茂蘭喀斯特森林不同地形部位優勢喬木種群的生態位研究
2019-09-10 07:22:44秦隨濤龍翠玲吳邦利
廣西植物 2019年5期
秦隨濤 龍翠玲 吳邦利






摘 要:為了解茂蘭喀斯特森林優勢喬木種群的生態位特征,該研究對茂蘭國家級喀斯特森林自然保護區三種地形(坡地、槽谷、漏斗)進行了群落調查,并計算三種地形部位優勢喬木種群的生態位寬度、生態位重疊等特征。結果表明:不同地形部位的優勢喬木種群的生態位寬度Bi和Ba值的排列順序雖稍有差異,但總體上是一致的。輪葉木姜子(Litsea verticillata)在坡地、槽谷地形中的生態位寬度較大,其Bi/Ba分別為0.782 4/0.415 7、0.891 3/0.703 0;齒葉黃皮(Clausena dunniana)在坡地、漏斗地形中有較大的生態位寬度,其Bi/Ba分別為0.788 0/0.518 3、0.962 7/0.826 0。生態位重疊值(Lhi)最大的是漏斗中的樸樹(Celtis sinensis)和云貴鵝耳櫪(Carpinus pubescens)(0.138 2),最小的是坡地中的圓葉烏桕(Triadica rotundifolia)和巴東莢蒾(Viburnum henryi)(0.001 5)、槽谷中的輪葉木姜子和青岡(Cyclobalanopsis glauca)(0.001 5)。生態學特性和對生境需求相似的物種,生態位重疊值通常較大,反之則小;物種的生態位寬度較大,則其生態位重疊值亦較大,反之則小。這說明三種地形中群落優勢種群間的生態位重疊程度較小,茂蘭喀斯特森林處于穩定的頂極群落階段,但群落內種群因環境資源有限而產生的競爭較為激烈,在不同地形生境中的競爭激烈程度表現為漏斗>槽谷>坡地,光照的差異是影響各地形部位優勢喬木種群天然分布的主要因子。
關鍵詞: 喀斯特森林, 不同地形, 優勢種群, 生態位, 茂蘭
中圖分類號:Q948.12
文獻標識碼:A
文章編號:1000-3142(2019)05-0681-09
Niche characteristics of dominant tree populations inkarst forest at different topography sites in MaolanNational Natural Reserve of Guizhou Province
QIN Suitao, LONG Cuiling*, WU Bangli
( College of Geography and Environmental Sciences, Guizhou Normal University, Guiyang 550001, China )
Abstract:In order to understand the niche characteristics of dominant tree populations in karst forest, community investigation was conducted on three topography sites (hillside, valley and funnel) in Maolan National Natural Reserve. Niche characteristics of dominant tree populations at three topography sites were analyzed by niche breadth and niche overlapformula. The results were as follows: The niche breadths Biand Ba of the dominant tree populations at different topographical sites were slightly different, but overall, they were consistent. The niche breadth of Litsea verticillata in hillside and valley was larger, and its Bi/Bawere 0.782 4/0.415 7, 0.891 3/ 0.703 0, respectively. The niche breadth of Clausena dunniana in hillside and funnel was larger, and its Bi/Bawere 0.788 0/0.518 3, 0.962 7/0.826 0, respectively. The niche overlap between Celtis sinensis and Carpinus pubescensin funnel was the largest, being 0.138 2, while those between Viburnum henryi and Triadica rotundifoliain hillside, Litsea verticillata and Cyclobalanopsis glauca in valley were the smallest, being 0.001 5. In addition, species with similar ecological characteristics and habit requirements often had high degrees of niche overlap, species with different ecological characteristics and habit requirements often had low degrees of niche overlap. Species with wide niche breadth often had high degrees of niche overlap, species with narrow niche breadth often had low degrees of niche overlap. In a word, low degrees of niche overlap of dominant species were found at different topography sites and therefore, Maolan karst forest is in a stable climax community stage. But it suggested fierce interspecies competition among dominant species for limited environmental resources in climax community in the forest. The intensity of competition of dominant species at different topography sites is funnel> valley> hillside, light is the main factor affecting the natural distribution of dominant arbor populations.
