植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
2019-09-10 07:22:44張婷婷劉文耀黃俊彪胡濤湯丹丹陳泉
廣西植物 2019年5期
關(guān)鍵詞:植物
張婷婷 劉文耀 黃俊彪 胡濤 湯丹丹 陳泉

摘 要:化學(xué)計(jì)量?jī)?nèi)穩(wěn)性是生態(tài)化學(xué)計(jì)量學(xué)研究的核心概念之一,是指生物在面對(duì)外界變化的時(shí)候保持自身化學(xué)組成相對(duì)穩(wěn)定的能力,其反映了生物對(duì)周?chē)h(huán)境變化作出的生理和生化響應(yīng)與適應(yīng)。通過(guò)研究植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性,有助于深入了解植物對(duì)環(huán)境的適應(yīng)策略和生態(tài)適應(yīng)性,以及植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性與生態(tài)系統(tǒng)功能的關(guān)系,但目前關(guān)于植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究較少。已有的研究結(jié)果表明:不同物種或功能群由于其生長(zhǎng)策略不同而具有不同的生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征;同一物種的不同器官、不同生長(zhǎng)階段以及不同元素的內(nèi)穩(wěn)性存在較大的差異。該文對(duì)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性概念、內(nèi)穩(wěn)性指數(shù)的測(cè)算方法,不同植物物種或功能群、不同器官、不同生長(zhǎng)階段內(nèi)穩(wěn)性特征,以及植物內(nèi)穩(wěn)性與生態(tài)系統(tǒng)結(jié)構(gòu)、功能和穩(wěn)定性的關(guān)系等方面進(jìn)行了綜述,并結(jié)合現(xiàn)已開(kāi)展的工作,對(duì)有待進(jìn)一步拓展的相關(guān)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究領(lǐng)域進(jìn)行了展望,以期為促進(jìn)國(guó)內(nèi)相關(guān)研究工作的開(kāi)展提供參考。
關(guān)鍵詞: 生態(tài)化學(xué)計(jì)量學(xué), 內(nèi)穩(wěn)性, 植物, 生態(tài)系統(tǒng)穩(wěn)定性
中圖分類(lèi)號(hào):Q948
文獻(xiàn)標(biāo)識(shí)碼:A
文章編號(hào):1000-3142(2019)05-0701-12
Characteristics of plant ecological stoichiometry homeostasis
ZHANG Tingting1,2, LIU Wenyao1*, HUANG Junbiao1,2,HU Tao1,2, TANG Dandan1,2, CHEN Quan1,2
( 1. CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences,Kunming 650223, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China )
Abstract:Stoichiometric homeostasis is one of the core concepts of ecological stoichiometry. It refers to the ability of an organism to maintain a given elemental composition in the body despite variation in the elemental composition of its environment or diet. It reflects the net outcome of many underlying physiological and biochemical adjustments as organisms respond to their surroundings. Studies of stoichiometric homeostasis of plant will help us to understand the adaptation strategies and ecological adaptabilites of plants to the environment, as well as the relationships between plant ecological stoichiometry homeostasis and ecosystem functions. However, there are few studies on the internal stability of plant ecological stoichiometry. Previous research showed that the stoichiometric homeostasis characteristics of different plant species or functional groups were different. There were differences in stoichiometric homeostasis among different growth stages, different organs of the same species, as well as different nutrient elements. This paper reviews the concept of stoichiometric homeostasis, the estimation of plant homeostatic index, the stoichiometric homeostasis characteristics of different plant species or functional groups, different organs and growth stages and different elements, as well as linking plant stoichiometric homoeostasis with ecosystem structure, functioning and stability. Based on the obtained achievements and the current study, we put forward some perspectives of plant stoichiometric homoeostasis for future researches to be conducted with an aim to promote this discipline of research in China.
Key words: ecological stoichiometry, homeostasis, plant, ecosystem stability
生態(tài)化學(xué)計(jì)量學(xué)是一門(mén)研究生態(tài)交互過(guò)程中能量以及多種化學(xué)元素平衡的學(xué)科 (Elser et al., 2000)。它不僅將生物學(xué)、生態(tài)學(xué)、化學(xué)、物理學(xué)、化學(xué)計(jì)量學(xué)等不同學(xué)科的基本原理結(jié)合起來(lái),還涵蓋了熱力學(xué)第一定律、生物進(jìn)化的自然選擇原理和分子生物學(xué)中心法則理論,從而使生物學(xué)科從分子、細(xì)胞、有機(jī)體、種群、群落、生態(tài)系統(tǒng)等不同尺度的研究理論都能夠有機(jī)地統(tǒng)一起來(lái) (曾徳慧和陳廣生,2005;王紹強(qiáng)和于貴瑞,2008;賀金生和韓興國(guó),2010)。生物體是由化學(xué)元素組成的,生態(tài)系統(tǒng)中不同生物體的交互作用都涉及化學(xué)元素的重組。生物體的C、N、P化學(xué)計(jì)量特征與其自身的穩(wěn)定性、群落結(jié)構(gòu)、生態(tài)系統(tǒng)的穩(wěn)定性以及生物地球化學(xué)循環(huán)密切相關(guān) (Loladze et al., 2000; Sterner & Hessen, 1994; Elser et al., 2009)。
