丹參和藏丹參毛狀根MYB和bHLH轉(zhuǎn)錄因子基因表達(dá)差異研究
2019-09-24 01:01:28王艷婷郭妍宏王飛艷方譽(yù)民夏鵬國梁宗鎖楊東風(fēng)
浙江農(nóng)業(yè)科學(xué) 2019年9期
關(guān)鍵詞:差異
王艷婷,郭妍宏,王飛艷,方譽(yù)民,夏鵬國,梁宗鎖,楊東風(fēng)
(浙江理工大學(xué)生命科學(xué)與醫(yī)藥學(xué)院,浙江省植物次生代謝調(diào)控重點(diǎn)實(shí)驗(yàn)室,浙江 杭州 310018)
丹參是唇形科鼠尾草屬植物丹參(SalviamiltiorrhizaBunge)干燥根和根莖,具有保護(hù)心腦血管、抗腫瘤、抗氧化、抗炎、抗纖維化等作用[1-2]。藏丹參(SalviacastaneaDiels f.tomentosaStib)產(chǎn)于西藏林芝地區(qū),當(dāng)?shù)蒯t(yī)師把它的根和根莖作為丹參的替代品使用[3]。二氫丹參酮Ⅰ(dihydrotanshinone Ⅰ)、隱丹參酮(cryptotanshinone)、丹參酮Ⅰ(tanshinone Ⅰ)和丹參酮ⅡA(tanshinone ⅡA)等丹參酮和迷迭香酸(rosmarinic acid)、咖啡酸(caffeic acid)和丹酚酸B(salvianolicacid B)等酚酸是丹參的活性成分[4]。萜類化合物是植物次生代謝物質(zhì)中最大的一個(gè)家族,其生物合成途徑包括發(fā)生在胞質(zhì)的甲羥戊酸途徑(MVA)和發(fā)生在質(zhì)體的赤蘚糖磷酸途徑(MEP)[5]。酚酸生物合成與苯丙烷途徑和酪氨酸途徑有關(guān)。藏丹參和丹參活性成分相似,但含量差異較大。丹參的丹酚酸B含量更高,藏丹參丹參酮ⅡA和迷迭香酸的含量更高[6],然而兩種丹參活性物質(zhì)積累差異的形成機(jī)制尚不明確。
