黃花蒿醇提物對奶牛乳腺細胞中共軛亞油酸合成相關酶基因表達的作用
2019-10-10 06:00:10王麗芳張興夫
中國農業科學 2019年18期
關鍵詞:研究
王麗芳,張興夫
黃花蒿醇提物對奶牛乳腺細胞中共軛亞油酸合成相關酶基因表達的作用
王麗芳,張興夫
(內蒙古自治區農牧業科學院,呼和浩特 010031)
【】通過奶牛乳腺上皮細胞體外培養試驗,研究黃花蒿乙醇提取物對乳腺上皮細胞中與共軛亞油酸(CLA)合成相關酶的基因SCD表達的影響,探討其對乳脂肪合成相關酶ACACA和FASN基因以及與脂肪轉運相關酶LPL基因表達的影響,旨在從乳腺層次探明黃花蒿乙醇提取物對乳中CLA合成的部分作用機制。將采自于健康泌乳中期荷斯坦奶牛的乳腺組織用保溫盒帶回實驗室,在無菌條件選取腺泡較多的深層組織,采用膠原酶消化法獲得乳腺上皮細胞,放于37℃,CO2濃度為5%的培養箱中進行原代培養。試驗所用的細胞是經過復蘇的第二代細胞,待細胞復蘇后,以3×104個/mL的密度接種細胞到24孔板中,分別置于37 ℃的5% CO2培養箱中培養,采用臺盼藍計數法每天進行一次計數,設3個重復,連續7 d,未計數組每2天換液一次,繪制細胞生長曲線。另外,待細胞生長至對數增殖期,更換新鮮培養液,隨機分為4組,即培養液中黃花蒿提取物的濃度為0、3.0、6.0、12.0 mg·L-1,提取物作用時間均為48 h,檢測不同濃度黃花蒿提取物對與脂肪酸合成相關酶SCD、ACC、FAS、LPL 基因表達量的影響。每個處理3個重復。使用倒置顯微鏡觀察,乳腺上皮細胞形態呈鋪路石樣;在3×104個/mL的接種密度下,細胞生長曲線呈S型,在1—2d乳腺上皮細胞為潛伏期,3—6d為指數增長期,之后進入平臺期,符合一般細胞生長曲線規律,說明培養的乳腺上皮細胞具有正常的增殖能力,可以用于后續的研究;與對照組相比,添加黃花蒿乙醇提取物有增加SCD酶基因表達量的趨勢,其中3mg·L-1組顯著增加了SCD酶的基因表達量(<0.05),而6 mg·L-1組和12mg·L-1組雖然增加了SCD酶的基因表達量,但與對照組相比差異不顯著(>0.05);添加黃花蒿乙醇提取物有增加乳腺上皮細胞中ACACA基因表達量的趨勢,但與對照組相比差異不顯著(>0.05),且隨著添加劑量的增加,有下降趨勢;添加黃花蒿乙醇提取物可以增加乳腺上皮細胞中FASN基因表達量,呈劑量依賴型增加,且12mg·L-1組可以顯著增加乳腺上皮細胞中FASN基因表達量(<0.05);添加黃花蒿乙醇提取物有降低乳腺上皮細胞中LPL基因表達量的趨勢,呈劑量依賴型降低,且6 mg·L-1組和12mg·L-1組顯著降低了乳腺上皮細胞中LPL基因表達量(<0.05)。黃花蒿乙醇提取物可以增加奶牛乳腺上皮細胞中SCD、ACACA和FASN酶基因的表達,有利于調控CLA和乳脂肪的生成。
黃花蒿乙醇提取物;奶牛乳腺上皮細胞;SCD;ACACA;FASN;LPL
0 引言
【研究意義】共軛亞油酸(CLA)是一組亞油酸的幾何異構體和位置異構體的總稱,主要來源于反芻動物,在瘤胃氫化過程中形成,也可以通過trans-11C18:1轉化而成[1-2]。反式結構中含有雙鍵的異構體具有生物活性,研究最多的具有生物活性的CLA是cis-9,trans-11和trans-10,cis-12。CLA具有許多生物學作用,對人和動物健康影響的研究表明,CLA可能在癌癥、心血管疾病、肥胖、骨質疏松以及免疫和炎癥反應中發揮重要的保護作用[3-6]。因此增加反芻動物產品中CLA含量成為當前的研究熱點。【前人研究進展】Stefanie等[7]研究表明,奶牛日糧中添加亞麻籽可以增加乳中cis-9,trans-11CLA含量,其中以谷物青貯為粗飼料的日糧作為對照組,在此基礎上添加亞麻籽作為試驗組,試驗組中cis-9,trans-11CLA含量比對照組增加了126.92%,以牧草青貯為粗飼料的日糧中添加亞麻籽cis-9,trans-11CLA含量比對照組增加了132.69%。