生物有機(jī)體元素組成的內(nèi)穩(wěn)性理論是生態(tài)化學(xué)計(jì)量學(xué)的核心概念之一 (Persson et al., 2010),其強(qiáng)弱與物種的生態(tài)策略和適應(yīng)性有關(guān) (Jeyasingh et al., 2009)。從國(guó)內(nèi)外研究來(lái)看,化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究屬于一個(gè)新興的領(lǐng)域,研究成果也非常有限,化學(xué)計(jì)量?jī)?nèi)穩(wěn)性是生態(tài)學(xué)研究中一個(gè)被低估的焦點(diǎn)(Giordano, 2013)。生物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究的對(duì)象由最初的真菌和細(xì)菌(Levi & Cowling, 1969; Goldman et al., 1987)發(fā)展到藻類(lèi)、浮游動(dòng)物和草本植物(Rhee, 1978; Andersen & Hessen, 1991; Güsewell, 2005)。由于高等植物體內(nèi)元素的內(nèi)穩(wěn)性和化學(xué)計(jì)量特征復(fù)雜,對(duì)高等植物的研究甚少(Cernusak et al., 2009; Matzek & Vitousek, 2009; Yu et al., 2010; 蔣利玲等,2017)。早期的化學(xué)計(jì)量學(xué)理論認(rèn)為,細(xì)菌和多細(xì)胞動(dòng)物的化學(xué)計(jì)量具有絕對(duì)的內(nèi)穩(wěn)性,而光合自養(yǎng)生物 (藍(lán)細(xì)菌、藻類(lèi)、植物)的內(nèi)穩(wěn)性非常弱 (Elser et al., 2010)。總體上,從早期的原核生物到后期的原核生物,再到單細(xì)胞真核生物和多細(xì)胞真核生物,內(nèi)穩(wěn)性可能是逐漸增強(qiáng)的 (Williams & DaSilva, 1996)。一方面,藻類(lèi)和真菌的內(nèi)穩(wěn)性低于低等植物,低等植物低于高等植物,植物低于動(dòng)物 (Sterner & Elser, 2002);另一方面,不同元素的內(nèi)穩(wěn)性也不相同,大量元素的內(nèi)穩(wěn)性高于微量元素,微量元素高于非必要元素 (Karimi & Folt, 2006)。植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性受養(yǎng)分供應(yīng)狀況 (嚴(yán)正兵等,2013)、光強(qiáng)以及施肥 (Sterner & Elser, 2002; Limpens & Berendse, 2004)、植物器官、生長(zhǎng)發(fā)育階段和元素 (Yu et al., 2011)的影響。
生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性能夠較好地反映生物對(duì)環(huán)境變化的生理和生化的適應(yīng) (Hessen et al., 2004; Elser et al., 2010),其內(nèi)穩(wěn)性的強(qiáng)弱與物種的生態(tài)策略和適應(yīng)性有關(guān) (Yu et al., 2011)。生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性調(diào)節(jié)作用反映了植物的生理和生化分配作用對(duì)其外部環(huán)境的響應(yīng),并通過(guò)根系對(duì)N、P等元素的吸收與釋放完成這種調(diào)節(jié)作用 (Güsewell, 2004)。N元素對(duì)植物生長(zhǎng)的最重要影響包括調(diào)節(jié)器官的數(shù)量和大小以及平衡營(yíng)養(yǎng)和生殖生長(zhǎng),P元素主要影響葉片的形成及其形狀,調(diào)控植物開(kāi)花和種子的形成 (Plénet et al., 2000; Marschner, 2012)。在內(nèi)蒙古羊草草原,生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性高的物種具有較高的優(yōu)勢(shì)度和穩(wěn)定性 (Yu et al., 2010),但環(huán)境條件 (如氮肥添加) 可能會(huì)改變生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性與生態(tài)系統(tǒng)特性的關(guān)系 (Bai et al., 2010)。由于不同的生態(tài)系統(tǒng)類(lèi)型、不同植物群落類(lèi)型之間存在巨大差異,生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性在一些特殊的生態(tài)系統(tǒng)和群落,如濕地生態(tài)系統(tǒng)、林冠附生生物群落等中的特征亟待研究。
雖然國(guó)內(nèi)外有關(guān)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究還不多,但生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的生態(tài)學(xué)和進(jìn)化學(xué)意義已得到學(xué)者們的高度重視 (Sterner & Elser, 2002; Elser et al., 2010; Yu et al., 2010, 2011; 曾冬萍等,2013;蔣利玲等,2017)。有關(guān)研究表明,物種水平的內(nèi)穩(wěn)性與物種優(yōu)勢(shì)度和穩(wěn)定性正相關(guān),而且在多數(shù)情況下,群落水平的內(nèi)穩(wěn)性與生態(tài)系統(tǒng)功能以及穩(wěn)定性也正相關(guān) (Yu et al., 2015)。內(nèi)穩(wěn)性可作為衡量物種競(jìng)爭(zhēng)力的重要指標(biāo) (蔣利玲等,2017)。因此,該文綜述國(guó)內(nèi)外關(guān)于植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究現(xiàn)狀,同時(shí)歸納和總結(jié)相應(yīng)的機(jī)理,以期為更好地認(rèn)識(shí)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性在生態(tài)系統(tǒng)結(jié)構(gòu)、功能和穩(wěn)定性維持方面的作用,進(jìn)而為國(guó)內(nèi)深入開(kāi)展生態(tài)化學(xué)計(jì)量學(xué)研究提供理論基礎(chǔ)和參考。
1 生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的概念及原理
內(nèi)穩(wěn)性理論即是指生物在面對(duì)外界環(huán)境中元素可利用性變化的時(shí)候保持自身化學(xué)計(jì)量特征的相對(duì)穩(wěn)定 (Sterner & Elser, 2002)。在生物長(zhǎng)期的進(jìn)化過(guò)程中,生物有機(jī)體通過(guò)動(dòng)態(tài)平衡維持其自身化學(xué)組成的相對(duì)穩(wěn)定,使生物有機(jī)體內(nèi)部環(huán)境的變化保持在較小的范圍,生物有機(jī)體內(nèi)形成一定的內(nèi)穩(wěn)態(tài)機(jī)制 (Sterner & Elser, 2002; Zhang et al., 2003)。這種由生物在長(zhǎng)期進(jìn)化過(guò)程中為適應(yīng)外界環(huán)境變化而形成的維持自身化學(xué)組成相對(duì)穩(wěn)定的能力就叫作生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性(簡(jiǎn)稱(chēng)內(nèi)穩(wěn)性) (Kooijman, 1995; 蔣利玲等,2017)。有機(jī)體化學(xué)元素組成的動(dòng)態(tài)平衡原理是生態(tài)化學(xué)計(jì)量學(xué)理論成立的理論基礎(chǔ)。生物學(xué)中的動(dòng)態(tài)平衡就要使有機(jī)體內(nèi)部的養(yǎng)分平衡、pH值等不隨外部環(huán)境的變化而劇烈變化,從而使整個(gè)有機(jī)體基本保持穩(wěn)定,故動(dòng)態(tài)平衡被認(rèn)為是生命的本質(zhì)特征 (曾德慧和陳廣生,2005)。在生態(tài)化學(xué)計(jì)量學(xué)中,有機(jī)體元素的動(dòng)態(tài)平衡是指有機(jī)體中元素組成與它們周?chē)h(huán)境、可利用的資源和養(yǎng)分元素供應(yīng)保持相對(duì)穩(wěn)定的一種狀態(tài),動(dòng)態(tài)平衡是生態(tài)化學(xué)計(jì)量學(xué)存在的理論基礎(chǔ) (Sterner & Elser, 2002)。
植物的生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征是植物在長(zhǎng)期的進(jìn)化過(guò)程中對(duì)環(huán)境適應(yīng)的結(jié)果,面對(duì)環(huán)境變化時(shí),植物通過(guò)生化、生理與生態(tài)過(guò)程來(lái)維持其自身元素特征的穩(wěn)定 (Sterner & Elser, 2002; 蔣利玲等,2014)。當(dāng)某種元素限制植物的生長(zhǎng)時(shí),植物可通過(guò)多種生理生化機(jī)制改變?cè)撛氐目衫眯砸约袄眯剩瑥亩S持機(jī)體的養(yǎng)分含量以及相關(guān)性狀的穩(wěn)定 (Hessen et al., 2004)。元素的吸收不僅受植物體內(nèi)此種元素的貯存量調(diào)控,也受其它元素含量的影響,當(dāng)細(xì)胞中C、N含量較高或P含量較低時(shí)植物均會(huì)減少對(duì)NO-3的吸收 (Chapin, 1991;Grossman & Takahashi, 2001; Frost et al., 2005)。C、N、P是主要的生命元素,也是生物體組成的重要基礎(chǔ),因此,生態(tài)化學(xué)計(jì)量學(xué)中的內(nèi)穩(wěn)性特征主要是指C、N、P等元素含量以及比例關(guān)系的穩(wěn)定 (Tang & Dam, 1999; Sterner & Elser, 2002; 蘇強(qiáng),2012)。
生物有機(jī)體由多種不同的化合物組成,每種化合物有其特有的元素組成和比例。此外,有機(jī)體內(nèi)也含有大量自由離子,植物以離子形式吸收營(yíng)養(yǎng)元素,因此,有機(jī)體的元素組成及比例關(guān)系受環(huán)境中元素組成的影響。如果生物有機(jī)體中元素含量及比例關(guān)系的變化與外界環(huán)境中的元素含量及比例變化完全一致,則有機(jī)體為非穩(wěn)態(tài)。此時(shí)有機(jī)體與外界環(huán)境中的化學(xué)計(jì)量特征為正相關(guān)關(guān)系,斜率為1 (圖1:A)。如果有機(jī)體的化學(xué)計(jì)量特征與環(huán)境資源的化學(xué)計(jì)量特征無(wú)關(guān),則兩者的化學(xué)計(jì)量關(guān)系曲線(xiàn)的斜率為0 (圖1:B),有機(jī)體為絕對(duì)的內(nèi)穩(wěn)態(tài)。生物有機(jī)體元素含量的變化具有一定范圍,絕對(duì)非穩(wěn)態(tài)與絕對(duì)穩(wěn)態(tài)的生物均不存在 (蘇強(qiáng),2012)。
