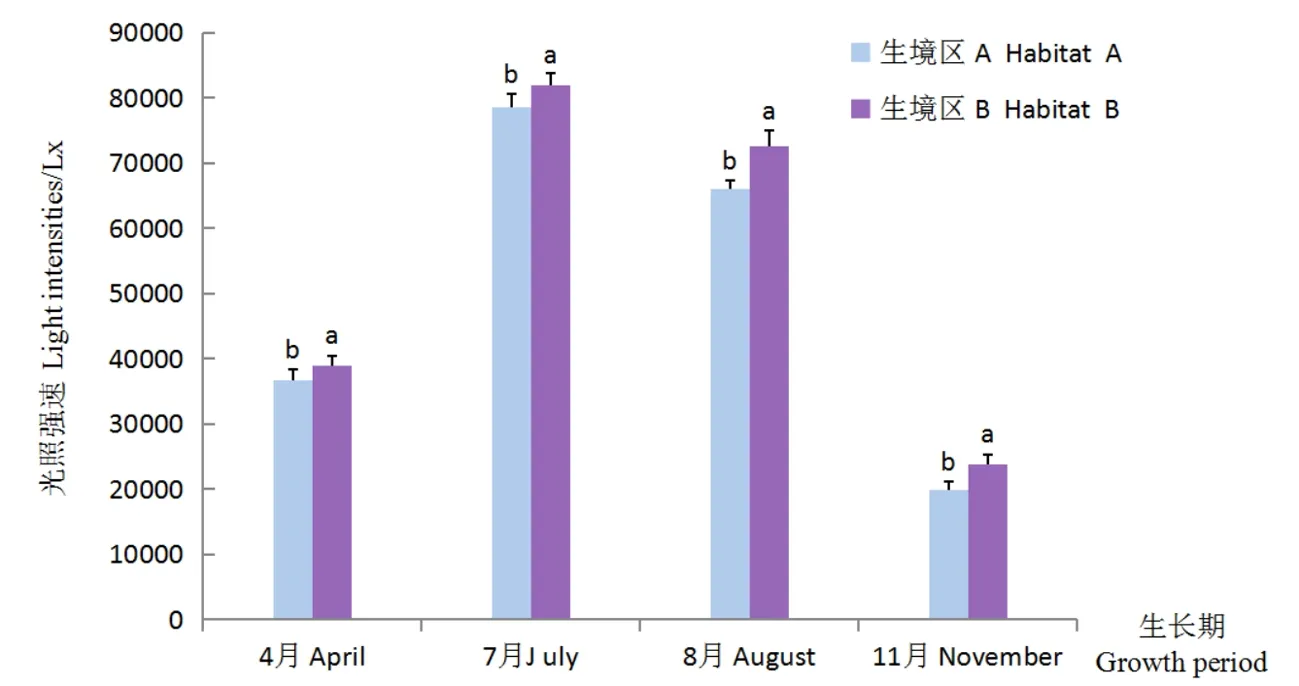
注:“T”形棒表示標(biāo)準(zhǔn)偏差,“T”形棒上方不同字母表示P<0.05水平差異顯著,下同
Note: Vertical T bars indicate standard deviation. Bars topped by different letters indicate significant differences in values(P<0.05), the same as below
圖1 不同梭梭幼苗生境區(qū)的地表光照強(qiáng)度狀況
Fig. 1 Characteristics of surface light intensities in different seedlings habitats
2.1.2 不同梭梭幼苗生境區(qū)的土壤溫度變化
不同生長期梭梭幼苗生境區(qū)A與B表層的土壤溫度變化狀況均表現(xiàn)為:隨著土層深度的增加呈持續(xù)降低趨勢,但不同生長期內(nèi)各土層溫度的高低有所差異。4月,生境區(qū)A與B的表層土壤溫度分別由土壤垂直0 cm的28.92和29.70℃逐漸降低到6~9 cm土層的23.84和24.32℃,生境區(qū)A各土層溫度較生境區(qū)B分別低0.78、0.52、0.52和0.48℃。7月,生境區(qū)A與B的表層土壤溫度分別由地表的41.16和43.06℃逐漸降低到6~9 cm土層的33.98和35.50℃,生境區(qū)A各土層溫度較生境區(qū)B分別低1.90、1.32、1.84和1.52℃。8月,生境區(qū)A與B的表層土壤溫度均達(dá)到最高值,分別由地表的46.28和47.60℃逐漸降低到6~9 cm土層的38.58和40.16℃,生境區(qū)A各土層溫度較生境區(qū)B分別低1.32、1.42、1.90和1.58℃。11月,生境區(qū)A與B的表層土壤溫度分別由地表的13.20和14.10℃逐漸降低到6~9 cm土層的6.50和7.10℃,生境區(qū)A各土層溫度較生境區(qū)B分別低0.90、0.89、0.77和0.60℃。整個生長期,生境區(qū)A的表層土壤溫度均低于生境區(qū)B,且不同生長期幼苗生境區(qū)土壤各土層的溫度變化特征為:8月>7月>4月>11月。圖2
2.1.3 不同梭梭幼苗生境區(qū)的表層土壤水分變化
研究表明,不同生長期梭梭幼苗生境區(qū)A與B表層的土壤水分含量變化狀況均表現(xiàn)為:隨著土層深度的增加呈持續(xù)升高趨勢,但不同生長期內(nèi)各土層的土壤水分含量高低有所差異。4月,生境區(qū)A與B土壤水分含量分別由土壤垂直0 cm的0.62%和0.45%遞增到6~9 cm土層的4.16%和3.34%,其中生境區(qū)A的3~6 cm土層水分含量較生境區(qū)B低0.82%,其它各層的水分含量均高于生境區(qū)B,分別高出0.17%、0.14%和0.88%。