Bayat等[8]研究表明,奶牛日糧中添加紅花油和亞麻油分別極顯著和顯著增加了乳中cis-9,trans-11CLA含量,而添加菜籽油和肉豆蔻酸甲酯對乳中CLA含量沒有顯著影響;王麗芳[9]研究表明,與對照組相比,奶牛日糧中添加整粒亞麻籽和棉籽的混合物可以使乳中cis-9,trans-11CLA含量增加16.7%;也有研究報道,植物及其提取物可以增加羊奶、牛奶及乳腺上皮細胞中cis-9,trans-11CLA含量[10-15];另外,也有研究對乳中CLA含量的調控機制開展了相關研究。王麗芳等[16]通過奶牛飼養試驗,研究了黃花蒿乙醇提取物對瘤胃中氫化菌—溶纖維丁酸弧菌和蛋白溶解梭菌的影響;孫攀峰[17]通過體外批次培養試驗,研究了苜蓿、稻草和青貯玉米不同組合對瘤胃氫化菌—溶纖維丁酸弧菌和蛋白溶解梭菌的影響。【本研究切入點】乳中CLA主要來源于兩方面,一方面是亞油酸和亞麻酸在瘤胃中通過氫化菌氫化形成,另一方面是trans-11C18:1在乳腺中通過硬脂酰輔酶A去飽和酶SCD去飽和形成。上述研究多數集中于如何增加乳中CLA含量,少數研究從瘤胃氫化角度探討了CLA生成的調控機制,但是對于如何從乳腺層次研究CLA生成的調控機制卻鮮有報道。【擬解決的關鍵問題】通過乳腺上皮細胞體外培養技術,研究黃花蒿乙醇提取物對影響CLA合成的SCD基因表達的影響,同時探討對脂肪酸合成相關酶的基因表達的影響,旨在從乳腺層次探明黃花蒿乙醇提取物對乳中CLA合成的部分作用機制。
1 材料與方法
1.1 材料與儀器
1.1.1 黃花蒿乙醇取物 黃花蒿乙醇提取物購于南京澤朗公司,提取工藝條件是:乙醇濃度55%,提取溫度95℃,提取2 h。GC-MS和LC-MS分析的黃花蒿乙醇提取物活性成分主要包括:倍半萜類35%、芳香族類29%、脂肪酸類6%、甾體類6%、三萜類6%、脂肪族類4%、生物堿類3%、酚類5%、雜環類2%、其他4%(GC-MS);黃酮類和萜類(LC-MS)。
1.1.2 試劑 DMEM/F12培養基(Gibco,美國)、雙抗(Gibco,美國)、兩性霉素B(Gibco,美國)、胎牛血清(Gibco,美國)、胰島素轉鐵蛋白(Gibco,美國)、DPBS平衡鹽緩沖溶液(Hyclone,美國)、0.25%的胰蛋白酶(Gibco,美國)、二甲基亞砜(DMSO,Sigma,美國)、0.25%Trypsin-EDTA(Gibco 25200- 056)、膠原酶Ⅱ(Gibco 17101-015)、EGF(Gibco PHG-0313)、催乳素(Sigma L6520)、氫化可的松(Sigma H0135)、L-谷氨酰胺(Sigma G-8540)、SYBR Premix Ex TaqTMⅡ(TaKaRa DRR 081A)、PrimeScript RT Master Mix(TaKaRa DRR036A)、細胞總RNA提取試劑盒(TIANGEN DP430)。
1.1.3 儀器 電熱恒溫CO2培養箱(Heal Force HF240)、超凈工作臺(泰斯特)、電熱干燥箱、細胞培養用倒置相差顯微鏡(Olympus LX71)、普通冰箱(海爾)、超純水機(Purelab ELGA LA-611)、超低溫冰箱(Thermo)、電熱鼓風干燥箱(上海恒科)、電熱恒溫水浴鍋(北京光明儀器)、高壓蒸汽滅菌儀(HIRAYAMA HVE-50)、細胞計數儀(Cytorecon CYT-1000)、常溫低速離心機(Thermo)、磁力攪拌器、pH計、電子天平、熒光定量PCR儀(Bio-Rad)。
1.2 試驗方法
1.2.1 乳腺上皮細胞培養 乳腺上皮細胞培養方法參照文獻[18],將采自于健康泌乳中期荷斯坦奶牛的乳腺組織用保溫盒帶回實驗室,在無菌條件選取腺泡較多的深層組織,采用膠原酶消化法獲得乳腺上皮細胞,放于37℃,CO2濃度為5%的培養箱中進行原代培養。再利用成纖維細胞和乳腺上皮細胞對0.25%胰蛋白酶感受靈敏度的不同,通過調整消化時間來去除成纖維細胞,進而使乳腺上皮細胞得到純化。純化后的細胞凍于-80℃冰箱備用。試驗所用的細胞是經過復蘇的第二代細胞。