生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性是一個(gè)可以定量計(jì)算的參數(shù),Sterner & Elser (2002) 提出了生物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性模型,將有機(jī)體的化學(xué)計(jì)量特征與環(huán)境化學(xué)計(jì)量特征的關(guān)系表示為dyy=1H dxx。
式中,x指環(huán)境中營(yíng)養(yǎng)物質(zhì)的供應(yīng)量,y指有機(jī)體中的元素含量,x和y為濃度百分比或元素含量的配比,如P%、N%或N∶P等。將該公式以指數(shù)形式表示,可得y=cx1H。
式中,c為常數(shù)。對(duì)公式兩邊進(jìn)行對(duì)數(shù)轉(zhuǎn)換,x和y的關(guān)系則可以表示為
lny=lnc+1Hlnx,內(nèi)穩(wěn)性指數(shù)H可以表示為H=lnxlny-lnc。
在此模型中,H>1可視為具有維持內(nèi)穩(wěn)態(tài)的能力 (Sterner & Elser, 2002)。為方便統(tǒng)計(jì),多用1/H(0<1/H<1)來(lái)衡量?jī)?nèi)穩(wěn)性的強(qiáng)弱 (Hood & Sterner, 2010)。Persson et al. (2010)將1/H劃分為四個(gè)類(lèi)型:0<1/H<0.25,穩(wěn)態(tài)型;0.25<1/H<0.5,弱穩(wěn)態(tài)型;0.5<1/H<0.75,弱敏感型;1/H>0.75,敏感型。但在維管植物內(nèi)穩(wěn)性的研究中,一些學(xué)者直接測(cè)算H值來(lái)表征內(nèi)穩(wěn)性的大小 (Yu et al., 2010, 2011, 2015; Li et al., 2016)。Yu et al. (2010)認(rèn)為,生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)H可預(yù)測(cè)有機(jī)體的多種內(nèi)在特性,如有機(jī)體對(duì)非生物因素的生理調(diào)節(jié)能力。
盡管化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)對(duì)預(yù)測(cè)物種在種群動(dòng)態(tài)、食物網(wǎng)和養(yǎng)分循環(huán)中的角色有著重要作用,但無(wú)論在生理上還是統(tǒng)計(jì)上,對(duì)參數(shù)1/H的運(yùn)用與解讀都需要謹(jǐn)慎,因?yàn)?/H和消費(fèi)者化學(xué)計(jì)量?jī)?nèi)穩(wěn)性調(diào)節(jié)程度之間不是線(xiàn)性關(guān)系而是指數(shù)關(guān)系,將1/H分為幾個(gè)類(lèi)別來(lái)衡量化學(xué)計(jì)量?jī)?nèi)穩(wěn)性可能過(guò)于簡(jiǎn)單 (Persson et al., 2010)。當(dāng)環(huán)境中化學(xué)計(jì)量特征變化很大,而消費(fèi)者化學(xué)計(jì)量受到強(qiáng)烈限制以及消費(fèi)者化學(xué)計(jì)量獨(dú)立于環(huán)境而變化很大時(shí),內(nèi)穩(wěn)性指數(shù)不能準(zhǔn)確地反映消費(fèi)者的內(nèi)穩(wěn)性。此外,通過(guò)野外試驗(yàn)研究,測(cè)算出有機(jī)體的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)為負(fù)值 (Persson et al., 2010; Xing et al., 2015)。這些一般都以?xún)?nèi)穩(wěn)性指數(shù)H的絕對(duì)值來(lái)表征生物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性之強(qiáng)弱。
2 植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
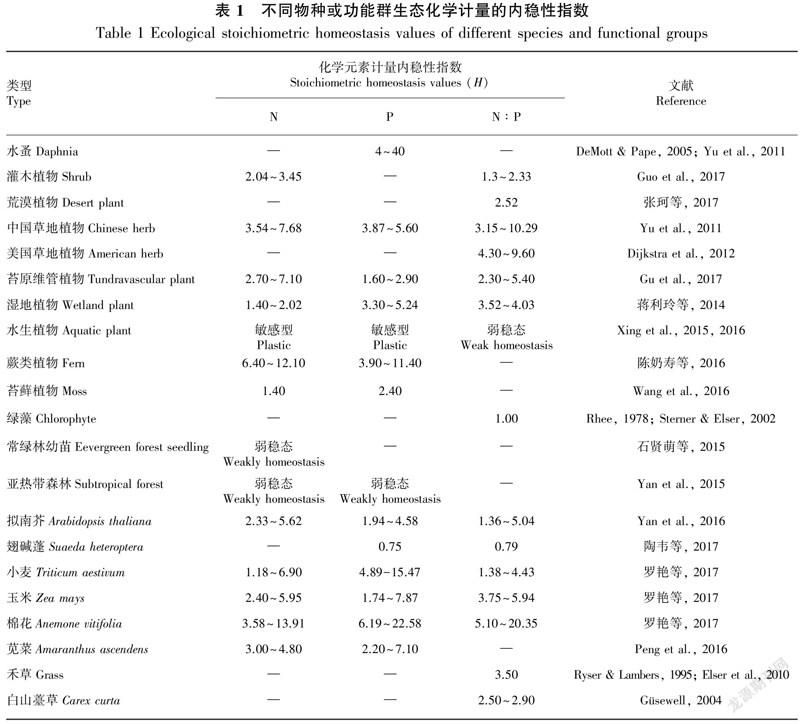
穩(wěn)態(tài)性是生物有機(jī)體的本質(zhì)特征,包括pH值、水分等參數(shù)的穩(wěn)定性,生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性主要指元素組成及比例的穩(wěn)態(tài)性特征。植物元素組成的穩(wěn)定性主要受其基本的生理過(guò)程調(diào)節(jié),如養(yǎng)分的吸收、同化、利用等 (Sterner & Elser, 2002)。由于不同生長(zhǎng)階段對(duì)養(yǎng)分的需求不同,植物的內(nèi)穩(wěn)性指數(shù)隨著生長(zhǎng)階段變化而具有較大的變化。內(nèi)穩(wěn)性指數(shù)不僅可以反應(yīng)植物的穩(wěn)定性特征以及對(duì)環(huán)境的適應(yīng)策略,同時(shí)優(yōu)勢(shì)種的內(nèi)穩(wěn)性強(qiáng)度也是反映生態(tài)系統(tǒng)穩(wěn)定性以及生產(chǎn)力的重要參數(shù) (Yu et al., 2010)。內(nèi)穩(wěn)性較強(qiáng)的植物的養(yǎng)分利用方式較為保守,在貧瘠的環(huán)境中也能維持機(jī)體的緩慢生長(zhǎng);而穩(wěn)定性較弱的植物的適應(yīng)性更強(qiáng) (Persson et al., 2010)。因此,內(nèi)穩(wěn)性強(qiáng)的植物可能更適應(yīng)于穩(wěn)定的環(huán)境,而內(nèi)穩(wěn)性指數(shù)較低的植物在多變的環(huán)境中更有優(yōu)勢(shì)。化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)(H)是生態(tài)化學(xué)計(jì)量學(xué)的重要參數(shù)之一,但關(guān)于植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)態(tài)指數(shù)的定量數(shù)據(jù)仍不多 (曾冬萍等,2013)。從目前有限的文獻(xiàn)來(lái)分析,生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)的大小或強(qiáng)弱在不同植物種類(lèi)或不同植物功能群、不同生長(zhǎng)階段、不同器官或組織中存在較大差異,因此我們從以下方面對(duì)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征進(jìn)行綜述。
2.1 不同物種和功能群的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
不同物種或功能群落對(duì)化學(xué)元素的吸收、運(yùn)輸、分配、利用和釋放過(guò)程存在顯著差異 (嚴(yán)正兵等,2013;Sistla et al., 2015)。不同植物種類(lèi)或功能群對(duì)環(huán)境的生態(tài)適應(yīng)策略有所不同 (Güsewell, 2004),他們通過(guò)生理生態(tài)過(guò)程調(diào)整其自身的化學(xué)元素的含量和比例關(guān)系來(lái)適應(yīng)環(huán)境,因此,不同種類(lèi)或功能群的植物可能具有不同的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征 (表1)。Persson et al. (2010)的研究發(fā)現(xiàn),有些物種的化學(xué)計(jì)量表現(xiàn)為嚴(yán)格的內(nèi)穩(wěn)態(tài),而有些物種的穩(wěn)態(tài)性較弱。Yu et al. (2010)的研究結(jié)果表明,植物的內(nèi)穩(wěn)性指數(shù)與其優(yōu)勢(shì)度高低呈正相關(guān)關(guān)系,即優(yōu)勢(shì)種的內(nèi)穩(wěn)性指數(shù)高于其他物種,認(rèn)為內(nèi)穩(wěn)性指數(shù)高的物種對(duì)養(yǎng)分的利用方式可能更加保守,從而更容易適應(yīng)草原生態(tài)系統(tǒng)貧瘠的環(huán)境而成為優(yōu)勢(shì)種。在P添加條件下,亞高寒草甸禾本科植物生物量和P的內(nèi)穩(wěn)性指數(shù)均高于豆科、莎草科以及雜類(lèi)草的其他物種,這可能與禾本科植物具有發(fā)達(dá)的根系,能夠較快的吸收添加的養(yǎng)分有關(guān),而且較高的P內(nèi)穩(wěn)性對(duì)禾草科的競(jìng)爭(zhēng)力也有很大影響 (張仁懿等,2015)。在水域生態(tài)系統(tǒng)中,大型植物的元素變化范圍比藻類(lèi)植物小,即大型植物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性高于藻類(lèi)植物 (Cross et al., 2005; Demars & Edwards 2007; Tsoi et al., 2011; Feijoó et al., 2014)。灌木和苔蘚植物在施肥試驗(yàn)中表現(xiàn)出不同的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征,苔蘚植物N元素的內(nèi)穩(wěn)性較灌木弱,而P元素的內(nèi)穩(wěn)性較灌木植物強(qiáng) (Wang et al., 2016)。羅艷 (2017)的研究發(fā)現(xiàn),小麥、玉米、棉花的內(nèi)穩(wěn)性指數(shù)隨著其植株生長(zhǎng)發(fā)育的變化趨勢(shì)存在顯著差異。豆科灌木對(duì)環(huán)境中N元素的依賴(lài)性低,豆科灌木中N含量和N∶P高于非豆科灌木,與豆科灌木相比,非豆科灌木中N含量與土壤N含量的相關(guān)性更強(qiáng),表明豆科灌木比非豆科灌木具有更高的N內(nèi)穩(wěn)性 (Guo et al., 2017)。入侵植物內(nèi)穩(wěn)性高于本土植物,其較高的內(nèi)穩(wěn)性可能是入侵成功的一個(gè)原因 (蔣利玲等,2014,2017)。