Key words: karst forest, different topographies, dominant population, niche, Maolan
生態位是種群生態研究中的核心問題,不僅能夠反映種群在所處生態環境中的適應能力與對環境資源的利用能力,還可反映種群在群落或生態系統中所發揮功能和所處位置(冶民生等,2006)。作為評價種間關系和種內關系的重要手段之一,生態位研究在生物多樣性及其形成、維持機制(楊利民等,2001)、群落結構與演替(蘇志堯等,2003)以及森林資源保護與利用(史小華等,2007)等領域得到廣泛運用。目前,學者對處于高寒草甸(魏斌等,2017)、熱帶(陳瑞國等,2017)、亞熱帶(胡正華等,2009)和溫帶森林(劉巍和曹偉,2011)中植物種群生態位進行了深入研究并取得了豐碩的成果,研究對象包括喬木、灌木、草本及藤本植物;有關喀斯特森林植物種群生態位也有見報道,如廣西木論鐵欖群落優勢種群的生態位研究(鐘軍弟等,2009),茂蘭喀斯特森林不同演替階段優勢喬木種群生態位研究(覃弦等,2017)等,但相對而言,對喀斯特森林植物種群生態位的研究較少,因此相關研究亟待加強。
作為一類特殊的亞熱帶森林生態系統,茂蘭喀斯特森林在森林生境、植物區系組成、種群演替更新動態、森林群落性質等方面都與常態地貌上的森林植被有顯著不同(周政賢,1987)。目前,學者對茂蘭喀斯特森林中植物群落數量特征(劉映良和薛建輝,2005)、種間分離特征(張忠華等,2010)、植物種間聯結性(鄭振宇和龍翠玲,2014)、樹種空間分布格局(張忠華等,2015)等方面開展了大量研究,其工作為深入研究喀斯特地區森林群落特征奠定了基礎。茂蘭喀斯特森林地形地貌復雜多樣,有洼地、漏斗、槽谷等地貌形態,不同地形中土壤、水分、光照等環境因子差異較大,各植物種群在復雜地形與高度異質性的小生境中經過長期的適應,對環境資源的利用情況產生了差異,從而具有不同的生態位。在喀斯特森林中,喬木廣泛分布于各種地形中,種類豐富且數量眾多,在森林群落演替、種群進化和群落結構穩定等方面具有重要意義。因此,為深入探討茂蘭喀斯特森林主要喬木種群對高異質性生境的適應能力和對環境資源的利用狀況的差異,了解其在不同地形群落中所處的地位,本文以茂蘭喀斯特森林優勢喬木種群為研究對象,分析其在不同地形中的生態位特征,為茂蘭喀斯特森林的合理保護和退化喀斯特森林植被恢復提供參考依據。
1 研究區概況
研究地位于貴州省荔波縣黔、桂交界處的茂蘭喀斯特森林自然保護區(107°52′10″—108°05′40″E,25°09′20″—25°20′50″ N),其東、南面與廣西省環江縣毗鄰,總面積約20 000hm2,屬中亞熱帶季風濕潤氣候。該區≥10 ℃的年積溫為5 767.9℃,年平均溫為15.3 ℃,氣溫年較差為18.3 ℃,生長期315 d,年降水量為1 752.5mm,年均相對濕度為80%,雨量充沛、濕度大,有利于林木生長發育。茂蘭喀斯特森林母巖以石灰巖和白云巖為主。土壤類型主要為黑色石灰土,其有機質、氮、磷等豐富,呈弱堿性。茂蘭主要植被類型為常綠落葉闊葉混交林,有少部分針闊混交林。森林內喬木層主要物種為輪葉木姜子(Litsea verticillata)、青岡(Cyclobalanopsis glauca)、四照花(Cornus kousasubsp. chinensis)、狹葉潤楠(Machilus rehderi)等;灌木層為皺葉海桐(Pittosporum crispulum)、裂果衛矛(Euonymus dielsianus)、革葉鐵欖(Sinosideroxylon wightianum)、南天竹(Nandina domestica)等。