轉(zhuǎn)錄因子可以整合內(nèi)部(通常是發(fā)育)和外部(環(huán)境)信號(hào)來調(diào)節(jié)酶基因表達(dá),從而控制次級(jí)代謝物的特定積累[7]。MYB和bHLH轉(zhuǎn)錄因子廣泛參與苯丙烷類代謝途徑和萜類代謝途徑的調(diào)控,歐芹、玉米、金魚草、擬南芥、苦蕎麥和矮牽牛中黃酮類的合成均受MYB轉(zhuǎn)錄因子的調(diào)控[7-9],MYB轉(zhuǎn)錄因子也參與調(diào)控了丹參、擬南芥、火炬松中萜類化合物的合成[10-12];金魚草、龍膽中黃酮類的合成均受bHLH轉(zhuǎn)錄因子的調(diào)控[13-14];bHLH轉(zhuǎn)錄因子調(diào)控紅豆杉和丹參中萜類成分,長春花和黃連中生物堿類成分的積累,也已有報(bào)道[15-19]。本文通過比較紫花丹參和藏丹參毛狀根中MYB和bHLH基因表達(dá)的差異,分析可能參與調(diào)控丹參酮和丹酚酸的轉(zhuǎn)錄因子基因,以期揭示藏丹參和紫花丹參次生代謝的差異機(jī)制,為丹參酮和丹酚酸的合成及其調(diào)控奠定基礎(chǔ)。
1 材料與方法
1.1 材料與儀器
材料:發(fā)根農(nóng)桿菌ATCC15834侵染丹參和藏丹參無菌苗獲得丹參和藏丹參毛狀根(種質(zhì)來源由浙江理工大學(xué)生命科學(xué)學(xué)院梁宗鎖教授鑒定)。
基因和蛋白序列:GenBank數(shù)據(jù)庫中搜索(https://www.ncbi.nlm.nih.gov/)。
試劑:多糖多酚植物總RNA提取試劑盒(天根生化科技有限公司),反轉(zhuǎn)錄試劑盒 PrimeScriptTMRT reagent Kit和熒光定量PCR試劑盒SYBR Premix ExTaqTMⅡ(TaKaRa公司),熒光定量PCR引物由杭州擎科梓熙生物技術(shù)有限公司合成。
儀器:熒光定量PCR儀(QuantStudio 6 Flex,ABI)。
1.2 蛋白序列系統(tǒng)進(jìn)化分析
采用MEGA7.0中的NJ(Neighbor-joining)法構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹,參數(shù)選擇Bootstrap為1 000。
1.3 實(shí)驗(yàn)方法
1.3.1 丹參毛狀根培養(yǎng)