1.2.2 細胞生長曲線繪制 以3×104個/mL的密度接種細胞到24孔板中,分別置于37 ℃的5% CO2培養箱中培養,采用臺盼藍計數法每天進行一次計數,設3個重復,連續7d,未計數組每2d換液一次,繪制細胞生長曲線。
1.3 測定指標
1.3.1 脂肪酸合成相關酶的測定 待細胞生長至對數增殖期,更換新鮮培養液,隨機分為4組,即培養液中黃花蒿提取物的濃度為0、3.0、6.0、12.0 mg·L-1,提取物作用時間均為48 h,檢測不同濃度黃花蒿提取物對與脂肪酸合成相關酶SCD、ACC、FAS、LPL 基因表達量的影響。每個處理3個重復。
1.3.2 細胞總RNA提取 采用天根細胞RNA提取試劑盒(RNAprep pure Cell/Bacteria Kit ,DP430,離心柱型)提取細胞RNA。
1.3.3 反轉錄反應 按下列組配制RT 反應液(反應液的配制應在冰上進行):向200μL PCR管中依次加入5×PrimeScript Buffer 2μL、總RNA 2μL、Rnase Free dH2O 6μL;小心混勻;按以下條件進行反轉錄反應:37 ℃ 15 min ,85 ℃ 5 s ,4 ℃ 2 h ;得到的RT 反應液(cDNA)保存于4 ℃,用于后續的Real-Time PCR 反應。
1.3.4 Real-Time PCR 反應 在Bio-Rad IQ5 Real-Time PCR 儀上進行,試驗所用引物參考見下表1。選用管家基因β-Actin為內參。選用TaKaRa Real-Time PCR試劑盒(SYBR Premix Ex TaqTMⅡ),反應體系為20 μL:10 μL SYBR Premix Ex TaqTMⅡ(2×),0.4 μL上游引物,0.4μL下游引物,2 μL cDNA模板,7.2μL dH2O。反應程序為:95℃ 30 s ;95 ℃ 10 s;退火溫度下 20 s;72 ℃ 30 s;40個循環。熔解曲線程序為:95 ℃ 1 min;70 ℃ 1 min;溫度變化為0.5 ℃/6 s 。反應結束后,儀器自動生成關于Ct值的數據報告,再根據文獻采用2-ΔΔCt法計算得知各基因的相對表達量。
1.4 數據處理
試驗數據采用Excel 2007 整理后,根據文獻選用2-ΔΔCt法計算得知各基因的相對表達量[19],采用SAS9.0軟件中的ANOVA程序進行方差分析,均值多重比較采用Duncan法,<0.05為差異顯著水平。
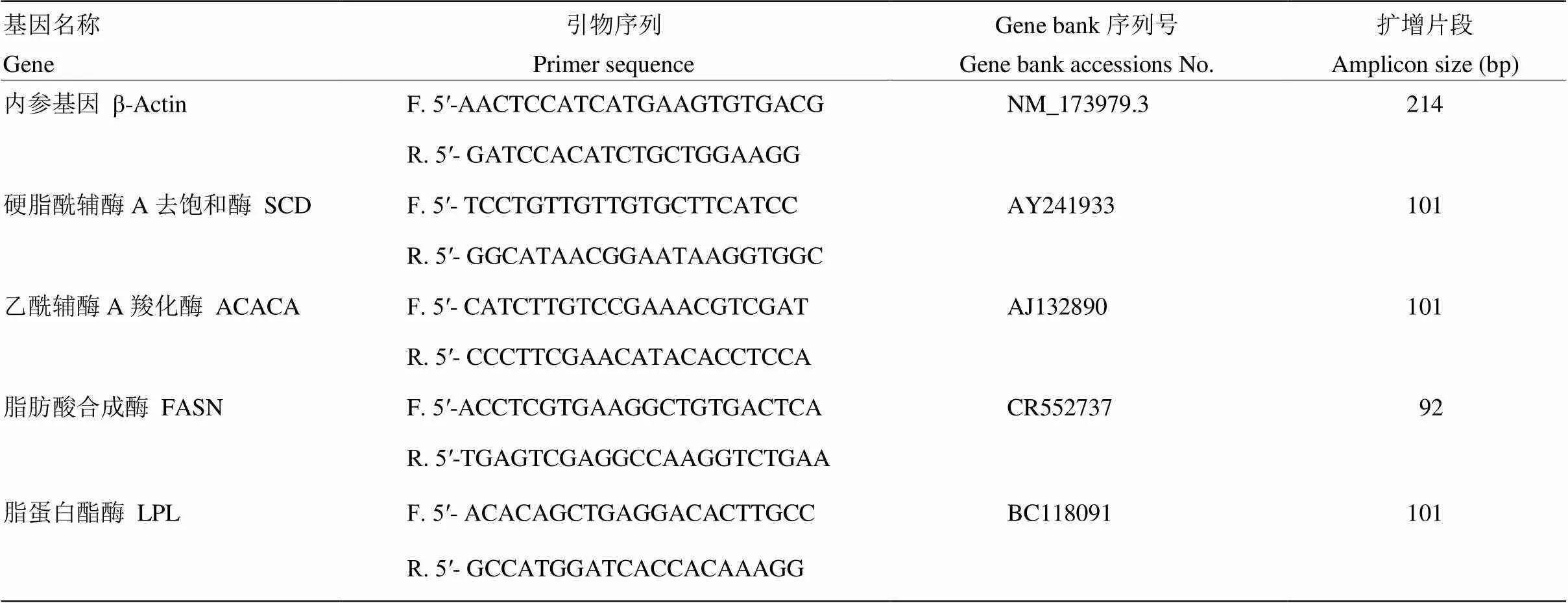
表1 脂肪酸合成相關酶基因的PCR引物序列
2 結果
2.1 乳腺上皮細胞生長曲線
從圖1可以看出,在3×104個/mL 的接種密度下,細胞生長曲線呈S型,在1—2d乳腺上皮細胞為潛伏期,3—6d為指數增長期,之后進入平臺期,符合一般細胞生長曲線規律,說明培養的乳腺上皮細胞具有正常的增殖能力,可以用于后續的研究。
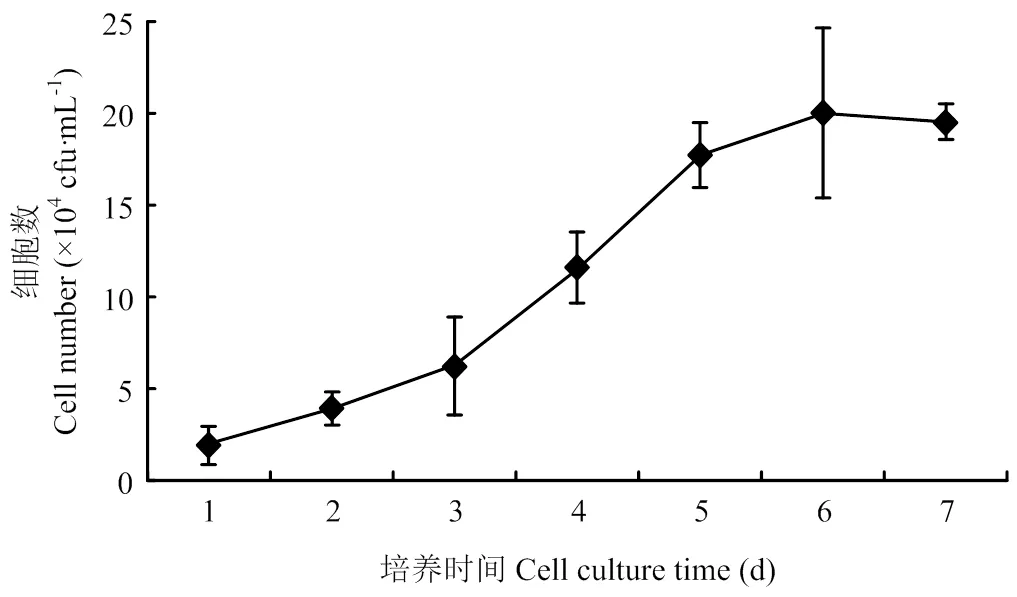
圖1 乳腺上皮細胞生長曲線
2.2 不同濃度黃花蒿醇提物對SCD基因表達的影響
從圖2可以看出,與對照組相比,添加黃花蒿乙醇提取物有增加SCD酶基因表達量的趨勢,其中3 mg·L-1組顯著增加了SCD酶的基因表達量(<0.05),而6 mg·L-1組和12mg·L-1組雖然增加了SCD酶的基因表達量,但與對照組相比差異不顯著(>0.05)。
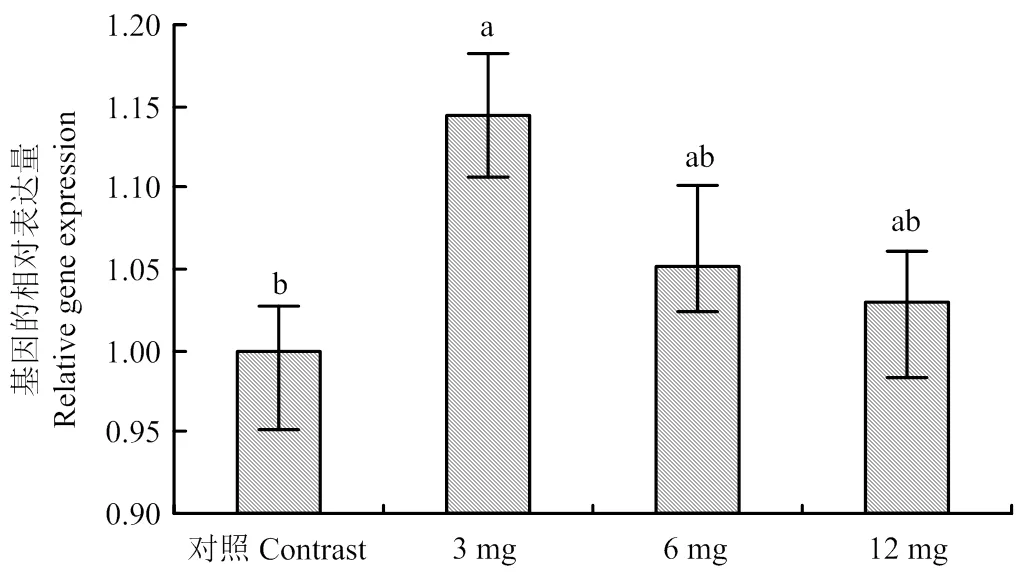
圖中標注不同小寫字母表示差異顯著(P<0.05)。下同
2.3 不同濃度黃花蒿醇提物對脂肪酸合成相關基因表達的影響