2 研究方法
2.1 樣方設置及群落調查
于2016年6月植物生長季開展野外群落調查,分別在保護區坡地、槽谷和漏斗三種地形部位(表1)的典型地段中設置20 m×20 m不連續樣地10個,共30個,總面積1.2 hm2(朱守謙和魏魯明,1993)。記錄每個樣地的海拔高度、坡向、坡度、土壤厚度及覆蓋度、水分和光照條件等環境因子。采用常規的群落調查方法(楊允菲和祝廷成,2011),在每個樣地中設置16個5 m×5 m的小樣方,調查樣方內所有喬、灌木,記錄其種名、數量、高度、胸徑、冠幅等指標。
2.2 計算方法
(1)重要值的計算:重要值=(相對密度+相對顯著度+相對頻度)/3(1)
統計坡地(1-10號樣地)、槽谷(11-20號樣地)、漏斗(21-30號樣地)三種地形的群落調查數據,計算種群重要值,根據各種群重要值的排序,確定14個種群作為茂蘭喀斯特森林三種地形部位中的優勢喬木種群,其重要值見表2。
(2)生態位寬度:采用Levins和Hurlbert的生態位寬度計算公式進行計算。
Levins生態位寬度Bi=-∑rj=1PijlogPij(2)
式中,Bi為種i的生態位寬度,Pij為種i對第j個資源的利用占它對全部資源利用的頻度,即Pij=nijNi,而Ni=∑rj=1nij,nij為種i在資源j上的優勢度(本文即樣方中物種的重要值),r為資源等級數。上述方程具有值域 [0,logr]。
Hurlbert生態位寬度Ba=Bi-1r-1(3)
式中,Bi=1/∑rj=1P2ij,Ba為生態位寬度,Pij和r的含義同上式,其值域為 [0,1](劉錦霞等,2007)。
(3)生態位重疊:生態位重疊是指在一定資源序列上,兩個物種利用同等級資源而互相重疊的情況,按以下公式進行計算。
Lih=B(L)i∑rj=1Pij×Phj(4)
Lhi=B(L)h∑rj=1Pij×Phj(5)
BLi=1/(r∑rj=1P2ij)(6)
式中,Lih為物種i重疊物種h的生態位重疊指數;Lhi為物種h重疊物種i的生態位重疊指數;B(L)為Levins的生態位寬度指數;BLi和BLh具有值域 [1/r,1];Lih,Lhi具有值域 [0,1](胡正華和于明堅,2005)。
3 結果與分析
3.1 生態位寬度
由表3中物種生態位寬度Bi可知,在坡地中,翅莢香槐(Cladrastis platycarpa)、齒葉黃皮(Clausena dunniana)和輪葉木姜子具有較大的生態位寬度,分別為0.847 4、0.788 0和0.782 4;在槽谷中,輪葉木姜子、香葉樹(Lindera communis)和柿(Diospyros kaki)具有較大的生態位寬度,分別為0.891 3、0.846 6和0.800 1;在漏斗中,齒葉黃皮、樸樹、狹葉潤楠和青岡具有較大的生態位寬度,分別為0.962 7、 0.937 6、 0.904 2和0.894 6。優勢喬學特性等方面有關,如輪葉木姜子具有較強的耐干旱貧瘠能力,分布在坡地中的輪葉木姜子個體較大、數量多,在坡地較干旱的環境中對維護群落的穩定具有重要作用。同時作為耐蔭樹種,在林冠郁閉度較高、水分、土壤和光照良好的槽谷中,輪葉木姜子幼苗能夠進行貯備與更新,因而在坡地與槽谷中的生態位寬度較大。處于林冠上層的陽性落葉樹種翅莢香槐在林內主要為個體大的成樹,只有少量幼苗和幼樹,個體數量少且多分布在坡地中,因而在坡地中生態位寬度較大,而在槽谷和漏斗中生態位寬度較小。