配制MS液體培養(yǎng)基,調(diào)pH值至5.8;0.2 g新鮮的毛狀根被轉(zhuǎn)至培養(yǎng)基中,在恒溫?fù)u床中暗培養(yǎng)(25 ℃、110 r·min-1)24 d后采樣。
1.3.2 基因表達(dá)分析
參照多糖多酚植物總RNA提取試劑盒說明書提取總RNA后進(jìn)行反轉(zhuǎn)錄(PrimeScriptTMRT reagent Kit)。采用SYBR Premix ExTaqTMⅡ進(jìn)行熒光定量PCR反應(yīng),引物序列見表1,SmActin基因用作內(nèi)參。反應(yīng)條件為:95 ℃、30 s;95 ℃、5 s,58 ℃、30 s,40個(gè)循環(huán)。基因表達(dá)分析用比較CT法(2-ΔΔCT)。
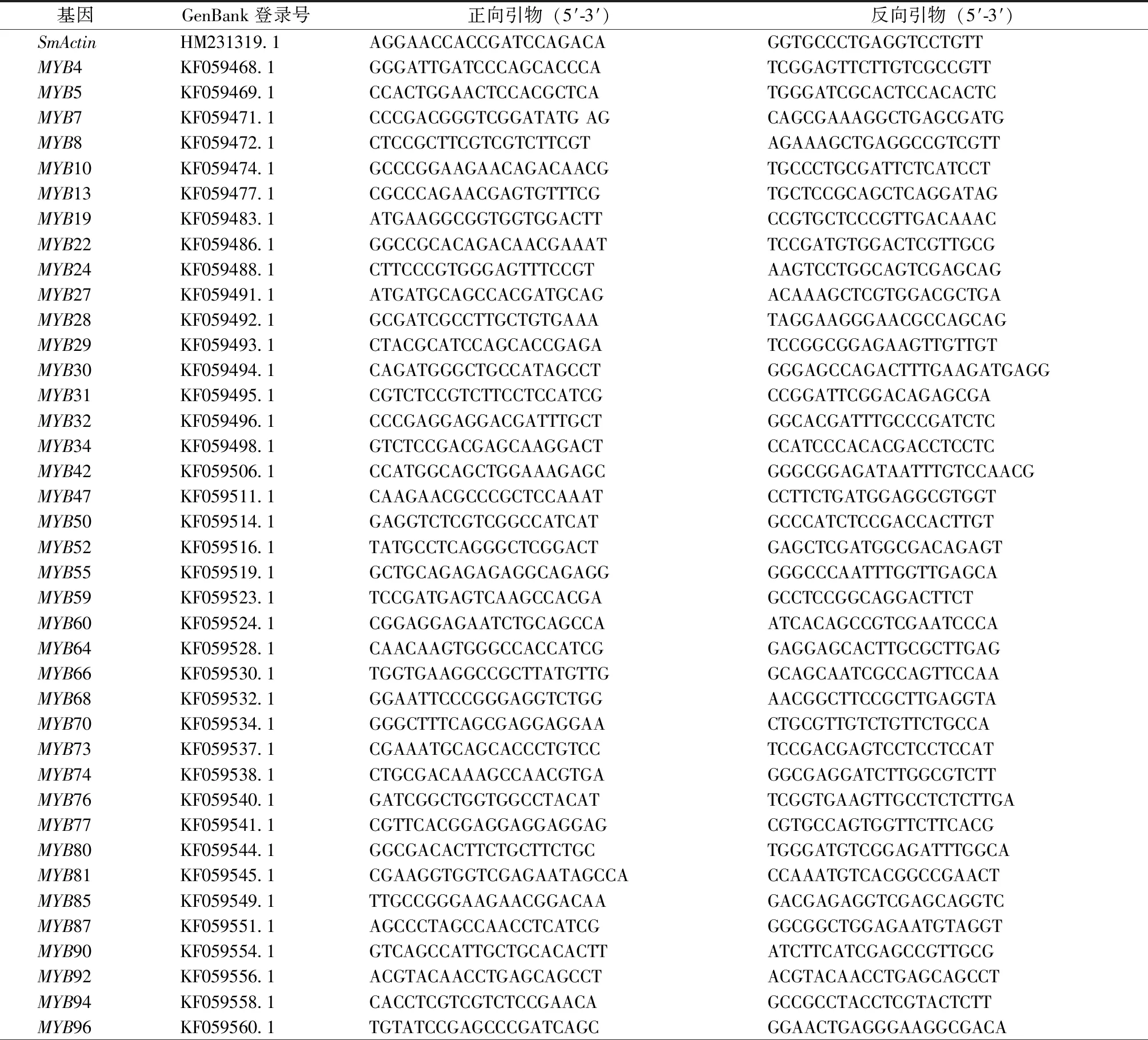
表1 實(shí)時(shí)熒光定量PCR引物

表1(續(xù))
1.3.3 數(shù)據(jù)分析
使用GraphPad Prism7作圖,數(shù)據(jù)以平均值±標(biāo)準(zhǔn)差表示,3個(gè)生物學(xué)重復(fù)。
2 結(jié)果與討論
2.1 藏丹參和丹參毛狀根中MYB表達(dá)差異
MYB轉(zhuǎn)錄因子不僅僅在植物細(xì)胞形態(tài)及模式建成、生長發(fā)育、逆境脅迫的應(yīng)答中發(fā)揮重要的作用,在生理活動(dòng)代謝、初生和次生代謝反應(yīng)的調(diào)節(jié)也扮演者重要角色[20]。自從第一個(gè)MYB轉(zhuǎn)錄因子MYBC1(和細(xì)胞色素合成有關(guān))在玉米中被鑒定[21],各類MYB基因從各種植物中分離鑒定。截至2019年3月17日,NCBI(National Center for Biotechnology Information)中MYB基因已至59 586條。MYB是調(diào)節(jié)丹參酮和丹酚酸的調(diào)控因子。為了研究不同的轉(zhuǎn)錄因子在藏丹參和丹參的表達(dá)差異,對(duì)47個(gè)MYB轉(zhuǎn)錄因子基因進(jìn)行了基因表達(dá)分析。