在極端干旱的7和8月,兩個生境區(qū)地表的土壤水分含量均為0,7月二者的土壤水分含量分別由0~3 cm層的0.06%和0.04%遞增到6~9 cm層的1.88%和1.34%,生境區(qū)A各土層水分含量較生境區(qū)B分別高出0.02%、0.46%和0.54%;而8月二者的土壤水分含量分別由0~3 cm層的0.05%和0.03%遞增到6~9 cm層的1.72%和1.02%,生境區(qū)A各土層水分含量較生境區(qū)B分別高出0.02%、0.42%和0.60%。11月,生境區(qū)A與B土壤水分含量分別由地表的1.12%和1.10%遞增到6~9 cm土層的6.87%和6.57%,其中生境區(qū)A的各土層水分含量較生境區(qū)B分別高出0.02%、0.13%、0.27%和0.30%。整個生長期,不同幼苗生境區(qū)土壤各土層的水分含量變化特征為:11月>4月>7月>8月。圖3

圖2 不同梭梭幼苗生境區(qū)的土壤表層溫度變化
Fig. 2 Characteristics of soil temperature of surface in different seedlings habitats
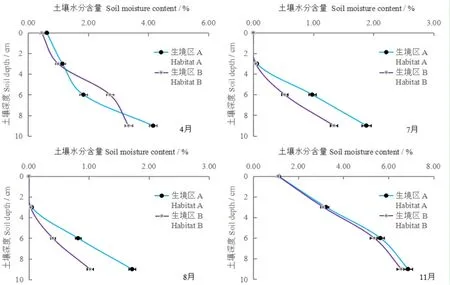
圖3 不同梭梭幼苗生境區(qū)的表層土壤水分變化
Fig. 3 Characteristics of soil moisture content of surface in different seedlings habitats
2.2 不同生境區(qū)梭梭幼苗的生長狀況
研究表明,不同生境區(qū)生長期梭梭幼苗的各類生長指標(biāo)變化不一,隨著生長期的推進(jìn),生境區(qū)A與B梭梭幼苗的株高和地徑呈遞增的變化趨勢,其最大值分別可達(dá)40.81、38.73 cm和0.41、0.36 cm;而冠幅和梢長呈先增加后降低再增加的變化趨勢,其最大值出現(xiàn)在7月,分別為21.65、19.73 cm和18.72、17.12 cm。不同生長期生境區(qū)A與B的幼苗各類生長指標(biāo)差異表現(xiàn)不同,4月,生境區(qū)A的幼苗株高、地徑和梢長值較生境區(qū)B的高,分別高出2.61、0.01和0.16 cm。其中幼苗株高差異顯著。7月,生境區(qū)A的幼苗株高、冠幅、地徑和梢長值均高于生境區(qū)B,分別高出3.64、1.92、0.01和1.60 cm,其中幼苗株高、冠幅和梢長差異顯著。8月,生境區(qū)A的幼苗株高、冠幅、地徑和梢長值也均高于生境區(qū)B,分別高出2.93、1.06、0.03和0.65 cm,其中幼苗株高差異顯著。11月,生境區(qū)A的幼苗株高、冠幅、地徑和梢長值均高于生境區(qū)B,分別高出2.08、2.74、0.05和0.20 cm,其中幼苗株高和冠幅差異顯著。表1
2.3 不同生境區(qū)梭梭幼苗的同化枝葉綠素SPAD值