3.2 生態位重疊分析
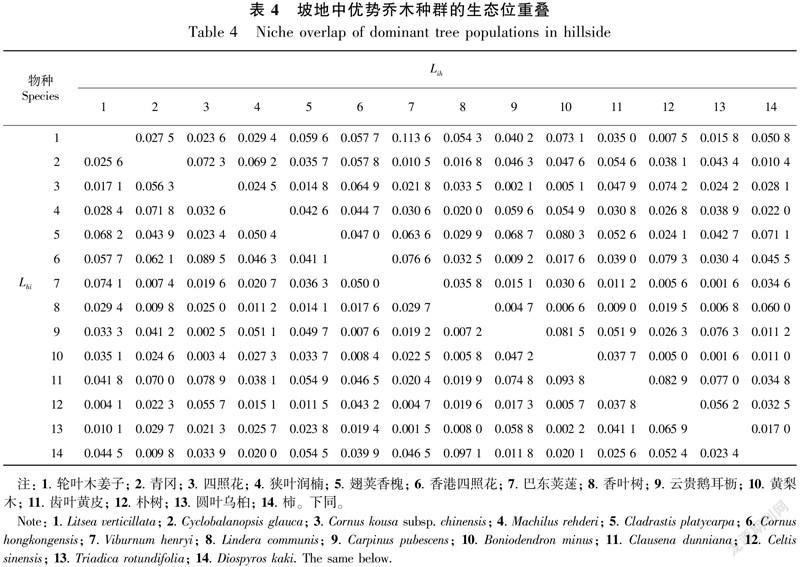
3.2.1 坡地優勢種群生態位重疊分析 由表4可知,在坡地中,優勢喬木種群的生態位重疊值大于等于0.07的有20對(占總數的11.0%),大于等于0.05的有48對(占總數的26.4%)。在坡地中生態位寬度值較大的翅莢香槐—齒葉黃皮生態位重疊值較高, Lih/Lhi值為0.052 6/0.054 9, 生態位寬度木種群在三種地形中的生態位寬度出現差異,這與地形生境特征、種群個體大小、分布數量及生態值較小的樸樹—黃梨木生態位重疊值較低,Lih/Lhi值為0.005 0/0.005 7。翅莢香槐是陽性落葉樹種,齒葉黃皮具有喜陽耐蔭、耐旱的特性,由于種群間對光資源的競爭,兩者在坡地中都多分布于林冠上層,對光照需求強烈而導致生態位重疊;樸樹與黃梨木雖均為喜陽耐蔭樹種,對光照需求強烈,但樸樹在坡地中數量少,且主要分布于林冠上層,而黃梨木在坡地中數量多,但主要分布于林冠中下層,種群在林冠產生了垂直分布的差異,彼此生態位發生分離,因而二者在坡地中的競爭并不激烈。
3.2.2 槽谷優勢種群生態位重疊分析 由表5可知,在槽谷中,優勢喬木種群的生態位重疊值大于等于0.07的有31對(占總數的17.0%),大于等于0.05的有63對(占總數的34.6%)。輪葉木姜子、香葉樹和柿的生態位寬度在槽谷中排前三位,三者之間的生態位重疊值高低各異,其中輪葉木姜子—香葉樹、輪葉木姜子—柿的Lih/Lhi值分別為0.054 9/0.046 5、0.048 4/0.038 3,香葉樹—柿的Lih/Lhi值為0.074 9/0.070 0。生態位寬度值較小的青岡—香港四照花的生態位重疊值也較小,Lih/Lhi值為0.026 0/0.027 9。在槽谷中,林分郁閉度高,種群對光資源的競爭導致輪葉木姜子、香葉樹和柿都分布于林冠中下層,對光照需求強,但輪葉木姜子數量少,個體大,香葉樹與柿的種群數量多,個體較小,因而輪葉木姜子對香葉樹、柿的生態位重疊值較低,而香葉樹與柿的生態位重疊值則較高;青岡在槽谷中主要分布在林冠上層,利用上層空間資源,而常綠耐蔭樹種香港四照花則分布在林冠的中下層,二者在林冠垂直分布上產生了分離,因而生態位重疊值低。
3.2.3 漏斗優勢種群生態位重疊分析 由表6可知,在漏斗中,優勢喬木種群的生態位重疊值大于等于0.07的有46對(占總數的25.3%),大于等于0.05的有99對(占總數的54.4%)。齒葉黃皮、樸樹和狹葉潤楠的生態位寬度值在漏斗中排前三位,三者之間的生態位重疊值都較大,其中齒葉黃皮—樸樹、齒葉黃皮—狹葉潤楠的Lih/Lhi值分別為0.075 8/0.067 8、0.077 1/0.091 7,樸樹—狹葉潤楠的Lih/Lhi值為0.059 6/0.063 4。生態位寬度值小的翅莢香槐—云貴鵝耳櫪生態位重疊值也小,Lih/Lhi值為0.004 8/0.004 3。齒葉黃皮、樸樹為喜陽樹種,狹葉潤楠為耐蔭樹種,漏斗地形中的光照條件差,三個種群對光照需求強烈且都分布于林冠上層,因而對光因子的競爭最激烈;云貴鵝耳櫪分布在林冠中下層,與分布在林冠上層的翅莢香槐生態位發生分離,因而生態位重疊值小。