從圖3可以看出,添加黃花蒿乙醇提取物有增加乳腺上皮細胞中ACACA基因表達量的趨勢,但與對照組相比差異不顯著(>0.05),且隨著添加劑量的增加,有下降趨勢;添加黃花蒿乙醇提取物可以增加乳腺上皮細胞中FASN基因表達量,呈劑量依賴型增加,且12mg·L-1組可以顯著增加乳腺上皮細胞中FASN基因表達量;添加黃花蒿乙醇提取物有降低乳腺上皮細胞中LPL基因表達量的趨勢,呈劑量依賴型降低,且6 mg·L-1組和12mg·L-1組顯著降低了乳腺上皮細胞中LPL基因表達量(<0.05)。
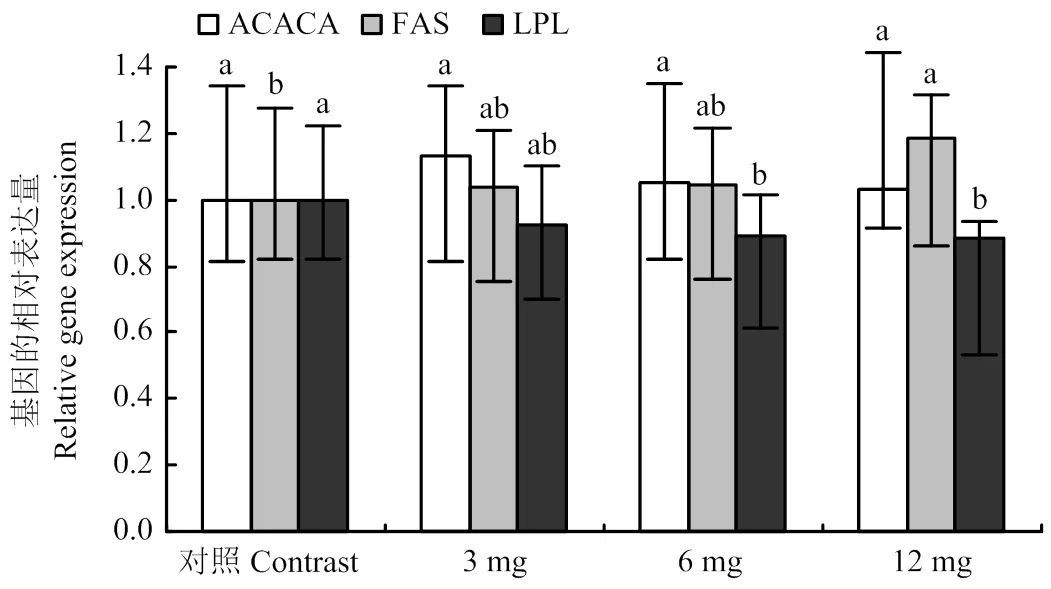
圖3 不同濃度黃花蒿醇提物對脂肪酸合成相關酶基因表達的影響
3 討論
3.1 黃花蒿醇提物對乳腺上皮細胞中SCD基因表達的影響
對于乳腺上皮細胞中SCD基因表達調控的研究多數集中于添加油酸、亞油酸、硬脂酸等游離脂肪酸。李大彪等[20]研究表明,與對照組相比,添加不同濃度亞油酸顯著降低了乳腺上皮細胞中SCD酶的基因表達,且呈劑量依賴型下降;王治國等[21]研究表明,棕櫚酸、硬脂酸、油酸、亞油酸對SCD基因的表達有抑制作用;YAO等[22]研究報道,亞油酸顯著降低SCD1基因的表達;研究報道,奶牛皺胃灌注或靜脈注射trans-10,cis-12 CLA可降低乳腺SCD1基因的轉錄[23-24];關于trans-10,cis-12 CLA對奶山羊SCD基因的影響,有研究報道十二指腸灌注trans-10,cis-12 CLA降低了乳脂肪中反應SCD酶基因的指數[25];上述研究均是關于不飽和脂肪酸對SCD基因的影響,INVERNIZZI等[26]研究報道,飽和長鏈脂肪酸或乳脂肪抑制劑可誘導牛乳腺組織中SCD1基因轉錄的持續上調;對于植物提取物影響乳腺中SCD基因表達的研究較少,王麗芳[11]研究報道,黃花蒿乙醇提取物可以降低奶山羊乳腺組織中SCD酶的基因表達。
本研究結果表明,黃花蒿醇提物有增加奶牛乳腺上皮細胞中SCD酶基因表達的趨勢,其中低劑量組顯著增加了乳腺上皮細胞中SCD酶的基因表達量。本研究結果與上述研究結果的作用機制可能不同,上述研究主要是脂肪酸對SCD酶基因表達的調控,本研究黃花蒿醇提物對SCD酶基因的表達調控可能主要是通過倍半萜類和芳香族類等多種活性成分;本研究結果與黃花蒿乙醇提取物對奶山羊乳腺組織中SCD酶基因表達的研究結果相反[11],原因可能是在研究黃花蒿乙醇提取物對奶山羊乳腺組織中SCD酶基因表達的影響時,奶山羊是活體動物,黃花蒿乙醇提取物進入奶山羊體內后在瘤胃微生物的作用下,經過了代謝轉化,到達乳腺組織中的物質已完全不同于黃花蒿乙醇提取物中的活性物質,本研究是直接在乳腺上皮細胞培養液中添加黃花蒿乙醇提取物,對乳腺上皮細胞作用的是黃花蒿乙醇提取物中原始的活性物質,另外,也可能是奶牛和奶山羊屬于不同種屬的奶畜,因此會存在差異。
3.2 黃花蒿醇提物對乳腺上皮細胞中脂肪酸合成相關酶基因表達的影響
ACACA和FASN是乳腺脂肪酸從頭合成的關鍵酶。首先,ACACA羧基化乙酰輔酶A形成丙二酰輔酶A,然后FASN利用乙酰輔酶A和丙二酰輔酶A合成脂肪酸[27],這兩種酶的合成受許多因素的影響。研究報道,熱應激會極顯著降低ACACA和FASN的合成(<0.01),添加精氨酸和蛋氨酸可以極顯著增加FASN的豐度(<0.01),但是對ACACA沒有顯著影響(>0.05)[28];ZHANG等[29]通過奶山羊乳腺上皮細胞體外培養試驗研究表明,高劑量trans-10,cis-12 CLA(200μmol·L-1)和低劑量cis-12 CLA(100μmol·L-1)均顯著降低了ACACA的mRNA豐度(<0.05),高劑量trans-10,cis-12 CLA顯著降低了FASN的蛋白表達(<0.05);李大彪等[20]研究表明,與對照組相比,添加不同濃度亞油酸顯著降低了乳腺上皮細胞中FASN酶的基因表達(<0.05);蘇芮等[30]研究表明,添加VA均增加了ACACA和FASN的基因表達。從上述研究結果可以看出,脂肪酸有降低ACACA和FASN基因表達的趨勢,氨基酸有增加FASN基因表達的趨勢,VA有增加ACACA和FASN基因表達的趨勢。