2.2 植物不同生長(zhǎng)階段的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
植物的生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性隨著植物的生長(zhǎng)發(fā)育進(jìn)程而發(fā)生一定的變化,不同物種的內(nèi)穩(wěn)性可能由于其生長(zhǎng)策略不同而具有不同的變化趨勢(shì);而且由于植物不同器官行使的功能不同,同一物種不同器官的內(nèi)穩(wěn)性變化趨勢(shì)也存在一定差異。Yu et al. (2011)的研究表明,葉片中N、P元素的內(nèi)穩(wěn)性指數(shù)HN、HP和N∶P比的內(nèi)穩(wěn)性指數(shù)HN∶P均隨著生長(zhǎng)季節(jié)的延長(zhǎng)而增大,即葉片的內(nèi)穩(wěn)性指數(shù)與生長(zhǎng)階段呈正比例關(guān)系,由此說(shuō)明隨著植物調(diào)控系統(tǒng)的逐漸發(fā)育成熟,他們對(duì)環(huán)境中養(yǎng)分含量變化的響應(yīng)逐漸變?nèi)酢S捎诓煌卦谥参飪?nèi)部的作用不同,N、P等不同元素隨著植物的生長(zhǎng)發(fā)育呈現(xiàn)不同的變化趨勢(shì) (Peng et al., 2016)。在濕地生態(tài)系統(tǒng)中,不同物種器官的內(nèi)穩(wěn)性指數(shù)在不同生長(zhǎng)發(fā)育階段有所不同, 隨著植物生長(zhǎng)發(fā)育,互花米草根的HN、HP、HN∶P、蘆葦?shù)母颓o的HP、HN∶P和短葉茳芏根的HN∶P下降,而互米草莖和葉的HP、HN∶P、蘆葦葉的HP、HN∶P和短葉茳芏莖的HN∶P則呈現(xiàn)出增加趨勢(shì),這反映了在養(yǎng)分受限的環(huán)境中植物對(duì)其生長(zhǎng)過(guò)程中地上與地下器官養(yǎng)分分配的權(quán)衡,也是其對(duì)環(huán)境長(zhǎng)期適應(yīng)的結(jié)果 (蔣利玲等,2014)。Peng et al. (2016)通過(guò)對(duì)莧菜不同生長(zhǎng)階段的N、P元素的內(nèi)穩(wěn)性指數(shù)的研究發(fā)現(xiàn),莧菜幼苗期、開(kāi)花期、種子灌漿期葉片的HN分別為4.76、3.03、4.35,Hp在幼苗期為2.17,在開(kāi)花期達(dá)到最大,為7.14,他們認(rèn)為N元素調(diào)控植物器官的形成,莧菜生長(zhǎng)階段N的內(nèi)穩(wěn)性指數(shù)高于繁殖階段,而在繁殖階段P元素的HP較高是由于植物需要穩(wěn)定的P供給來(lái)維持其生殖生長(zhǎng)。
2.3 植物不同器官的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
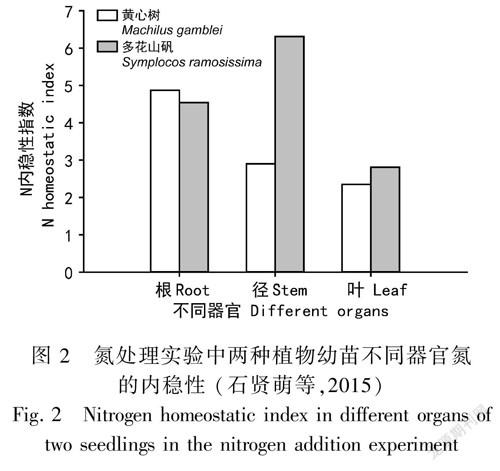
由于植物的不同器官所行使的功能不同,同種生物的不同器官也具有不同的內(nèi)穩(wěn)性特征。在內(nèi)蒙古草原,維管植物地下部分的內(nèi)穩(wěn)性指數(shù)與地上部分呈負(fù)相關(guān)關(guān)系,這反映了植物通過(guò)調(diào)整不同器官養(yǎng)分的分配量以不同的生長(zhǎng)策略適應(yīng)貧瘠的環(huán)境,如優(yōu)勢(shì)種羊草通過(guò)內(nèi)穩(wěn)性較低的地下部分對(duì)養(yǎng)分的吸收、貯存來(lái)維持地上部分較高的內(nèi)穩(wěn)性,從而使植物能夠穩(wěn)定生長(zhǎng) (Yu et al., 2011)。在東北退化草原,羊草根的HN、HP均高于葉 (Li et al., 2016)。在濕地生態(tài)系統(tǒng)中,互花米草、短葉茳芏根系的HN均顯著高于莖、鞘的HN∶P顯著高于根 (蔣利玲等,2017)。對(duì)喬木幼苗和灌木植物的研究也發(fā)現(xiàn),植物葉片的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性高于根 (Garrish et al., 2010; Minden & Kleyer, 2014; Schreeg et al., 2014)。根據(jù)我們對(duì)云南哀牢山中山濕性常綠闊葉林中多花山礬、黃心樹(shù)兩種優(yōu)勢(shì)幼苗的化學(xué)計(jì)量特征及其對(duì)N沉降的研究結(jié)果表明,兩種幼苗葉片的N內(nèi)穩(wěn)性均低于莖和根系,而且多花山礬幼苗莖和葉的N內(nèi)穩(wěn)性均高于黃心樹(shù),而其根的N內(nèi)穩(wěn)性低于黃心樹(shù) (圖2),表明不同物種不同器官的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性存在較大差異,其中葉片對(duì)N的輸入要比根系和莖更為敏感(石賢萌等,2015)。從上述研究結(jié)果可看出,植物的地上和地下部分化學(xué)元素內(nèi)穩(wěn)性的變化趨勢(shì)相反,植物通過(guò)調(diào)整不同器官的養(yǎng)分分配和利用方式來(lái)適應(yīng)環(huán)境,以滿(mǎn)足自身生長(zhǎng)發(fā)育的需要。
2.4 不同元素的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征
C、N、P等是植物生長(zhǎng)發(fā)育必需的大量元素,不同元素在植物體內(nèi)發(fā)揮不同的作用,導(dǎo)致不同元素的含量以及內(nèi)穩(wěn)性特征具有一定差異。浮游生物體內(nèi)含量較高的元素的內(nèi)穩(wěn)性要高于含量少的元素,如N的內(nèi)穩(wěn)性高于P;微量元素的內(nèi)穩(wěn)性低于主要元素 (大量元素) (Karimi & Folt, 2006; Han et al., 2011)。目前,對(duì)不同類(lèi)群植物內(nèi)穩(wěn)性的研究結(jié)果大多支持這一觀點(diǎn)。如內(nèi)蒙古草原維管植物地上部分和地下部分的HN均高于HP,表明植物對(duì)其組織內(nèi)部含量更高的化學(xué)元素N有更強(qiáng)的調(diào)控能力 (Yu et al., 2011);長(zhǎng)芒草中C元素含量的變異系數(shù)較小,為3.65%,表現(xiàn)出較強(qiáng)的內(nèi)穩(wěn)性,而N、P元素含量的變異系數(shù)均大于30%,內(nèi)穩(wěn)性較弱 (牛得草等,2011);不同發(fā)育階段烤煙中C元素含量的變異系數(shù)最小,內(nèi)穩(wěn)性最高 (楊梅等,2015);東北退化草原優(yōu)勢(shì)物種羊草葉片的HN大于HP(Li et al., 2016);在濕地植物中,N元素的內(nèi)穩(wěn)性指數(shù)高于P元素 (蔣利玲等,2017)。
此外,植物N、P元素的內(nèi)穩(wěn)性指數(shù) (HN、HP)小于HN∶P,這是因?yàn)橹参锝M織內(nèi)部N和P的變化具有協(xié)同性,即植物組織內(nèi)N元素的累積通常伴隨著P含量的升高 (Sterner & Elser, 2002)。相較于HN和HP,HN∶P受外界環(huán)境等因素的影響較小,與植物本身的相關(guān)性更強(qiáng),表明植物對(duì)其自身內(nèi)穩(wěn)性的調(diào)節(jié)主要體現(xiàn)在NP值而不是每種元素的含量上 (趙君等,2011)。因此,在判斷某種植物內(nèi)穩(wěn)性強(qiáng)弱時(shí),用HN∶P表示植物的內(nèi)穩(wěn)性更為接近植物維持其自身內(nèi)穩(wěn)態(tài)的真實(shí)能力。
3 植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性與生態(tài)系統(tǒng)結(jié)構(gòu)、功能和穩(wěn)定性的關(guān)系
生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性不僅是用來(lái)檢驗(yàn)生物與其外界環(huán)境的關(guān)系的工具,它也可作為判斷物種豐富度與生態(tài)系統(tǒng)結(jié)構(gòu)、功能與穩(wěn)定性的重要指標(biāo) (Sterner & Elser, 2002; Yu et al., 2010)。植物群落中優(yōu)勢(shì)種的內(nèi)穩(wěn)性較其他物種高,內(nèi)穩(wěn)性指數(shù)的大小與植物的適應(yīng)策略以及對(duì)環(huán)境適應(yīng)性的強(qiáng)弱有關(guān)。植物的養(yǎng)分利用策略是調(diào)控生物多樣性與生態(tài)系統(tǒng)結(jié)構(gòu)、功能以及穩(wěn)定性的關(guān)鍵因素,優(yōu)勢(shì)種內(nèi)穩(wěn)性高的生態(tài)系統(tǒng)的生產(chǎn)力更高,表明植物的生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性可能是維持生態(tài)系統(tǒng)的結(jié)構(gòu)以及穩(wěn)定性的重要機(jī)制 (Yu et al., 2010)。在草原生態(tài)系統(tǒng)中,N∶P比值高的物種具有較高的內(nèi)穩(wěn)性指數(shù),而內(nèi)穩(wěn)性高的物種有更高和更穩(wěn)定的生物量,由內(nèi)穩(wěn)性高的物種占優(yōu)勢(shì)的生態(tài)系統(tǒng)有更高的生產(chǎn)力和穩(wěn)定性,化學(xué)計(jì)量?jī)?nèi)穩(wěn)性可能是草原生態(tài)系統(tǒng)結(jié)構(gòu)、功能和穩(wěn)定性維持的重要機(jī)理 (Yu et al., 2010, 2011)。