綜上所述,Lih最大的是坡地中的輪葉木姜子—巴東莢蒾,為0.113 6,輪葉木姜子與巴東莢蒾都具有喜陽、耐干旱貧瘠的特性,兩者都主要分布在光照較好的坡地和槽谷地形的林冠中下層,兩者因利用生境因子而產生的競爭較激烈,生態位重疊值大。Lhi最大的是漏斗中的樸樹—云貴鵝耳櫪,為0.138 2,樸樹與云貴鵝耳櫪都為喜陽樹種,兩個種群在漏斗中分布數量較多,但漏斗中的光照最差,因而兩者對光照因子的競爭也最為激烈。Lih最小的是坡地中的圓葉烏桕—巴東莢蒾、黃梨木,為0.001 6。Lhi最小的是坡地中的圓葉烏桕—巴東莢蒾和槽谷中的輪葉木姜子—青岡,都為0.001 5,圓葉烏桕雖為喜陽耐旱樹種,但主要分布于光照弱、土層較厚且濕度大的漏斗中,與巴東莢蒾對生境的要求相差太大,兩者產生了生態位分離,因而生態位重疊值很低;槽谷中的輪葉木姜子分布于林冠中下層,青岡則分布在林冠上層,二者發生林冠垂直分離,因而生態位重疊值低。總體來看,14個優勢喬木種群在三種地形中生態位重疊值大于等于0.07的共有97對(占總數的17.8%),生態位重疊值在0.001 5~0.138 2之間波動,三種地形中所有的Lih值和Lhi值平均為0.043 0,群落14個優勢喬木種群在三種地形中的生態位重疊值很低,表明茂蘭喀斯特森林群落穩定性較大,已演替到頂極群落階段。
4 討論
物種對生境的適應情況和對資源的利用能力可通過生態位寬度表征。在環境資源缺少的條件下,物種的生態位寬度往往較大,才能使種群獲得更多的資源,利用環境資源的能力更強,更能適應惡劣的環境(吳友貴等,2016)。坡地中的光照強,但坡度較大,水分及土壤易流失,林內灌木較豐富,喬木主要為成樹,適宜耐旱的樹木生長,生態位寬度排前三位的翅莢香槐、齒葉黃皮和輪葉木姜子,在坡地中主要為成樹,它們都具有較強的耐干旱能力,說明它們能夠較好地適應坡地干旱貧瘠的環境。槽谷的生存環境最好,既有適中的光照和水分條件,也有較厚的土壤層,所以喬灌木生長良好,輪葉木姜子、香葉樹和柿的生態位寬度在槽谷中排前三位,這可能與三個種群在群落中的數量、分布特點等有關,輪葉木姜子雖然主要分布在坡地中,但生長在槽谷中的數量也較多,而香葉樹和柿的幼樹則主要分布在槽谷中。漏斗中生存環境較差,主要是光照時間短,低凹地段有積水,土層較厚但粘重,更適合幼苗、幼樹和藤本植物生長,在漏斗中,齒葉黃皮、樸樹、狹葉潤楠和青岡的生態位寬度排前四位,它們都主要以幼苗、幼樹的形式生長在漏斗中。從樹木生長不同階段和生態環境綜合考慮,認為槽谷是樹木最適生境,但對如齒葉黃皮、青岡等喜陽耐蔭種群而言,當它們處于幼苗、幼樹階段時,水分充足、蔭蔽的漏斗比較適宜其生長,而當其成長為成樹時,光照充足的坡地則更適宜生長。
當兩個物種共同利用同一資源或占有某一資源因素(生存空間、食物等)時,就會出現生態位重疊現象(王伯蓀等,1995;胡正華和于明堅,2005)。種群的生態位重疊值較大的原因通常是因為它們之間有相近的生態特性或者對生境因子有互補性要求(林思祖等,2002;冶民生等,2006)。物種的生態位寬度與生態位重疊之間的關系復雜,通常具有較大生態位寬度種群的生態位重疊通常也較大,反之則小。如翅莢香槐和齒葉黃皮在坡地中的生態位寬度較大,其在坡地中的生態位重疊值也較高,而生態位寬度較小的樸樹與黃梨木間的生態位重疊值則較小。這與趙永華等(2004)對秦嶺銳齒櫟林種群的生態位研究所得結論相一致。物種的生態位重疊與物種本身的生物生態學特性有關(張俊欽,2005),因此也有生態位寬度較小的種群之間生態位重疊值較高的情況出現,如漏斗中的樸樹與云貴鵝耳櫪為喜陽耐蔭樹種,且以幼苗、幼樹居多,對土壤養分、光照等資源的需求強烈,故二者間生態位重疊值亦較高。