本研究結果表明,添加黃花蒿乙醇提取物有增加FASN基因表達的趨勢,呈劑量依賴性增加,其中高劑量組(12mg·L-1)顯著增加了FASN基因的表達;添加黃花蒿乙醇提取物有增加ACACA基因表達的趨勢,但是與對照組差異不顯著。本研究結果與添加脂肪酸研究結果不同,可能原因是上述研究主要是脂肪酸對ACACA和FASN酶基因表達的調控,本研究黃花蒿醇提物對ACACA和FASN酶基因的表達調控可能主要是通過倍半萜類和芳香族類等多種活性成分,因此對ACACA和FASN的調控機制不同;本研究結果與VA和氨基酸的作用相似,可能本研究添加的植物提取物中所含有的活性成分結構與氨基酸和VA相似。
LPL是一種參與哺乳動物多種組織中長鏈脂肪酸LCFA 細胞內轉運的重要基因[31],其表達具有組織特異性。胡仕良[32]研究表明,奶山羊脂肪、心臟、肺臟等組織LPL基因的表達水平較高,乳腺組織LPL基因的表達水平較低。LPL mRNA的特異性表達受許多因素的影響,包括日糧、激素和禁食等,其中關于日糧調控LPL基因表達的研究較多。茅慧玲[33]研究表明,添加茶皂素和豆油可以顯著增加羔羊肌肉組織中LPL基因mRNA豐度(<0.05),但是對肝臟中LPL基因的表達卻沒有影響(>0.05);蘇芮等[30]研究表明,維生素A 對奶牛乳腺上皮細胞中乳脂合成相關酶LPL活性無顯著影響(>0.05);張博綦研究表明[34],LPS引起的奶牛乳腺上皮細胞氧化損傷可降低LPL的基因表達及其酶活性,但是添加硒可以有效減緩這種損傷作用;朱智[35]研究表明,添加不同濃度的大蒜油有增加奶山羊乳腺組織中LPL基因表達的趨勢。本研究結果表明,添加黃花蒿醇提物有降低奶牛乳腺上皮細胞中LPL基因表達的趨勢,與對照組相比,中高劑量組顯著降低了LPL的基因表達(<0.05)。本研究結果與上述研究結果不同,可能原因是本研究所添加的黃花蒿乙醇提取物所含有的活性成分與上述研究所添加的大蒜油等的活性成分不同相關。
4 結論
黃花蒿乙醇提取物可以增加奶牛乳腺上皮細胞中SCD、ACACA和FASN酶基因的表達,有利于調控共軛亞油酸和乳脂肪的生成。
[1] SCHMID A, COLLOMB M, SIEBER R, BEE G. Conjugated linoleic acid in meat and meat products: A review., 2006, 73:29-41.
[2] BAUMAN D E, PERFIELD J W I, HARVATINE K J, BAUMGARD L H.Regulation of fat synthesis by conjugated linoleic acid: lactation and the ruminant model.2008, 138: 403-409.
[3] BENJAMIN S, SPENER F.Conjugated linoleic acids as functional food: An insight into their health benefits.2009, 6:36.
[4] MORAES M L, RIBEIRO A M L, KESSLER A M, LEDUR V S, FISCHER M M, BOCKOR L, GAVA D. Effect of CLA on performance and immune response of weanling piglets., 2012, 90(8):2590-2598.
[5] OLIVEIRA D E, GAMA M A S, FEMANDES D, TEDESCHI L O, BAUMAN D E. An unprotected conjugated linoleic acid supplement decreases milk production and secretion of milk components in grazing dairy ewes., 2012, 95:1437-1446.