結(jié)果表明,24個(gè)MYB(SmMYB60、SmMYB98、SmMYB87、SmMYB29等)在丹參中的表達(dá)顯著高于藏丹參,其中SmMYB60表達(dá)高達(dá)藏丹參的1 726.23倍,SmMYB98表達(dá)高達(dá)707.33倍(圖1中a)。SmMYB42、SmMYB34、SmMYB107、SmMYB85等4個(gè)MYB基因(圖1中c)在藏丹參中表達(dá)較高,分別高達(dá)丹參2.45、4.24、5.95、114.90倍。SmMYB96、SmMYB94、SmMYB80、SmMYB64等19個(gè)基因(圖1中b)表達(dá)在兩種丹參中表達(dá)差異不顯著。
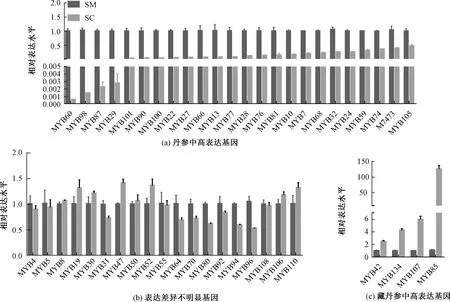
SM,丹參;SC,藏丹參。圖3同。圖1 MYB基因的表達(dá)模式
Li等[11]首次對(duì)丹參中MYB轉(zhuǎn)錄因子做了系統(tǒng)的分析,他們報(bào)道了110個(gè)R2R3-MYB基因,并且預(yù)測(cè)第4、5和20亞組的SmMYB可能參與調(diào)控了萜類化合物生物合成[11]。SmMYB36轉(zhuǎn)錄因子基因是一個(gè)典型的R2R3轉(zhuǎn)錄因子,SmMYB36含有第四和第五亞族特有的DNEI結(jié)構(gòu),團(tuán)隊(duì)前期克隆得到了SmMYB36轉(zhuǎn)錄因子基因,發(fā)現(xiàn)過表達(dá)SmMYB36促進(jìn)了丹參酮的積累,但抑制了丹參中酚酸和類黃酮的生物合成[22]。序列分析SmMYB36和SmMYB34進(jìn)化樹在同一分支(圖2),實(shí)驗(yàn)表明,SmMYB34在藏丹參中的表達(dá)量是丹參的4.43倍,SmMYB34可能與藏丹參對(duì)丹參酮的高積累有關(guān)。SmMYB39轉(zhuǎn)錄因子基因[23]和Li等[11]注釋的SmMYB76基因?qū)儆谕粭l基因,為第4亞組[11],研究表明,SmMYB76可以抑制丹酚酸的積累。在本研究中SmMYB76和SmMYB28在同一分支(圖2),與前人研究一致[11],本實(shí)驗(yàn)中它在丹參中的表達(dá)量是藏丹參的8.18倍,這很可能與藏丹參低丹酚酸B積累有關(guān)。