Yu et al. (2010)的研究結(jié)果表明,在物種水平上,短期、長(zhǎng)期和空間梯度試驗(yàn)中植物葉片的HN均與其物種的豐富度和內(nèi)穩(wěn)性呈嚴(yán)密的正相關(guān)關(guān)系,而HP則與物種豐富度、穩(wěn)定性相關(guān)性較差,表明草原生態(tài)系統(tǒng)主要受N調(diào)控;在群落水平上,兩年的野外施肥試驗(yàn)和27 a的監(jiān)測(cè)試驗(yàn)中群落的H值均與群落的穩(wěn)定性、生產(chǎn)力呈正相關(guān)關(guān)系,而在1 200km的空間梯度試驗(yàn)中這種相關(guān)性關(guān)系僅見(jiàn)于草甸草原,在典型性草原和荒漠草原中群落的內(nèi)穩(wěn)性強(qiáng)弱和群落的生產(chǎn)力、穩(wěn)定性不相關(guān)。Bai et al. (2010)的研究結(jié)果發(fā)現(xiàn),經(jīng)過(guò)4 a的N添加試驗(yàn),生態(tài)系統(tǒng)中內(nèi)穩(wěn)性較高的物種的生物量仍然較高。在美國(guó)C4植物為優(yōu)勢(shì)種的草原生態(tài)系統(tǒng)中,經(jīng)過(guò)9 a的N添加,HN較高的物種的多度降低,HN較低的物種的多度增高,HN可以準(zhǔn)確的預(yù)測(cè)不同物種對(duì)N添加的響應(yīng)趨勢(shì);而在一系列模擬水分變化的氣候變化實(shí)驗(yàn)結(jié)果表明,N內(nèi)穩(wěn)性最高的物種穩(wěn)定性較強(qiáng),對(duì)土壤水分供應(yīng)變化的響應(yīng)最小,N內(nèi)穩(wěn)性較低的物種則對(duì)水分變化更敏感,HN和物種的優(yōu)勢(shì)度之間的正相關(guān)關(guān)系未受水分變化的影響 (Yu et al., 2015)。相對(duì)而言,化學(xué)計(jì)量?jī)?nèi)穩(wěn)性高的植物物種對(duì)水分變化的敏感性較低。我們前期對(duì)云南哀牢山地區(qū)森林附生苔蘚的研究發(fā)現(xiàn):附生苔蘚對(duì)N沉降、空氣濕度變化非常敏感,這主要與附生苔蘚無(wú)根系、不從土壤吸收養(yǎng)分,其生命過(guò)程所需養(yǎng)分和水分來(lái)自于大氣,因此附生苔蘚可作為檢測(cè)環(huán)境變化的指示生物 (Song et al., 2012a, 2012b)。此外,我們對(duì)兼性附生植物大果假瘤蕨的研究表明,附生與地生植株的形態(tài)和生理性狀均發(fā)生了顯著的可塑性變化,附生植株功能性狀的可塑性變化有利于緩解水分虧缺的負(fù)面作用,而地生植株的性狀則表現(xiàn)為降低林下低光脅迫的消極影響(Lu et al., 2015)。因此我們推測(cè),附生植物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性可能較低、群落結(jié)構(gòu)穩(wěn)定性可能較弱。目前我們正在開(kāi)展相關(guān)的實(shí)驗(yàn)研究,深入探討在不受土壤基質(zhì)養(yǎng)分影響下森林附生植物的生態(tài)化學(xué)計(jì)量學(xué)特征。
內(nèi)穩(wěn)性高的物種具有更高的穩(wěn)定性和優(yōu)勢(shì)度,且可能提高生態(tài)系統(tǒng)的抵抗力穩(wěn)定性,而內(nèi)穩(wěn)性低的物種會(huì)影響生態(tài)系統(tǒng)保持其穩(wěn)定性的能力。Li et al. (2016)的研究表明,草原優(yōu)勢(shì)物種的內(nèi)穩(wěn)性較高,在維持草原生態(tài)系統(tǒng)穩(wěn)定性方面具有重要作用,而且退化草原土壤的C、N、P含量明顯降低。在草原生態(tài)系統(tǒng)中,HN高的物種比HN低的物種具有更高的穩(wěn)定性和多度,HN可能可以預(yù)測(cè)物種多度和穩(wěn)定性,以及植物物種和生態(tài)系統(tǒng)將如何響應(yīng)全球變化引起的資源可利用性的改變 (Yu et al., 2015),但這種預(yù)測(cè)是否能擴(kuò)展到其他生活型或生態(tài)系統(tǒng)如沙漠、森林、濕地生態(tài)系統(tǒng)等,仍需做進(jìn)一步的研究探索。
4 植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的影響因素
由于內(nèi)穩(wěn)性是植物在進(jìn)化過(guò)程中對(duì)環(huán)境的長(zhǎng)期適應(yīng)而形成的 (Elser et al., 2010),目前對(duì)植物內(nèi)穩(wěn)性影響因素的研究主要集中在植物的內(nèi)在因素,而關(guān)于全球氣候變化 (如氣候變暖、CO2濃度升高、N、P沉降等),人為干擾 (如施肥、放牧、土地利用方式變化、火災(zāi)等)等外界因素對(duì)植物內(nèi)穩(wěn)性的影響的研究較為少見(jiàn)。全球變化以及人為活動(dòng)干擾通過(guò)改變土壤中N、P元素的含量而導(dǎo)致物種的優(yōu)勢(shì)度改變,從而影響群落以及生態(tài)系統(tǒng)的結(jié)構(gòu)和生產(chǎn)力 (Güsewell, 2004)。外界環(huán)境因素的變化是否會(huì)引起植物內(nèi)穩(wěn)性的改變尚無(wú)相關(guān)的報(bào)道,還需加強(qiáng)此方面的研究。
植物群落中養(yǎng)分元素的含量隨不同演替階段而發(fā)生變化。Yan et al. (2015)通過(guò)對(duì)亞熱帶森林演替過(guò)程中先鋒種馬尾松的研究發(fā)現(xiàn),在演替初期,土壤中的N元素含量較低,HN較高的馬尾松更能適應(yīng)環(huán)境而成為優(yōu)勢(shì)種;隨著森林生態(tài)系統(tǒng)的演替,土壤中的N含量增高,喜N的闊葉物種將大量的N、P吸收并積累在其體內(nèi),造成土壤中P元素尤其是有效P的含量明顯減少,P內(nèi)穩(wěn)性較低的馬尾松的優(yōu)勢(shì)度降低。Yu et al. (2011)的研究結(jié)果表明,盡管2006年和2007年的降雨量具有明顯差異 (304、240 mm),但羊草、糙隱子草、灰綠藜3種植物的內(nèi)穩(wěn)性指數(shù)(HN,HP和HN∶P)在這2 a內(nèi)的差異均未達(dá)到顯著水平。Yu et al. (2015)對(duì)美國(guó)草原生態(tài)系統(tǒng)的研究發(fā)現(xiàn),在N添加和改變水分可利用量的條件下,物種間的穩(wěn)定性關(guān)系并未改變。因此,我們推測(cè)植物的內(nèi)穩(wěn)性是植物的基本屬性,尤其是對(duì)內(nèi)穩(wěn)性高的物種而言,受降水、N沉降等外界環(huán)境因素變化的影響可能較小。
5 研究展望
以“化學(xué)計(jì)量?jī)?nèi)穩(wěn)性”為關(guān)鍵詞在中國(guó)知網(wǎng)、Web of Science等網(wǎng)站搜索相關(guān)的文獻(xiàn)或資料,并對(duì)其進(jìn)行學(xué)術(shù)關(guān)注度分析比較,結(jié)果表明國(guó)際上對(duì)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究起步較早,但主要集中在動(dòng)物研究,對(duì)植物的研究較少,植物內(nèi)穩(wěn)性相關(guān)文章較為少見(jiàn);雖然我國(guó)植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的關(guān)注度自2014年呈緩慢上升的趨勢(shì),但到現(xiàn)在為止所發(fā)表的相關(guān)論文數(shù)量仍不多。目前,我國(guó)對(duì)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究的關(guān)注度不是很高,研究成果也較少。
植物的內(nèi)穩(wěn)性與其對(duì)環(huán)境的適應(yīng)策略以及群落組成、結(jié)構(gòu)和生態(tài)系統(tǒng)的穩(wěn)定性以及生產(chǎn)力息息相關(guān) (Yu et al., 2010)。植物是構(gòu)成陸地生態(tài)系統(tǒng)的主體,為生態(tài)系統(tǒng)中各級(jí)消費(fèi)者直接或間接地提供能量,是生態(tài)系統(tǒng)物質(zhì)循環(huán)和能量流動(dòng)的重要環(huán)節(jié)。由人類(lèi)活動(dòng)干擾導(dǎo)致的全球氣候、環(huán)境變化、N沉降等顯著影響植物的生長(zhǎng)發(fā)育、物種組成、物種多樣性與分布、生產(chǎn)力,引起植物群落組成與結(jié)構(gòu)的改變,從而導(dǎo)致生態(tài)系統(tǒng)的變動(dòng)加劇 (Elser et al., 2010)。生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性從養(yǎng)分元素的角度研究植物的穩(wěn)定性,以及他們與群落結(jié)構(gòu)、生態(tài)系統(tǒng)功能與穩(wěn)定性和生產(chǎn)力的關(guān)系,以及植物的內(nèi)穩(wěn)性特征對(duì)N沉降的響應(yīng),為生態(tài)學(xué)研究提供了新的切入點(diǎn)。因此,加強(qiáng)對(duì)植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究至關(guān)重要。基于我國(guó)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究現(xiàn)狀,在未來(lái)的研究中建議對(duì)以下幾個(gè)方面進(jìn)行深入的研究探討。