[6] DU R, ZHONG T, ZhANG W Q, SONG P, SONG W D, ZHAO Y, WANG C, TANG Y Q, ZHANG X, ZHANG Q. Antitumor effect of iRGD-modified liposomes containing conjugated linoleic acid- paclitaxel (CLA-PTX) on B16-F10 melanoma.2014, 9:3091-3105.
[7] ENGELKE S W, DAS G, DERNO M, TUCHSCHERER A, WIMMERS K, RYCHLIK M, KIENBERGER H, BERG W, KUHLA B, METGES C C. Methane prediction based on individual or groups of milk fatty acids for dairy cows fed rations with or without linseed., 2019,102(2) :1-15.
[8] BAYAT A R, TAPIO I, VILKKI J, SHINGFIELD K J, LESKINEN H. Plant oil supplements reduce methane emissions and improve milk fatty acid composition in dairy cows fed grass silage-based diets without affecting milk yield., 2018, 101(2) :1136-1151.
[9] 王麗芳. 奶牛日糧中添加整粒胡麻籽和棉籽對牛奶品質的影響[D]. 呼和浩特: 內蒙古農業大學, 2005.
WANG L F. Effect of linseed and cottonseed on milk quality in lactating dairy cows[D]. Hohhot: Inner Mongolia Agricultural University, 2005. (in Chinese)
[10] 斯琴畢力格, 王麗芳, 丁赫, 劉旺景, 連海飛, 敖長金. 飼糧中添加黃花蒿乙醇提取物對奶牛產奶性能及乳脂中共軛亞油酸含量的影響. 動物營養學報, 2017, 29(11): 4137-4142.
SI Q B L G, WANG L F, DING H, LIU W J, LIAN H F, AO C J. Effects of dietaryextracts on milk performance and conjugated linoleic acid content in milk fat of lactating cows. Chinese Journal of Animal Nutrition, 2017, 29(11): 4137-4142. (in Chinese)
[11] 王麗芳. 添加菊科植物黃花蒿提取物對奶山羊乳中CLA含量影響及機理研究[D]. 呼和浩特: 內蒙古農業大學, 2010.
WANG L F. The study on effects of compositaeextracts on cla content in goat milk and the relative mechanism[D]. Hohhot: Inner Mongolia Agricultural University, 2010. (in Chinese)
[12] 王麗芳, 盧德勛, 高民. 添加菊科植物黃花蒿乙醇提取物對奶牛乳腺上皮細胞脂肪酸組成的影響. 飼料工業, 2010, 31(19):36-38.