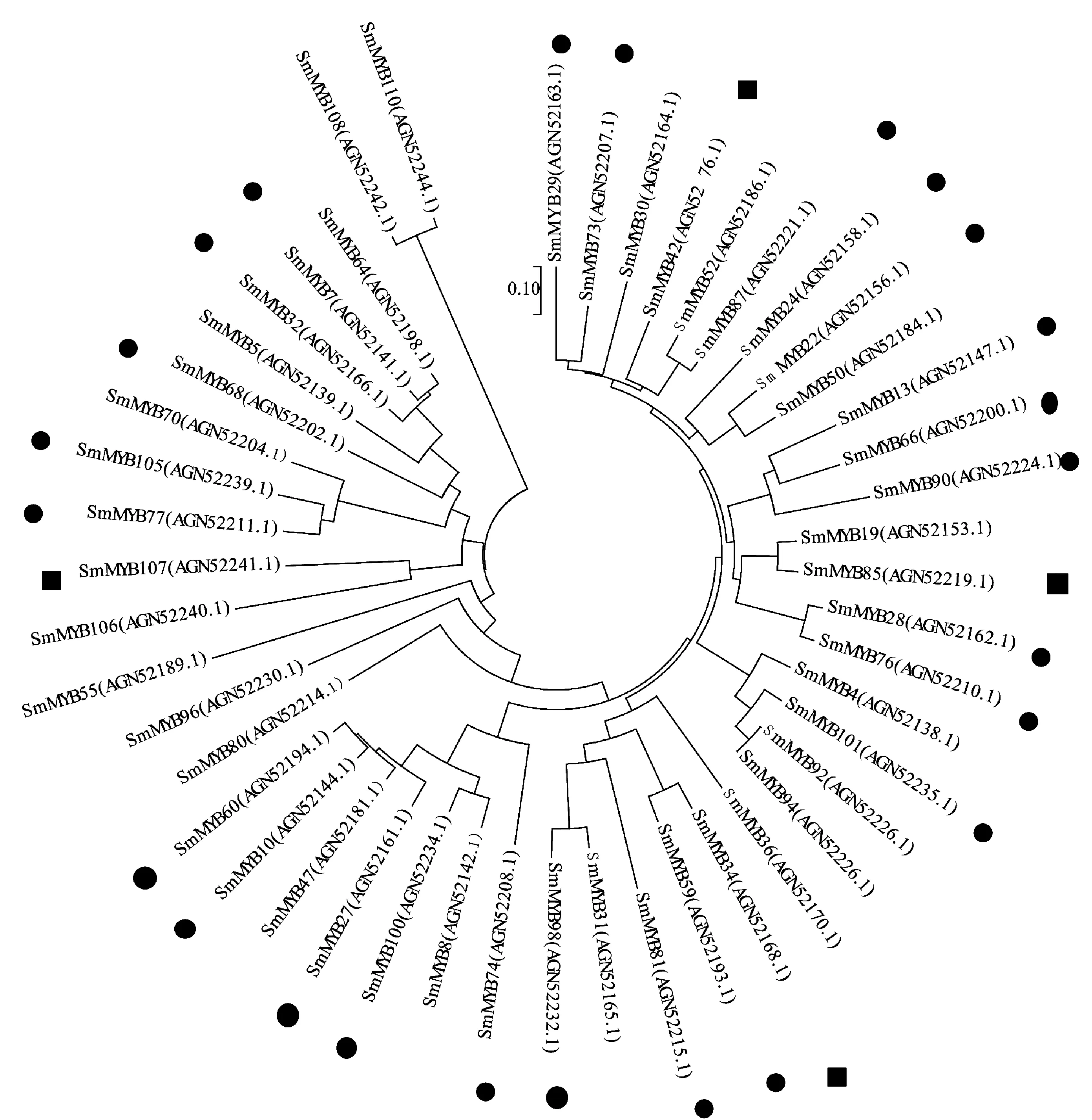
●丹參中表達(dá)量較高;■藏丹參中表達(dá)量較高。圖4同。圖2 丹參MYB轉(zhuǎn)錄因子系統(tǒng)進(jìn)化樹
丁慶倩等[24]從大豆中鑒定出一個(gè)在脅迫條件下明顯上調(diào)的MYB類轉(zhuǎn)錄因子SiMYB42。藏丹參(海拔2 900~3 500 m林芝地區(qū))相對(duì)丹參(平原)面臨更多的高UV-B輻射等逆境壓力[25]。研究發(fā)現(xiàn),酵母提取物和水解乳蛋白可以提高藏丹參和丹參中丹參酮的含量,并且藏丹參對(duì)酵母提取物的影響反應(yīng)更大[26-27]。在玉米種MYB42與木質(zhì)素含量的負(fù)調(diào)節(jié)有關(guān)[28],木質(zhì)素和酚酸生物合成都源于苯丙烷途徑。在本實(shí)驗(yàn)中SmMYB42在藏丹參中的表達(dá)量是丹參的2.45倍,根據(jù)這些,推測(cè)SmMYB42在酚酸的積累過程發(fā)揮著重要的作用。
研究發(fā)現(xiàn),過表達(dá)SmMYB98b毛狀根株系中丹參酮含量和丹參酮合成通路關(guān)鍵酶基因的表達(dá)都有所提高。干擾SmMYB98b毛狀根株系中丹參酮含量和丹參酮合成通路關(guān)鍵酶基因的表達(dá)都被抑制[29]。在本實(shí)驗(yàn)中在丹參中的表達(dá)量高達(dá)藏丹參的707.33倍,SmMYB98可能酚酸的積累過程也扮演重要的角色。在草莓中FaMYB10正調(diào)節(jié)花青素的積累[30],花青素等酮類的合成與酚酸的合成都依賴于PAL途徑,本實(shí)驗(yàn)中SmMYB10在丹參中的表達(dá)量高達(dá)藏丹參的6.12倍,MYB10轉(zhuǎn)錄因子基因也可能是酚酸積累的一個(gè)重要調(diào)控因子。