研究表明,梭梭幼苗生長的關(guān)鍵期7至8月,不同生境區(qū)幼苗的同化枝葉綠素SPAD值均呈遞減的變化趨勢,生境區(qū)A的幼苗同化枝葉綠素SPAD值由7月的18.37下降到8月的9.27,而生境區(qū)B的幼苗同化枝葉綠素SPAD值由7月的16.05下降到8月的10.03。不同生長期生境區(qū)A與B的幼苗同化枝葉綠素SPAD值差異表現(xiàn)不同,7月,生境區(qū)A的幼苗同化枝葉綠素SPAD值較生境區(qū)B高出2.32,差異顯著,而8月,生境區(qū)B的幼苗同化枝葉綠素SPAD值較生境區(qū)A高出1.03,差異不顯著。圖4
表1 不同生境區(qū)生長期梭梭幼苗生長指標(biāo)的變化
Table 1 The growth indexes ofHaloxylonammodendronseedlings of different habitatsin growth period(cm)

生長期Growth period地點(diǎn)Sites株高Height冠幅Crown地徑Ground diameter梢長Shoots length4月April生境區(qū)A23.46a10.26a0.22a12.51a生境區(qū)B20.85b10.38a0.21a12.35a7月July生境區(qū)A32.52a21.65a0.29a18.72a生境區(qū)B28.88b19.73b0.28a17.12b8月August生境區(qū)A35.18a16.32a0.35a15.23a生境區(qū)B32.25b15.26ab0.32a14.58a11月November生境區(qū)A40.81a18.89a0.41a17.12a生境區(qū)B38.73b16.15b0.36a16.92ab
注:同一生長期的同列數(shù)據(jù)后的不同字母表示P<0.05水平差異顯著
Note: Values followed by different letters of the same indicator in a linear significant at the 5% level
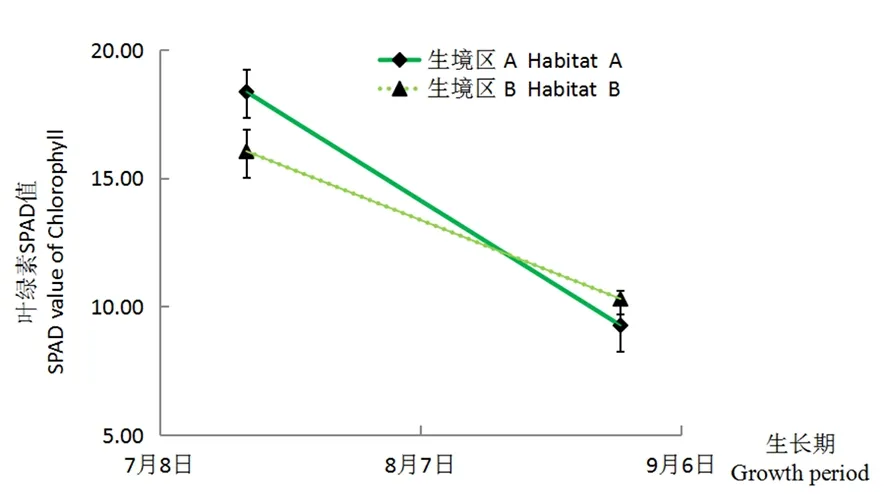
圖4 不同生境區(qū)梭梭幼苗的同化枝葉綠素SPAD值變化
Fig. 4 Characteristics of Chlorophyll SPAD value of assimilation branches in differentseedlings habitats
3 討 論
荒漠生態(tài)環(huán)境中,光照強(qiáng)度、土壤溫度和土壤水分是影響梭梭生長發(fā)育、生理代謝和形態(tài)建成的重要環(huán)境因子[17-19]。幼苗生境區(qū)A與B的地表光照強(qiáng)度的最大值均出現(xiàn)在7月,分別達(dá)到了78 500和82 000 lx,而土壤表層溫度的最大值均出現(xiàn)在8月,且隨著土層深度的增加呈持續(xù)降低趨勢,其地表最高溫度分別達(dá)到了46.28和47.60℃,小于俞闐等[20]在古爾班通古特沙漠南緣沙地夏季時的地表溫度值55℃,這可能與調(diào)查時期和調(diào)查當(dāng)天的氣溫差異有關(guān)。表明此階段幼苗生境區(qū)的光照強(qiáng)度和土壤溫度達(dá)到了全年最高水平。