WANG L F, LU D X, GAO M. The effects of compositaeextracts on fatty acid composition of mammary epithelial cells in dairy cows., 2010, 31(19):36-38. (in Chinese)
[13] COLLOMB M, BUTIKOFER U, SIEBER R, JEANGROS B, BOSSET J O. Correlation between fatty acids in cows’ milk fat produced in the Lowlands, Mountains and Highlands of Switzerland and botanical composition of the fodder., 2002, 12(8): 661-666.
[14] CABIDDU A, ADDIS M, PINNA G, SPADA S, FIORI M, SITZIA M, PIRISI A, PIREDDA G, MOLLE G. The inclusion of a daisy plant () in dairy sheep diet. 1: Effect on milk and cheese fatty acid composition with particular reference to C18: 2 cis-9, trans-11., 2006, 101(1): 57-67.
[15] WALLACE R J. Biohydrogenation of fatty acids in the rumen//The 6th Joint Symposium of China-Korea-Japan on Rumen Metabolism and Physiology. 2007, 4.
[16] 王麗芳, 斯琴畢力格, 敖長金. 黃花蒿提取物對奶牛瘤胃發酵指標的影響. 中國農業科學, 2018, 51(23): 4548-4555.
WANG L F, SI Q B L G, AO C J. The Effects ofextracts on the rumen fermentation in dairy cows., 2018, 51(23): 4548-4555. (in Chinese)
[17] 孫攀峰.粗飼料組合對瘤胃發酵及乳脂肪酸組成的影響及其機理研究[D]. 杭州: 浙江大學, 2007.
SUN P F. Effect of Forage Combination on Ruminal Fermentation and Milk Fatty Acids Composition[D].Hangzhou: Zhejiang University, 2007. (in Chinese)
[18] 張興夫. 不同日糧模式對泌乳奶牛乳腺乳蛋白合成影響的研究[D]. 呼和浩特: 內蒙古農業大學, 2013.
ZHANG X F. Study on effects of different diets on milk protein synthesis in the mammary gland of dairy cows[D]. Hohhot: Inner Mongolia Agricultural University, 2013. (in Chinese)
[19] 塔娜, 李紅磊, 侯先志, 考桂蘭, 高民, 李大彪. 乙酸鈉和β-羥丁酸鈉對奶牛乳腺上皮細胞乳脂和乳蛋白合成相關基因表達的影響. 動物營養學報, 2014, 26(6): 1527-1534.
TA N, LI H L, HOU X Z, KAO G L, GAO M, LI D B. Effects of sodium acetate and sodium β-hydroxybutyrate on expressions of genes involved in milk fat and protein synthesis in bovine mammary epithelial cells., 2014, 26(6): 1527-1534. (in Chinese)
[20] 李大彪, 李紅磊, 邢媛媛, 于永強, 王衛云, 陳玲, 李平. 亞油酸對奶牛乳腺上皮細胞乳脂肪和乳蛋白合成相關基因表達的影響. 中國細胞生物學報,2016, 38(3): 257-264.
LI D B, LI H L, XING Y Y, YU Y Q, WANG W Y, CHEN L, LI P. Effect of linoleic acid on the expression of genes associated with milk fat and milk protein synthesis of bovine mammary epithelial cells., 2016, 38(3)::257-264.
[21] 王治國. 奶牛乳腺上皮細胞的體外培養及應用[D]. 北京:中國農業科學院,2007.
WANG Z G.culture and application of bovine mammary epithelial cell[D]. Beijing: Chinese Academy of Agricultural Sciences, 2007. (in Chinese)
[22] YAO D W, LUO J, HE Q Y, LI J, WANG H, SHI H B, XU H F, WANG M, LOOR J J. Characterization of the liver X receptor- dependent regulatory mechanism of goat stearoyl-coenzyme A desaturase 1 gene by linoleic acid., 2016, 99(5): 3945-3957.
[23] BAUMGARD L H, MATITASHVILI E, CORL B, DWYER D, BAUMAN D. Trans-10, cis-12 conjugated linoleic acid decreases lipogenic rates and expression of genes involved in milk lipid synthesis in dairy cows., 2002, 85(9): 2155-2163.
[24] GERVAIS R, MCFADDEN J W, LENGI A J, CORL B A, CHOUINARD P Y. Effects of intravenous infusion of trans-10, cis-12 18:2 on mammary lipid metabolism in lactating dairy cows., 2009, 92(10) :5167-5177.
[25] DEANDRADE P V, SCHMIDELY P. Effect of duodenal infusion of trans-10, cis-12-CLA on milk performance and milk fatty acid profile in dairy goats fed high or low concentrate diet in combination with rolled canola seed., 2006, 46(1):31-48.
[26] INVERNIZZI G, THERING B J, MCGUIRE M A, SAVOINI G, LOOR J J. Sustained upregulation of stearoyl-CoA desaturase in bovine mammary tissue with contrasting changes in milk fat synthesis and lipogenic gene networks caused by lipid supplements., 2010, 10(4):561-575.
[27] CHAJèS V, CAMBOT M, MOREAU K, LENOIR G M, JOULIN V. Acetyl-CoA carboxylase alpha is essential to breast cancer cell survival., 2006, 66(10): 5287-5294.
[28] SALAMA A A K, DUQUE M, WANG L, SHAHZAD K, OLIVERA M, LOOR J J. Enhanced supply of methionine or arginine alters mechanistic target of rapamycin signaling proteins, messenger RNA, and microRNA abundance in heat-stressed bovine mammary epithelial cells., 2019,102(3):1-12.