競爭是在環境資源缺乏且生態位重疊的條件下形成的,生態位重疊與競爭間的關系復雜,環境資源量、供求比和生物對資源需求的滿足程度等對二者的關系十分重要(趙永華等,2004)。總體來看,茂蘭喀斯特森林中優勢喬木種群的生態位重疊值偏低,森林群落穩定性較大,但種間競爭仍較激烈,究其原因,這與喀斯特林內生境惡劣,資源不足有關。通過對茂蘭喀斯特森林三種地形優勢喬木種群的生態位重疊的分析可以發現,坡地中優勢喬木種群的生態位重疊值大于等于0.05的有48對(占總數的26.4%),槽谷、漏斗中則分別為63對(占總數的34.6%)和99對(占總數的54.4%),可見種群在不同地形部位生境中的競爭激烈程度為漏斗>槽谷>坡地。綜合優勢喬木種群生長特性、個體大小、生態位重疊及生態環境的探討,種群在漏斗中競爭最強,槽谷次之,坡地最弱,而光照強度在三種地形的分布為坡地最強,槽谷次之,漏斗最弱,因此認為光照是影響茂蘭喀斯特森林優勢喬木種群天然分布的主要因子,與影響茂蘭喀斯特森林優勢灌木種群分布的主要因子相同(秦隨濤等,2018)。
參考文獻:
CHEN RG, FAN SH, LIU GL, et al., 2017. Niche characteristics of dominant species of rattan accompanying community in secondary lowlan rain forest in Hainan Island, China [J]. Acta Bot Boreal-Occident Sin, 37(6):1226-1233. [陳瑞國, 范少輝, 劉廣路, 等, 2017. 海南島次生低地雨林棕櫚藤伴生群落優勢種生態位研究 [J]. 西北植物學報,37(6):1226-1233.]
HU ZH, QIAN HY, YU MJ, 2009. The niche of dominant species populations in Castanopsis eyreiforest in Gutian Mountain National Nature Reserve [J]. Acta Ecol Sin, 29(7): 3670-3677. [胡正華, 錢海源, 于明堅, 2009. 古田山國家級自然保護區甜櫧林優勢種群生態位 [J]. 生態學報,29(7): 3670-3677.]
HU ZH, YU MJ, 2005. Niche characteristics of dominant populations in Cyclobalanopsis glauca forest in Gutian Mountain [J]. Chin J Ecol, 24(10): 1159-1162. [胡正華,于明堅,2005. 古田山青岡林優勢種群生態位特征 [J]. 生態學雜志,24(10): 1159-1162.]