水分是荒漠植物生存最重要的生態(tài)因子,不僅是植物體內(nèi)重要的物質(zhì)組成部分,而且維持著植物體內(nèi)各項生理活動的正常進(jìn)行[21-22]。生長期幼苗生境區(qū)A與B的水分豐富期為4和11月,這兩個時期有降水補(bǔ)充,地表蒸發(fā)較弱,其土壤表層水分含量的范圍分別為0.45%~4.16%和1.10%~6.87%;而水分匱乏期為7和8月,此時氣溫急劇升高,地表蒸發(fā)強(qiáng)烈,表層土壤水分含量急劇降低,其土壤水分含量范圍分別為0~1.88%和0~1.72%,大于田媛等[23]在古爾班通古特阜康荒漠區(qū)中的研究結(jié)果0~1.50%,這可能與兩地的降雨量和蒸發(fā)量不同有關(guān)。表明土壤表層水分狀況具有明顯的季節(jié)性,在生長中后期的7和8月,土壤表層水分條件處于全年最低水平,幼苗生境區(qū)生態(tài)環(huán)境條件變得日趨惡劣。
不同的光照強(qiáng)度、土壤溫度和土壤水分狀況提供給梭梭幼苗不同的生長環(huán)境。隨著生長期的推進(jìn),生境區(qū)A與B幼苗的株高和地徑呈遞增的變化趨勢,而冠幅和梢長呈先增加后降低再增加的變化趨勢,其中冠幅的最大值出現(xiàn)在7月,分別為21.65 和19.73 cm,小于楊海梅等[24]在古爾班通古特沙漠南緣莫索灣沙地上的研究結(jié)果32.13 cm,這可能與兩地的土壤養(yǎng)分和氣候條件有關(guān)。在極端干旱的7至8月。生境區(qū)A和B的幼苗同化枝葉綠素SPAD值分別由18.37、16.05下降到9.27、10.03。表明在生長中后期的7和8月,土壤表層水分條件是決定幼苗生長發(fā)育和更新的關(guān)鍵因素,由于土壤水分含量急劇降低,使得幼苗在適應(yīng)極端干旱環(huán)境中表現(xiàn)出特有的“夏休眠”現(xiàn)象,光合作用不斷減弱,同化枝葉綠素含量呈下降趨勢,生長緩慢,這與尹林克等[25]、李建貴等[26]的研究結(jié)論一致。
4 結(jié) 論
整個生長季,梭梭幼苗生境區(qū)A與B的地表光照強(qiáng)度、表層土壤溫度和土壤水分含量都具有明顯的季節(jié)性,其中,光照強(qiáng)度和土壤溫度呈先升高后降低的變化趨勢,最大值出現(xiàn)在7~8月間,且隨著土層深度的增加呈持續(xù)降低趨勢,表層土壤水分的匱乏期為7~8月,且隨著土層深度的增加呈持續(xù)升高趨勢。幼苗由于受到生境區(qū)生態(tài)因子季節(jié)性變化的影響,株高和地徑呈遞增的變化趨勢,冠幅和梢長呈先增加后降低再增加的變化趨勢,尤其在生長中后期7~8月,受到土壤水分含量急劇下降的影響,幼苗同化枝葉綠素SPAD值呈遞減的趨勢。通過不同生境區(qū)的光照強(qiáng)度、土壤溫度、土壤水分以及幼苗的生長生理適應(yīng)狀況的一致性和差異性對比分析,生境區(qū)A與B的生態(tài)因子均有利于幼苗的生長,但生境區(qū)A的幼苗生長狀況要優(yōu)于生境區(qū)B。