[29] ZHANG T Y, HUANG J T, TIAN H B, MA Y, CHEN Z, WANG J J, SHI H P, LUO J. Trans-10, cis-12 conjugated linoleic acid alters lipid metabolism of goat mammary epithelial cells by regulation of de novo synthesis and the AMPK signaling pathway., 2018, 101(6):5571-5581.
[30] 蘇芮, 劉陽, 閆素梅, 史彬林, 趙艷麗, 石惠宇. 維生素A對奶牛乳腺上皮細胞乳脂和乳蛋白合成相關基因表達的影響. 動物營養學報, 2018, 30(8):3151-3158.
SU R, LIU Y, YAN S M, SHI B L, ZHAO Y L, SHI H Y. Effects of Vitamin A on gene expressions related to milk fat and protein synthesis in bovine mammary epithelial cells., 2018, 30(8): 3151-3158. (in Chinese)
[31] BIONAZ M, LOOR J J. Gene networks driving bovine milk fat synthesis during the lactation cycle., 2008, 9: 366-387.
[32] 胡仕良. 西農薩能奶山羊乳腺上皮細胞LPL基因特征與功能的初步研究[D]. 楊凌: 西北農林科技大學, 2012.
HU S L. Primary characteristics and function study of LPL in epithelial cell in Xinong Saanen dairy goat[D]. Yangling: Northwest A and F University, 2012. (in Chinese)
[33] 茅慧玲. 植物源性物質對生長湖羊瘤胃發酵和肉品質的影響及其作用機理的研究[D]. 杭州: 浙江大學, 2010.
MAO H L. Effects of plants-based materials on growth performance, rumen fermentation and meat quality of growing Hu lambs[D]. Hangzhou: Zhejiang University, 2010. (in Chinese)
[34] 張博綦. 硒對奶牛乳腺上皮細胞內乳脂肪和乳蛋白合成相關基因表達的影響[D]. 呼和浩特: 內蒙古農業大學, 2017.
ZHANG B Q. Effects of selenium on gene expression related to milk fat and protein synthesis in bovine mammary epithelial cells [D]. Hohhot: Inner Mongolia Agricultural University, 2017. (in Chinese)
[35] 朱智. 大蒜油對山羊瘤胃生物氫化、乳脂肪酸組成及乳腺脂肪生成相關基因表達影響的研究[D].南京: 南京農業大學, 2011.
ZHU Z. Effects of garlic oil on ruminal biohydrogenation, milk fatty acid profile and lipogenesis-related gene expression in mammary gland of goats [D]. Nanjing: Nanjing Agricultural University, 2011. (in Chinese)
Effects ofExtracts on CLA Synthesis and Mechanism
WANG LiFang, ZHANG XingFu
(Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031)
【】The effects ofextracts (AAE) on gene SCD, ACACA, FASN and LPL related to CLA and milk fat synthesis in the bovine mammary epithelial cells(BMECs) culturewere studied in this paper. The purpose of this study was to explore the effects of AAE on the part mechanism of effecting CLA synthesis from the mammary.【】We isolated BMEC by using tissue fragments from the mammary gland of a healthy Holstein cows in Mid lactation. The cells used in the experiment were the second generation ones which have been resuscitated. The cells were inoculated into 24-well plates at a density of 3×104cells/mL and cultured at 37℃ in a 5% CO2atmosphere. The cells were counted by trypan blue counting method once a day. The experiment lasted for 7 d to draw the growth curve of the cells with three replicates, and the uncounted groups changed liquid every 2 d. When the cells grew to logarithmic proliferation stage, fresh culture medium was replaced and randomly divided into four groups, and the concentration of AAE in culture medium was 0, 3.0, 6.0, and 12.0 mg·L-1individually for 48 h. The effects of different concentration of AAE on the gene expression of SCD, ACC, FAS and LPL related to fatty acid synthesis were detected. Each treatment was repeated for three times. 【】The results showed as follows: the morphology and the growth curve for the BMECS looked like paving appearance and S-shaped respectively at 3×104cfu/mL. The latent period and the exponential growth period for the BMECS was 1-2 d and 3-6 d, respectively, and then in the plateau stage, which were accorded with the general cells growth curve rule and showed that the cultured mammary epithelial cells had normal proliferation ability as used in the subsequent researches. Compared with the control group, there was the trend that AAE supplementation increased the expression of SCD enzyme gene, 3 mg·L-1group significantly increased the expression of SCD gene (<0.05), while the 6 mg·L-1group and the 12 mg·L-1group increased the expression of SCD gene, but there was no significant difference compared with the control group (>0.05). The addition of AAE increased the expression of ACACA gene in mammary epithelial cells (>0.05). The addition of AAE increased the expression of FASN gene with a dose-dependent manner, the 12mg·L-1group increased the expression of FASN gene significantly (<0.05). There was the trend that AAE supplementation decreased the expression of LPL enzyme gene with a dose-dependent manner, and the 6 mg·L-1group and the 12 mg·L-1group decreased the expression of LPL gene significantly (<0.05). 【】It was concluded that AAE supplementation could increase the expression of SCD, ACACA and FASN genes in BMECS, which was beneficial to CLA synthesis.
extracts; BMECS; SCD; ACACA; FASN; LPL
10.3864/j.issn.0578-1752.2019.18.018

2019-01-24;
2019-08-16
國家自然科學基金(31860663)、內蒙古自治區自然科學基金(2013MS0418)、內蒙古農牧業科學院青年創新基金(2014QNJJM02)
王麗芳,Tel:13848189461;E-mail:wanglifang100008@163.com
(責任編輯 林鑒非)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