LIU W, CAO W, 2011. Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains [J]. Chin J Ecol, 30(8): 1766-1774.[劉巍, 曹偉, 2011. 長白山云冷杉群落主要種群生態位特征 [J]. 生態學雜志, 30(8): 1766-1774.]
LIU YL, XUE JH, 2005. Quantitative properties of degraded karst forest communities in Maolan mountain area of Guizhou [J]. J Nanjing For Univ(Nat Sci Ed), 29(3): 23-27.[劉映良, 薛建輝, 2005. 貴州茂蘭退化喀斯特森林群落的數量特征 [J]. 南京林業大學學報(自然科學版), 29(3): 23-27.]
LIU JX, WU GL, MA T, 2007. Study on niche characteristics of desert plant community in marginal zone of Muus Desert [J]. Pratac Sci,24(12): 9-14. [劉錦霞, 武高林, 馬濤, 2007. 毛烏素沙漠邊緣荒漠植物群落生態位特征研究 [J]. 草業科學, 24(12): 9-14.]
LIN SZ, HUANG SG, HONG W, et al., 2002. The characteristics of Multi-dimension niche of dominant populations in Chinese fir and Broad-leaved mixed forest [J]. Acta Ecol Sin, 22(6): 962-968. [林思祖, 黃世國, 洪偉, 等, 2002. 杉闊混交林主要種群多維生態位特征 [J]. 生態學報, 22(6):962-968.]
QIN X, LONG CL, LI J, 2017. Analysis of ecological niches of dominant arbor populations in different succession stages in the karst forest in Maolan Nature Reserve [J]. J SW Univ(Nat Sci Ed), 39(6): 42-49.[覃弦, 龍翠玲, 李娟, 2017. 茂蘭喀斯特森林不同演替階段優勢喬木種群生態位研究 [J]. 西南大學學報(自然科學版), 39(6): 42-49.]
QIN ST, LONG CL, WU BL, 2018. Study on niche of dominant shrub populations in karst forest in Maolan Nature Reserve, Guizhou Province [J]. For Res Manag, (2): 58-64, 90.[秦隨濤, 龍翠玲, 吳邦利, 2018. 茂蘭喀斯特森林優勢灌木種群的生態位研究 [J]. 林業資源管理,(2): 58-64,90.]
SHI XH, XU XB, ZHANG WH, 2007. Study on the niche of the main populations in Abies chensiens community [J]. Bull Bot Res, 27(3): 345-349.[史小華, 許曉波, 張文輝, 2007. 秦嶺冷杉群落主要種群生態位研究 [J]. 植物研究, 27(3): 345-349.]
SU ZY, WU DR, CHEN BG, 2003. Niche characteristics of dominant populations in natural forest in North Guangdong [J]. Chin J Appl Ecol, 14(1): 25-29. [蘇志堯, 吳大榮, 陳北光, 2003. 粵北天然林優勢種群生態位研究 [J]. 應用生態學報, 14(1): 25-29.]
WEI B, LU N, LI JQ, et al., 2017. Effects of enclosure on plant community composition and niche characteristics in alpine meadow [J]. Acta Bot Boreal-Occident Sin, 37(5): 983-991. [魏斌, 陸妮, 李佳琪, 等, 2017. 封育對高寒草甸植物群落構成及生態位特征的影響 [J]. 西北植物學報, 37(5): 983-991.]
WANG BS, LI MG, PENG SL, 1995. Phytopopulology [M]. Guangzhou: Guangdong Higher Education Press: 132-148.[王伯蓀, 李鳴光, 彭少麟, 1995. 植物種群學 [M]. 廣州廣東高等出版社: 132-148.]
WU YG, YE ZL, ZHOU RF, et al., 2016. Niche of dominant species populations in an evergreen broad-leaved forest in Baishanzu [J]. Guihaia,36(2): 186-192. [吳友貴, 葉珍林, 周榮飛, 等, 2016. 百山祖常綠闊葉林優勢種群的生態位 [J]. 廣西植物, 36(2): 186-192.]
YE MS, GUAN WB, WU B, et al., 2006. Niche characteristics of main shrub populations in the arid valley of the Minjiang River, southwestern China [J]. J Beijing For Univ, 28(1): 7-13. [冶民生, 關文彬, 吳斌, 等, 2006. 岷江干旱河谷主要灌木種群生態位研究 [J]. 北京林業大學學報, 28(1):7-13.]