在蘋果中MYB22過表達(dá)株系花青素的積累和CHS、CHI、F3H等花青素通路關(guān)鍵酶基因的表達(dá)都被抑制,本實(shí)驗(yàn)中SmMYB22在丹參中的表達(dá)量高達(dá)藏丹參的16.79倍,SmMYB22可能與藏丹參高迷迭香酸積累有關(guān)。
2.2 藏丹參和丹參毛狀根bHLH表達(dá)差異
bHLH轉(zhuǎn)錄因子家族是植物中最大的轉(zhuǎn)錄因子家族之一,參與調(diào)控了黃酮類化合物,生物堿和萜類化合物的生物合成。已發(fā)現(xiàn)10 907多個(gè)bHLH轉(zhuǎn)錄因子,已經(jīng)完成系統(tǒng)性鑒定和分類的有擬南芥(Arabidopsisthaliana)、水稻(Oryzasativa)和煙草(NicotianatabacumL.)等[31-33]。為了研究不同的轉(zhuǎn)錄因子在藏丹參和丹參的表達(dá)差異,對(duì)47個(gè)bHLH轉(zhuǎn)錄因子進(jìn)行了分析。結(jié)果表明,15個(gè)SmbHLH(SmbHLH69、SmbHLH33、SmbHLH123、SmbHLH96等)在丹參中的表達(dá)量顯著高于藏丹參,其中SmbHLH69表達(dá)高達(dá)藏丹參的1 727.61倍,SmbHLH33高達(dá)29.72倍(圖3中b)。SmbHLH48、SmbHLH46、SmbHLH79、SmbHLH108、SmbHLH73和SmbHLH77等6個(gè)bHLH基因(圖3中c)在藏丹參中表達(dá)量較高,分別高達(dá)丹參中表達(dá)量的2.51、3.36、8.95、12.36、34.02和124.67倍。SmbHLH121、SmbHLH10、SmbHLH29、SmbHLH26等26個(gè)基因(圖3中a)在兩種丹參中表達(dá)量差異不顯著。
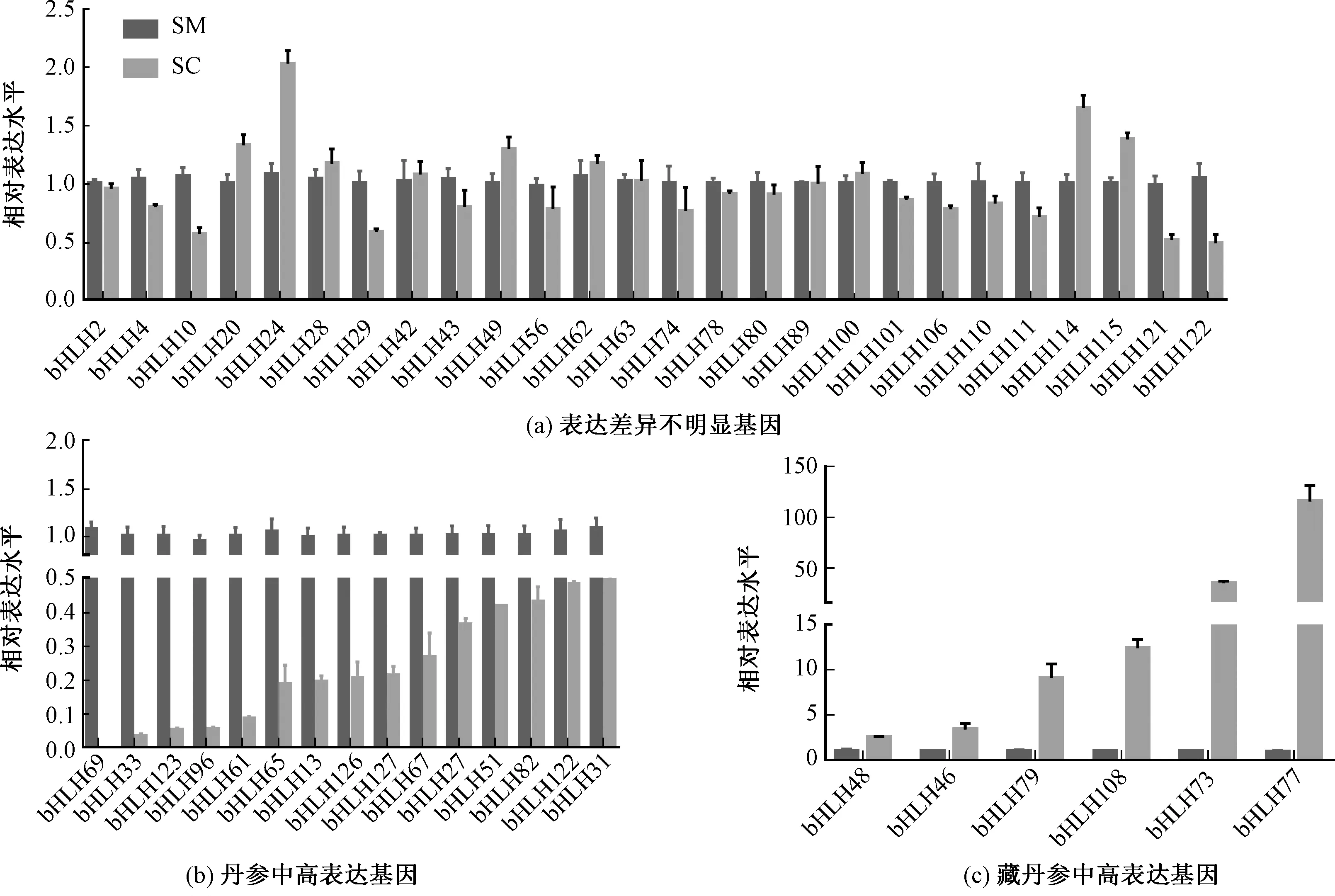
圖3 bHLH 基因的表達(dá)模式