(1)植物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征與生態(tài)系統(tǒng)結(jié)構(gòu)的穩(wěn)定性以及生態(tài)系統(tǒng)生產(chǎn)力的關(guān)系。植物的C、N、P元素含量以及N∶P的生態(tài)化學(xué)計(jì)量特征和環(huán)境中元素含量對(duì)植物生長(zhǎng)發(fā)育的影響已被廣泛研究 (Wright et al., 2004; Han et al., 2005)。但是,目前對(duì)植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的研究還較少,且已有的研究還停留在物種水平上,對(duì)植物內(nèi)穩(wěn)性與群落、生態(tài)系統(tǒng)的關(guān)系則研究更為少見(jiàn)。有學(xué)者認(rèn)為植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的強(qiáng)弱與植物的優(yōu)勢(shì)度正相關(guān),植物的內(nèi)穩(wěn)性可能會(huì)影響生態(tài)系統(tǒng)的結(jié)構(gòu)以及生產(chǎn)力 (Yu et al., 2010)。Dijkstra et al. (2012)對(duì)美國(guó)半干旱草原三個(gè)優(yōu)勢(shì)種的研究發(fā)現(xiàn),其中一個(gè)優(yōu)勢(shì)種的N∶P變異幅度很大,內(nèi)穩(wěn)性較弱,這與Yu et al. (2010)對(duì)內(nèi)蒙古草原的研究結(jié)果相反。目前,國(guó)內(nèi)關(guān)于植物的內(nèi)穩(wěn)性對(duì)植物的生長(zhǎng)發(fā)育以及生態(tài)系統(tǒng)結(jié)構(gòu)影響的研究主要集中于內(nèi)蒙古草原生態(tài)系統(tǒng),而對(duì)其它生態(tài)系統(tǒng)的研究鮮有報(bào)道。因此,應(yīng)加強(qiáng)對(duì)不同類(lèi)型生態(tài)系統(tǒng)和在更大地理尺度上來(lái)研究探討植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征及其變化規(guī)律,來(lái)驗(yàn)證植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性理論是否具有普適性。
(2)植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的生理生態(tài)學(xué)意義。植物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性與植物的適應(yīng)性以及生態(tài)策略密切相關(guān) (Güsewell, 2005; Frost et al., 2005; Yu et al., 2010)。內(nèi)穩(wěn)性較強(qiáng)的植物的養(yǎng)分利用方式較為保守,在貧瘠的環(huán)境中也能維持機(jī)體的緩慢生長(zhǎng),但內(nèi)穩(wěn)性指數(shù)較低的植物的適應(yīng)性卻更強(qiáng)。內(nèi)穩(wěn)性強(qiáng)的植物適應(yīng)于穩(wěn)定的環(huán)境,而內(nèi)穩(wěn)性指數(shù)較低的植物在多變的環(huán)境中卻更有優(yōu)勢(shì) (Persson et al., 2010)。但是,有關(guān)植物的內(nèi)穩(wěn)性強(qiáng)弱及其在環(huán)境變化的響應(yīng)機(jī)制與生態(tài)適應(yīng)策略卻還缺乏深入研究。因此,需要進(jìn)一步加強(qiáng)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性的生理生態(tài)學(xué)研究。
(3)植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性與生長(zhǎng)速率的關(guān)系。許多學(xué)者研究發(fā)現(xiàn),在相對(duì)受限的生化分配條件下,藻類(lèi)為了滿(mǎn)足自身快速生長(zhǎng)可能會(huì)將其N(xiāo)∶P限制在一個(gè)狹窄的范圍內(nèi),藻類(lèi)的生態(tài)化學(xué)計(jì)量的內(nèi)穩(wěn)性可能與生長(zhǎng)速率具有一定的內(nèi)在關(guān)聯(lián) (Elrifi & Turpin 1985;Shafik et al., 1997; Persson et al., 2010)。目前對(duì)維管植物的研究發(fā)現(xiàn),在物種水平上,生長(zhǎng)速率變化較大的物種,其N(xiāo)∶C,P∶C和 N∶P比的變化范圍較大;而生長(zhǎng)速率變化較小的物種,其C∶N∶P比的變化范圍也較小,兩者之間可能存在正相關(guān)的關(guān)系 (Yu et al., 2010, 2012)。然而,目前我國(guó)對(duì)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性高低與生長(zhǎng)速率的關(guān)系的研究卻較少,研究成果匱乏。因此,在未來(lái)的研究中,需加強(qiáng)植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性和生長(zhǎng)速率之間關(guān)系的研究。
(4)特殊生物類(lèi)群如森林附生植物的化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征。目前,關(guān)于生態(tài)化學(xué)內(nèi)穩(wěn)性研究主要集中于海洋生物、陸地植物和部分濕地植物方面。林冠附生植物是山地森林生態(tài)系統(tǒng)中一類(lèi)重要而特殊的植物類(lèi)群,相對(duì)于陸生植物而言,附生植物沒(méi)有與土壤直接相連的根系,不從土壤中吸收無(wú)機(jī)養(yǎng)分,其生命過(guò)程中所需的養(yǎng)分、水分主要來(lái)源于大氣。以往基于陸生植物獲得的有關(guān)生態(tài)化學(xué)計(jì)量學(xué)特征研究的相關(guān)結(jié)果和理論,是否適用于附生植物?在附生植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性指數(shù)(H)的測(cè)算中,其生長(zhǎng)基質(zhì)如何確定?如何評(píng)判其內(nèi)穩(wěn)性強(qiáng)弱,目前尚無(wú)相關(guān)的研究報(bào)道。此外,附生植物的生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征與林冠附生生物亞系統(tǒng),乃至整個(gè)生態(tài)系統(tǒng)的結(jié)構(gòu)、功能和穩(wěn)定性之間的關(guān)系等都是植物生態(tài)化學(xué)計(jì)量學(xué)需要研究探討的新問(wèn)題,在研究對(duì)象、研究方法等方面面臨新的挑戰(zhàn)!值得加強(qiáng)此方面的研究,進(jìn)一步豐富植物生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性研究的理論和方法。
參考文獻(xiàn):
ANDERSEN T, HESSEN DO, 1991 . Carbon, nitrogen, and phosphorus content of freshwater zooplankton [J]. Limnol Oceanogr, 36(4):807-814.
BAI YF, WU JG, CLARK CM, et al., 2010. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands[J]. Globa Change Biol, 16(1): 358-372.
CERNUSAK LA, TURNER WBL, 2009. Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls [J]. New Phytolt, 185(3):770-779.
CHAPIN FS, 1991. Effects of multiple environmental stresses on nutrient availability and use[M] // MOONEY HA, WINNER WE, PELL EJ. Response of plants to multiple stresses. Pittsburgh: Academic Press.
CHEN NS, ZHANG QF, CHEN T, et al., 2016. Stoi-chiometriccharacteristics of nitrogen and phosphorus in Dicranopreris dichotomaduring ecosystem restoration of eroded red-soil[J]. For Res, 29(5):735-742.[陳奶壽, 張秋芳, 陳坦, 等, 2016. 退化紅壤恢復(fù)過(guò)程中芒萁的N、P化學(xué)計(jì)量特征[J]. 林業(yè)科學(xué)研究, 29(5):735-742.]
CROSS, WF, BENSTEAD JP, FROST PC, et al., 2005. Ecological stoichiometry in freshwater benthic systems: Recent progress and perspectives[J]. Freshwater Biol, 50: 1895-1912.