YANG LM, ZHOU GS, WANG GH, 2001. Investigating realized niches as a mechanism of species diversity maintenance in a species-rich grassland community [J]. Acta Phytoecol Sin, 25(5): 634-638.[楊利民, 周廣勝, 王國宏, 2001. 草地群落物種多樣性維持機制的研究Ⅱ物種實現生態位 [J]. 植物生態學報, 25(5): 634-638.]
YANG YF, ZHU TC, 2011. Plant ecology[M]. 2nd ed. Beijing: Higher Education Press: 29-31.[楊允菲, 祝廷成, 2011. 植物生態學(第二版) [M]. 北京: 高等教育出版社: 29-31.]
ZHAO YH, LEI RD, HE XY, et al., 2004. Niche characteristics of plant populations in Quercus aliena var. acuteserrata stands in Qinling Mountains [J]. Chin J Appl Ecol, 15(6): 913-918.[趙永華, 雷瑞德, 何興元, 等, 2004. 秦嶺銳齒櫟林種群生態位特征研究 [J]. 應用生態學報, 15(6): 913-918.]
ZHU SQ, WEI LM, 1993. Studies on structure of Maolan karst forest communities [M] // ZHU SQ. Ecological research on karst forest(Ⅰ). Guiyang: Guizhou Publishing House of Science and Technology: 12-21.[朱守謙, 魏魯明, 1993. 茂蘭喀斯特森林群落結構研究//朱守謙. 喀斯特森林生態研究(I)(M). 貴陽: 貴州科技出版社: 12-21.]
ZHANG ZH, HU G, NI J, 2010. Interspecific segregation of old-growth Karst forests in Maolan, Southwest China [J]. Acta Ecol Sin, 30(9): 2235-2245.[張忠華, 胡剛, 倪健, 2010. 茂蘭喀斯特森林群落的種間分離特征 [J].生態學報, 30(9): 2235-2245.]
ZHANG ZH, HU G, NI J, 2015. Spatial distribution patterns and their fractal properties for trees in a subtropical mixed evergreen-deciduous broad-leaved karst forest in Maolan, southwestern China [J]. Acta Ecol Sin, 35(24): 8221-8230. [張忠華, 胡剛, 倪健, 2015. 茂蘭喀斯特常綠落葉闊葉混交林樹種的空間分布格局及其分形特征 [J]. 生態學報, 35(24): 8221-8230.]
ZHANG JQ, 2005. Main population niche studyof Phoebe bourneinatural forest in Mingxi, Fujian [J]. J Fujian For Sci Technol, 32(3):31-35. [張俊欽, 2005. 福建明溪閩楠天然林主要種群生態位研究 [J]. 福建林業科技, 32(3):31-35.]
ZHENG ZY, LONG CL, 2014. Interspecific association of woody plant species at different topography sites in Maolan karst forest[J]. For Res Manag, (4): 78-84.[鄭振宇, 龍翠玲, 2014. 茂蘭喀斯特森林不同地形部位木本植物種間聯結性分析 [J]. 林業資源管理, (4): 78-84]
ZHOU ZX, 1987. A comprehensive report on the survey of Maolankarst forest [M]// ZHOU ZX. Scientific survey of the Maolan karst forest. Guiyang: Guizhou People’s Publishing House: 1-23.[周政賢, 1987. 茂蘭喀斯特森林科學考察綜合報告 [M]//周政賢. 茂蘭喀斯特森林科學考察集. 貴陽: 貴州人民出版社: 1-23.]
ZHONG JD, LI XK, YE D, et al., 2009. Study on niches of dominant populations of Sinosideroxylon pedunculatumcommunity in Mulun National Nature Reserve of Guangxi [J]. J Plant Res Environ, 18(3): 38-43.[鐘軍弟, 李先琨, 葉鐸, 等, 2009. 廣西木論國家級自然保護區鐵欖群落優勢種群的生態位研究 [J]. 植物資源與環境學報, 18(3): 38-43.]