Zhang等[34]從丹參基因組中篩選出127個(gè)bHLH轉(zhuǎn)錄因子并進(jìn)行了系統(tǒng)的分析。他們預(yù)測(cè)7個(gè)可能調(diào)節(jié)丹參酮生物合成的基因,分別為SmbHLH37(亞族R)、SmbHLH51(亞族R)、SmbHLH53(亞族R)、SmbHLH60(亞族W)、SmbHLH74(亞族H)、SmbHLH92(亞族P)和SmbHLH103(亞族P)。丹參SmbHLH10轉(zhuǎn)錄因子基因,它和SmbHLH83基因?qū)儆谕粭l基因[35-36],前期研究表明,SmbHLH83可以促進(jìn)丹參酮的的積累[23],在本研究的進(jìn)化樹中單獨(dú)一個(gè)分支(圖4),但它在藏丹參和丹參中表達(dá)量差異不顯著,推測(cè)SmbHLH83可能不是藏丹參和丹參次生代謝產(chǎn)物積累差異的主要原因。前人的研究表明,SmbHLH48和SmbHLH74同屬亞族H[34],bHLH48和SmbHLH74序列相似,并且bHLH48在藏丹參中的表達(dá)量是丹參的2.51倍,bHLH48可能和藏丹參高丹參酮積累有關(guān)。
擬南芥MYC2[35]調(diào)節(jié)倍半萜合酶基因表達(dá),并且丹參酮和倍半萜烯都是萜類化合物[35]。構(gòu)建鄰接系統(tǒng)發(fā)育樹時(shí)發(fā)現(xiàn)bHLH51[34]和NtMYC2a,NtMYC2b和CrMYC2在相同的亞家族,并且SmbHLH51在根中表達(dá)水平比在其他3個(gè)器官的表達(dá)水平高,在用茉莉酸處理后呈上調(diào)的趨勢(shì)。SmbHLH51在丹參中的表達(dá)量是藏丹參的2.78倍,SmbHLH51在丹參酮生物合成中具有一定的調(diào)節(jié)作用。梨的紅色是花青素積累的結(jié)果,SmbHLH33參與花色素苷生物合成的差異調(diào)節(jié)[36],定量結(jié)果顯示SmbHLH33在丹參中的表達(dá)量高達(dá)藏丹參的29.72倍,苯丙烷代謝也是黃酮類化合物(例如花青素、黃酮、黃酮醇及其糖苷)的上游途徑,花青素都與酚酸生物合成共享相同的前體[37],SmbHLH33可能參與迷迭香酸和丹酚酸B的生物合成。