DEMARS BOL, EDWARDS AC, 2007. Tissue nutrient concentrations in freshwater aquatic macrophytes: High inter-taxon differences and low phenotypic response to nutrient supply[J]. Freshwater Biol, 52:2073-2086.
DEMOTT WR, PAPE BJ, 2005. Stoichiometry in an ecological context: Testing for links between Daphnia P-content, growth rate and habitat preference[J]. Oecologia, 142:20-27.
DIJKSTRA FA, PENDALL E, MORGAN JA, et al., 2012. Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland[J]. New Phytol, 196:807-815.
ELRIFI IR, TURPIN DH, 1985. Steady-state luxury consumption and the concept of optimum nutrient ratios: A study with phosphate and nitrate limited Selenastrum minutum(Chlorophyta)[J]. J Phycol, 21:592-602.
ELSER JJ, ANDERSEN T, BARON JS, et al., 2009. Shifts in lake N∶P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition[J]. Science, 326:835-837.
ELSER JJ, FAGAN WF, KERHOFFA J, et al., 2010. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change[J]. New Phytol, 186:593-608.
ELSER JJ, STERNER RW, GOROKHOVA E, et al., 2000. Biological stoichiometry from genes to ecosystems[J]. Ecol Lett, 3(6): 540-550.
FEIJOO C, LEGGIERI L, OCON C, et al., 2014. Stoichiometric homeostasis in the food web of a chronically nutrient-rich stream[J]. Freshwater Sci, 33(3):820-831.
FROST PC, EVANS-WHITE MA, FINKEL ZV, et al., 2005. Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world[J]. Oikos, 109: 18-28.
GARRISH V, CERNUSAK LA, WINTER K, et al., 2010. Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida [J]. J Exp Bot, 61:3735-3748.
GIORDANO M, 2013. Homeostasis: An underestimated focal point of ecology and evolution[J]. Plant Sci, 211(211):92-101.
GOLDMAN JC, CARON DA, DENETT MR,1987. Regulations of gross growth efficiency and ammonium regeneration in bacteria by substrate C∶N ratio[J] . Limnol Oceanogr, 32(6):1239-1252.
GROSSMAN A, TAKAHASHI H, 2001. Micronutrient utilization by photosynthetic eukaryotes and the fabric of interactions[J]. Ann Rev Plant Physiol Plant Mol Biol, 52(52):163-210.
GU Q, ZAMIN TJ, GROGAN P, 2017. Stoichiometric homeostasis: A test to predict tundra vascular plant species and community-level responses to climate change[J]. Arct Sci: 1-14.
GUO Y, YANG X, SCHOB C, et al., 2017. Legume shrubs are more nitrogen-homeostatic than non-legume shrubs[J]. Front Plant Sci, 8:1662.
GSEWELL S, 2005. Responses of wetland graminoids to the relative supply of nitrogen and phosphorus [J]. Plant Ecol, 176:35-55.
GSEWELL S, 2004. N∶P ratios in terrestrial plants: Variation and functional significance[J]. New Phytol, 164(2):243-266.
HAN WX, FANG JY, GUO DL, et al., 2005. Leaf nitrogen and phosphorus stoichio-metryacross 753 terrestrial plant species in China[J]. New Phytol, 168:377-85.
HAN WX, FANG JY, REICH PB, et al., 2011. Biogeography and variability of eleven mineral elementsin plant leaves across gradients of climate, soil and plant functional type in China[J]. Ecol Lett, 14(8):788-796.
HE JS, HAN XG, 2010. Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems[J]. Chin J Plant Ecol, 34(1):2-6.[賀金生, 韓興國(guó), 2010. 生態(tài)化學(xué)計(jì)量學(xué):探索從個(gè)體到生態(tài)系統(tǒng)的統(tǒng)一化理論[J]. 植物生態(tài)學(xué)報(bào), 34(1):2-6.]
HESSON DO, AGREN GI, ANDERSON TR, et al., 2004. Carbon sequestration in ecosystems: The role of stoichiometry[J]. Ecology, 85(5):1179-1192.
HOOD JM, STERNER R, 2010. Diet mixing: Do animals integrate growth or resources across temporal heterogeneity?[J]. Am Nat, 176(5):651-663.
JEYASINGH PD,WEIDER LJ,STERNER RW, 2009. Genetically-based trade-offs in response to stoichiometric food quality influence competition in a keystone aquatic herbivore [J]. Ecol Lett, 12(11): 1229-1237.
JIANG LL, HE S, WU LF, et al., 2014. Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang Estuary[J]. Wetl Sci, 12(3):293-298.[蔣利玲, 何詩(shī), 吳麗鳳, 等, 2014. 閩江河口濕地3種植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征[J]. 濕地科學(xué), 12(3):293-298.]
JIANG LL, ZENG CS, SHAO JJ, et al., 2017. Plant nutrient dynamics and stoichiometric homeostasis of invasive species Spartina alternifloraand native Cyperus malaccensisvar. brevifoliusin the Minjiang River estuarine wetlands[J]. Chin J Plant Ecol, 41(4):450-460.[蔣利玲, 曾從盛, 邵鈞炯, 等, 2017. 閩江河口入侵種互花米草和本地種短葉茳芏的養(yǎng)分動(dòng)態(tài)及植物化學(xué)計(jì)量?jī)?nèi)穩(wěn)性特征[J]. 植物生態(tài)學(xué)報(bào), 41(4):450-460.]
KARIMI R, FOLT CL, 2006. Beyond macronutrients: Element variability and multielement stoichiometry in freshwater invertebrates[J]. Ecol Lett, 9(12):1273-1283.
KOOIJMAN SALM, 1995. The stoichiometry of animal energetics[J]. J Theor Biol, 177, 139-149.
LEVI MP, COWLING EB,1969. Role of nitrogen in wood deterioration. VII. Physiological adaptation of wood-destroying and other fungi to substrates deficient in nitrogen [J] . Phytopathology, 59: 460-468.
LI YF, LI QY, GUO DY, et al., 2016. Ecological stoichio-metryhomeostasis ofLeymus chinensis, in degraded grassland in western Jilin Province, NE China[J]. Ecol Eng, 90:387-391.
LIMPENS J,BERENDSE F, 2003. Growth reduction of Sphagnum magellanicumsubjected to high nitrogen deposition: The role of amino acid nitrogen concentration[J]. Oecologia, 135(3):339-345.
LOLODZE I, KUANG Y, ELSER JJ, 2000. Stoichiometry in producer-grazer systems: Linking energy flow with element cycling[J]. Bull Math Biol, 62:1137-62.
LU HZ, LIU WY, YU FH, et al., 2015. Higher clonal integration in the facultative epiphytic fern Selliguea griffithianagrowing in the forest canopy compared with the forest understorey[J]. Ann Bot, 116(1):113-122.
LUO Y, 2017. Charateristics of ecological stoichiometry of oasis farmland ecosystem in the northern marigin of Tarim Basin[D]. Urumchi:Xinjiang University:1-65.[羅艷, 2017. 塔里木盆地北緣綠洲農(nóng)田生態(tài)系統(tǒng)生態(tài)化學(xué)計(jì)量特征[D]. 烏魯木齊: 新疆大學(xué): 1-65.]
MARSCHNER P, 2012. Marschners mineral nutrition of higher plants[M]. 3rd ed. Academic Press, 48(2) :305.
MATZEK V, VITOUSEK PM, 2009. N∶P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis [J]. Ecol Lett, 12(8):765-771.
MINDEN V, KLEYER M, 2014. Internal and external regulation of plant organ stoichio-metry[J]. Plant Biol, 16: 897-907.
NIU DC, DONG XY, FU H, 2011. Seasonal dynamics of carbon, nitrogen and phosphorus stoichiometry in Stipa bungeana [J]. Pratac Sci, 28(6):915-920.[牛得草, 董曉玉, 傅華, 2011. 長(zhǎng)芒草不同季節(jié)碳氮磷生態(tài)化學(xué)計(jì)量特征[J]. 草業(yè)科學(xué), 28(6):915-920.]
PENG HY, CHEN YH, YAN ZB, et al., 2016. Stage-dependent stoichiometric homeostasis and responses of nutrient resorption in Amaranthus mangostanusto nitrogen and phosphorus addition[J]. Sci Rep, 6:37219.
PERSSON J, FINK P, GOTO A, 2010. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs[J]. Oikos, 119:741-751.
PLNET D, ETCHEBEST S, MOLLIER A, et al., 2000. Growth analysis of maize field crops under phosphorus deficiency. I. Leaf growth[J]. Plant Soil, 223(1-2):119-132.
RHEE GY, 1978. Effects of N∶P atomic ratios and nitrate li-mitationon algal growth, cell composition, and nitrate uptake[J]. Limnol Oce, 23(1):10-25.
RYSER P, LAMERS H, 1995. Root and leaf attributes accoun-tingfor the performance of fast- and slow-growing grasses at different nutrient supply[J]. Plant Soil 170: 251-265.