擬南芥中miR393靶向轉(zhuǎn)錄因子bHLH77和生長素受體TIR1,AFB(1,2,3)[38],生長素在植物的生長發(fā)育中發(fā)揮著重要的作用,SmbHLH77在藏丹參中的表達(dá)量高達(dá)丹參的155.41倍,可能間接的調(diào)節(jié)丹參次生代謝物質(zhì)的積累。AtbHLH122轉(zhuǎn)錄因子可以調(diào)控ABA的積累來參與植物適應(yīng)逆境脅迫。有研究表明,施用適量的赤霉素能促進(jìn)丹參的生長和丹參根中丹參酮類物質(zhì)的積累。SmbHLH122在丹參中表達(dá)量僅僅是藏丹參的2.16倍,暗示轉(zhuǎn)錄因子可能不是藏丹參高丹參酮積累的關(guān)鍵轉(zhuǎn)錄因子[39]。
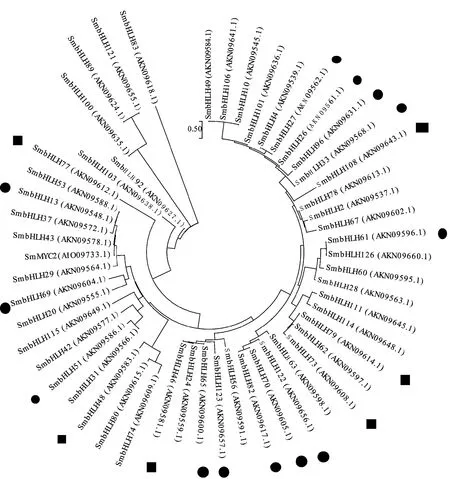
圖4 丹參bHLH轉(zhuǎn)錄因子系統(tǒng)進(jìn)化樹
3 小結(jié)
本研究采用熒光定量PCR分析了MYB和bHLH轉(zhuǎn)錄因子在藏丹參與丹參毛狀根中的表達(dá)差異,結(jié)果表明:有24個(gè)MYB(SmMYB60、SmMYB98、SmMYB87、SmMYB29等)和15個(gè)SmbHLH(SmbHLH69、SmbHLH33、SmbHLH123、SmbHLH96等)在丹參中高表達(dá),有4個(gè)MYB(SmMYB96、SmMYB94、SmMYB80、SmMYB64)和6個(gè)SmbHLH(SmbHLH48、SmbHLH46、SmbHLH79、SmbHLH108、SmbHLH73和SmbHLH77)在藏丹參中高表達(dá)。這些差異表達(dá)的轉(zhuǎn)錄因子可能參與調(diào)控了丹參酮和丹酚酸的積累,導(dǎo)致藏丹參和丹參次生代謝的差異。但是這些轉(zhuǎn)錄因子是如何參與代謝目前還不太清楚,需要做進(jìn)一步的研究。本研究為揭示丹參和藏丹參的差異以及丹酚酸、丹參酮的生物合成機(jī)制奠定了基礎(chǔ)。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