SCHREEG LA, SANTIAGO LS, WRIGHT SJ, et al., 2014. Stem, root, and older leaf N∶P ratios are more responsive indicators of soil nutrient availability than new foliage[J]. Ecology, 95: 2062-2068.
SHAFIK HM, HERODEK S, PRESING M, et al., 1997. Growth of Cyclotella meneghinianaKutz. II. growth and cell composition under different growth rates with different limiting nutrient[J]. Ann Limnol Int J Limnol, 33:223-233.
SHI XM, QI JH, SONG L, et al., 2015. C, N and P stoichio-metryof two dominant seedlings and their responses to nitrogen additions in the montane moist evergreen broad-leaved forest in Ailao Mountains, Yunnan[J]. Chin J Plant Ecol, 39(10) :962-970.[石賢萌, 杞金華, 宋亮, 等, 2015. 哀牢山中山濕性常綠闊葉林兩種優(yōu)勢(shì)幼苗C、N、P化學(xué)計(jì)量特征及其對(duì)N沉降增加的響應(yīng)[J]. 植物生態(tài)學(xué)報(bào), 39(10):962-970.]
SISTLA SA, APPLING AP, LEWANDOWSKA AM, et al., 2015. Stoichio-metricflexibility in response to fertilization along gradients of environmental and organismal nutrient richness[J]. Oikos, 124(7):949-959.
SONG L, LIU WY, NADKARNI NM, 2012a. Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in Southwest China[J]. Biol Conserv, 152(8):127-135.
SONG L, LIU WY, MA WZ, et al., 2012b. Response of epiphytic bryophytes to simulated N deposition in a subtropical montane cloud forest in southwestern China[J]. Oecologia, 170(3):847-856.
STERNER RW, ELSER JJ, 2002. Ecological stoichiometry: The biology of elements from molecules to the biosphere[M]// Ecological stoichiometry: The biology of elements from molecules to the biosphere: 225-226.
STERNER RW, HESSEN DO, 1994. Algal nutrient limitation and the nutrition of aquatic herbivores[J]. Ann Rev Ecol Evol S, 25:1-29.
SU Q, 2012. The framework of stoichiometry homeostasis in zooplankton elemental composition[J]. Acta Ecol Sin, 32(22): 7213-7219.[蘇強(qiáng), 2012. 浮游動(dòng)物化學(xué)計(jì)量學(xué)穩(wěn)態(tài)性特征研究進(jìn)展[J]. 生態(tài)學(xué)報(bào), 32(22):7213-7219.]
TANG KW, DAM HG, 1999. Limitation of zooplankton production: Beyond stoichiometry[J]. Oikos, 84:537-542.
TAO W, WU JW, LIU CF, et al., 2017. Response of stoichiometric homeostasis and allometric scaling in halophyte Suaeda heteropteraKitag. to simulated nitrogen and phosphorus deposition[J]. J Hydroecol, 38(4) :1674-3075.[陶韋, 武嘉文, 劉長(zhǎng)發(fā), 等, 2017. 翅堿蓬生態(tài)化學(xué)計(jì)量?jī)?nèi)穩(wěn)性對(duì)模擬氮磷沉降的響應(yīng)[J]. 水生態(tài)學(xué)雜志, 38(4):1674-3075.]
TSOI WY, HADWEN WL, FELLOWS CS, 2011. Spatial and temporal variation in the ecological stoichiometry of aquatic organisms in an urban catchment[J]. J N Am Benthol Soc 30:533-545.
WANG M, LARMOLA T, MURPHY MT, et al., 2016. Stoichiometric response of shrubs and mosses to long-term nutrient(N, P and K) addition in an ombrotrophic peatland[J]. Plant Soil, 400(1-2) :1-14.
WANG SQ, YU GR, 2008. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements[J]. Acta Ecol Sin, 28(8):3937-3947.[王紹強(qiáng), 于貴瑞, 2008. 生態(tài)系統(tǒng)碳氮磷元素的生態(tài)化學(xué)計(jì)量學(xué)特征[J]. 生態(tài)學(xué)報(bào), 28(8):3937-3947.]
WILLIAMS RJP,DASILVA JJRF, 1996. The natural selection of the chemical elements: The environment and lifes chemistry[M]. Oxford: Clarendon Press.
WRIGHT IJ, REICH PB, WESTOBY M, et al., 2004. World-wide leaf economics spectrum[J]. Nature, 428:821-827.
XING W, SHI Q, LIU H, et al., 2016. Growth rate, protein: RNA ratio and stoichiometric homeostasis of submerged macrophytes under eutrophication stress[J]. Knowledge Manag Aquat Ecosyst, 7(417).
XING W, WU H, SHI Q, et al., 2015. Multielement stoichiometry of submerged macrophytes across Yunnan plateau lakes(China)[J]. Sci Rep, 5:10186.
YAN JH, LI K, PENG XJ, et al., 2015. The mechanism for exclusion of Pinus massonianaduring the succession in subtropical forest ecosystems: Light competition or stoi-chiometrichomoeostasis?[J]. Sci Rep, 5:10994.
YAN ZB, GUAN HY, HAN WX, et al., 2016. Reproductive organ and young tissues show constrained elemental composition in Arabidopsis thaliana[J]. Ann Bot, 117(3):431.
YAN ZB, JIN NY, HAN TS, et al., 2013. Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana [J]. Chin J Plant Ecol, 37(6):551-557.[嚴(yán)正兵, 金南瑛, 韓廷申, 等, 2013. 氮磷施肥對(duì)擬南芥葉片碳氮磷化學(xué)計(jì)量特征的影響[J]. 植物生態(tài)學(xué)報(bào), 37(6):551-557.]
YANG M, WANG CQ, YUAN DG, et al., 2015. C, N, P stoichiometry traits of different flue-cured tobacco organs at different growth stages [J]. Chin J Eco-Agric, 23(6):686-693.[楊梅, 王昌全, 袁大剛, 等, 2015. 不同生長(zhǎng)期烤煙各器官C、N、P生態(tài)化學(xué)計(jì)量學(xué)特征 [J]. 中國(guó)生態(tài)農(nóng)業(yè)學(xué)報(bào), 23(6):686-693.]
YU Q, CHEN QS, ELSER JJ, et al., 2010. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability[J]. Ecol Lett, 13:1390-1399.
YU Q, ELSER JJ, HE NP, et al., 2011. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland[J]. Oecologia, 166:1-10.
YU Q, WILCOX K, LA PK, et al., 2015. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change[J]. Ecology, 96(9):2328-2335.
YU Q, WU H, HE NP, et al., 2012. Testing the growth rate hypothesis in vascular plants with above and below-ground biomass[J]. PLoS ONE, 7: e32162.
ZENG DH, CHEN GS, 2005. Ecological stoichiometry: A science to explore the complexity of living systems[J]. Chin J Plant Ecol, 29(6):1007-1019.[曾德慧, 陳廣生, 2005. 生態(tài)化學(xué)計(jì)量學(xué):復(fù)雜生命系統(tǒng)奧秘的探索[J]. 植物生態(tài)學(xué)報(bào), 29(6):1007-1019.]
ZENG DP, JIANG LL, ZENG CS, et al., 2013. Reviews on the ecological stoichiometry characteristics and its applications[J]. Acta Ecol Sin, 33(18):5484-5492.[曾冬萍, 蔣利玲, 曾從盛, 等, 2013. 生態(tài)化學(xué)計(jì)量學(xué)特征及其應(yīng)用研究進(jìn)展[J]. 生態(tài)學(xué)報(bào), 33(18) :5484-5492.]
ZHANG K, SU YZ, WANG T, et al., 2017. Leaf nitrogen and phosphorus stoichiometry of threedesert plants in the same habitat[J]. J Xuchang Univ,(5):116-121.[張珂, 蘇永中, 王婷, 等,2017. 同一生境下三種荒漠植物葉片及土壤氮、磷化學(xué)計(jì)量特征研究[J]. 許昌學(xué)院學(xué)報(bào),(5):116-121.]
ZHANG LX, BAI YF, HAN XG, 2003. Application of N∶P stoichiometry to ecology studies[J]. Acta Bot Sin, 45(9):1009-1018.
ZHANG RY, SHI XM, LI WJ, et al., 2015. Response of species homeostasis and biomass on a sub-alpine grassland[J]. Pratac Sci, 32(10):1539-1547.[張仁懿, 史小明, 李文金, 等, 2015. 亞高寒草甸物種內(nèi)穩(wěn)性與生物量變化模式[J]. 草業(yè)科學(xué), 32(10):1539-1547.]
ZHAO J, ZHANG RP, LI XE, et al., 2011. Leaf nitrogen and phosphorus stoichiometry in Potentilla fruticosa L. affected by internal and external factors in the subalpine in Gannan[J]. J Lanzhou Univ, 47(2):88-92.[趙君, 張仁彭, 李新娥, 等, 2011. 甘南亞高寒草甸金露梅葉片氮磷化學(xué)計(jì)量學(xué)動(dòng)態(tài)[J]. 蘭州大學(xué)學(xué)報(bào), 47(2):88-92.]
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
