3-磷酸甘油醛脫氫酶促進谷氨酸棒桿菌發酵生產L-精氨酸和L-鳥氨酸
2019-08-26 03:24:52闞寶軍董晉軍劉暉占米林許國超韓瑞枝倪曄
食品與發酵工業 2019年15期
關鍵詞:產量
闞寶軍,董晉軍,劉暉,占米林,許國超,韓瑞枝,倪曄*
1(江南大學 生物工程學院,生物催化與酶工程研究室,江蘇 無錫,214122) 2(工業生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)
L-鳥氨酸(L-Ornithine)和L-精氨酸(L-Arginine)作為尿素循環的中間代謝產物,具有多種重要的生理生化功能,在食品、醫藥和保健品等領域具有廣泛的應用[1]。L-精氨酸是合成肌酸、蛋白質的重要原料,是體內信號分子NO的唯一前體,可用于治療心絞痛、神經性疾病和肝硬化等[2]。L-鳥氨酸有益于創傷、燒傷、感染甚至癌癥等疾病的恢復,對肝病也有治療作用[3-4],還具有良好的減肥保健作用[5]。L-精氨酸的生產方法主要有毛發水解液提取和微生物發酵法。目前,L-鳥氨酸可以通過精氨酸酶水解L-精氨酸和微生物好氧發酵2種方法來生產[6]。
谷氨酸棒桿菌(Corynebacteriumglutamicum)是一種環境友好的非致病性革蘭氏陽性土壤細菌,已廣泛應用于多種有機酸和氨基酸的工業生產[7]。在谷氨酸棒桿菌中,合成氨基酸通常需要消耗大量的NADPH。在L-精氨酸合成途徑中每生產1 molL-精氨酸,需要消耗3 mol的NADPH[8],對于L-鳥氨酸的合成,至少需要消耗2 mol NADPH才能合成1 molL-鳥氨酸[9],因此,胞內NADPH的供給被認為是L-精氨酸及L-鳥氨酸合成的限制因素。
TAKENO等[10]用變異鏈球菌來源的NADP+依賴型甘油醛-3-磷酸脫氫酶(GAPDH)替代內源性NAD+依賴型甘油醛-3-磷酸脫氫酶,來改善谷氨酸棒桿菌NADPH水平和提高L-賴氨酸產量。郭雯等[11]通過加強胞內NAD+依賴型的GAPDH表達,L-絲氨酸的產量和生產強度均提高了17.4%[11],說明GAPDH是谷氨酸棒桿菌中的限速酶,通過增加該酶的表達可以增強糖酵解途徑代謝流。
谷氨酸棒狀桿菌SNK118(C.glutamicumSNK118)是本實驗室構建的L-精氨酸生產菌株[12]。如圖1所示,在谷氨酸棒桿菌中,L-谷氨酸可以通過由ArgC、ArgJ、ArgB和ArgD組成的代謝途徑生成L-鳥氨酸,再由ArgF、ArgG和ArgH 3個酶催化生成L-精氨酸。采用CRISPR-Cpf1技術[13]敲除argF基因以積累L-鳥氨酸,通過敲除argR(精氨酸操縱子阻遏蛋白基因[14])可增加L-鳥氨酸產量。本研究擬在谷氨酸棒桿菌中重組表達梭菌來源的NADP+依賴型的GAPDH編碼基因,以增加糖酵解途徑代謝流,同時提高胞內NADPH含量,提高L-精氨酸以及L-鳥氨酸的產量。
1 材料與方法
1.1 材料
1.1.1 菌種、質粒及引物
本研究中所涉及到的菌株與質粒和引物及其相關性質等描述見表1和表2。

表1 本文所用的菌株與質粒

表2 本研究所用的引物
1.1.2 試劑
非連接酶依賴型單片段快速一步克隆試劑盒,南京諾唯贊生物科技有限公司;Gibson無縫連接試劑盒,NEB公司;DL-三磷酸甘油醛,Sigma-Aldrich公司;限制性核酸內切酶、Primer STAR DNA聚合酶,Takara寶生物工程有限公司;其他試劑,國藥集團(上海)化學試劑,有限公司。
1.1.3 培養基
(1)谷氨酸棒桿菌活化用BHI培養基(g/L):NaCl 5,蛋白胨10,腦心浸液18.5,酵母粉5,pH值調至7.2~7.4。固體培養基添加17的瓊脂粉,115 ℃滅菌20 min。
(2)谷氨酸棒狀桿菌感受態細胞制備用EPO培養基(g/L):蛋白胨10,酵母粉 5,NaCl 10,甘氨酸 25,吐溫-80 1,異煙肼 0.4,腦心浸液18.5,121 ℃滅菌20 min。
(3)谷氨酸棒狀桿菌電轉恢復用LB-HIS培養基(g/L):NaCl 5,蛋白胨20,腦心浸液18.5,酵母粉10,D-山梨醇91,pH值調至7.2~7.4。固體培養基添加17 g/L的瓊脂粉,121 ℃滅菌20 min。
(4)種子培養基:葡萄糖30 g/L(115 ℃滅菌16 min,分消),生物素 50 μg/L(濾膜過濾除菌),玉米漿干粉25 g/L,(NH4)2SO45 g/L,尿素 0.8 g/L,MgSO4·7H2O 0.5 g/L,KH2PO41.5 g/L,K2HPO4·3H2O 0.5 g/L,pH值調至7.2~7.4,121 ℃滅菌20 min。
(5)發酵培養基:KH2PO41.5 g/L,K2HPO4·3H2O 0.5 g/L,尿素0.75 g/L,MgSO4·7H2O 1 g/L,玉米漿干粉20 g/L,(NH4)2SO440 g/L,MnSO4·H2O 22.38 mg/L,FeSO4·7H2O 18.3 mg/L,pH調至7.2~7.4,121 ℃滅菌20 min;葡萄糖100 g/L(115 ℃滅菌15 min,分消),CaCO330 g/L(160 ℃干熱滅菌1.5~2 h),生物素 100 μg/L,維生素B1200 μg/L。裝液量30 mL/500 mL。
1.2 試驗方法
1.2.1 重組表達質粒的構建
本研究共克隆了3個GAPDH編碼基因:CagapC、CbgapC和CsgapC,分別來源于梭菌C.beijerinckiiDSM 1739、C.acetobutylicum1.7和C.saccharobutylicumDSM 13864。以構建pXMJ19-CsgapC的方法為例,首先以pXMJ19為載體質粒,選取HindIII和XbaI雙酶切質粒使其線性化,將酶切產物純化回收。同時以ClostridiumsaccharobutylicumDSM 13864基因組為模板,以CsgapC-F/CsgapC-R為引物,PCR擴增得到CsgapC片段,其中片段兩端帶有質粒的同源臂片段,凝膠電泳驗證并通過膠回收試劑盒純化。將純化好的CsgapC片段與pXMJ19片段通過快速一步克隆試劑盒連接。連接產物轉化至E.coliJM109感受態細胞,通過pXMJ19多克隆位點上下游序列設計的通用引物pXMJ19-F/pXMJ19-R進行菌落PCR,驗證并提取質粒測序確認。質粒pXMJ19-CbgapC和pXMJ19-CagapC的構建參考本方法。重組質粒pXMJ19-CsgapC構建流程見圖1、圖2。
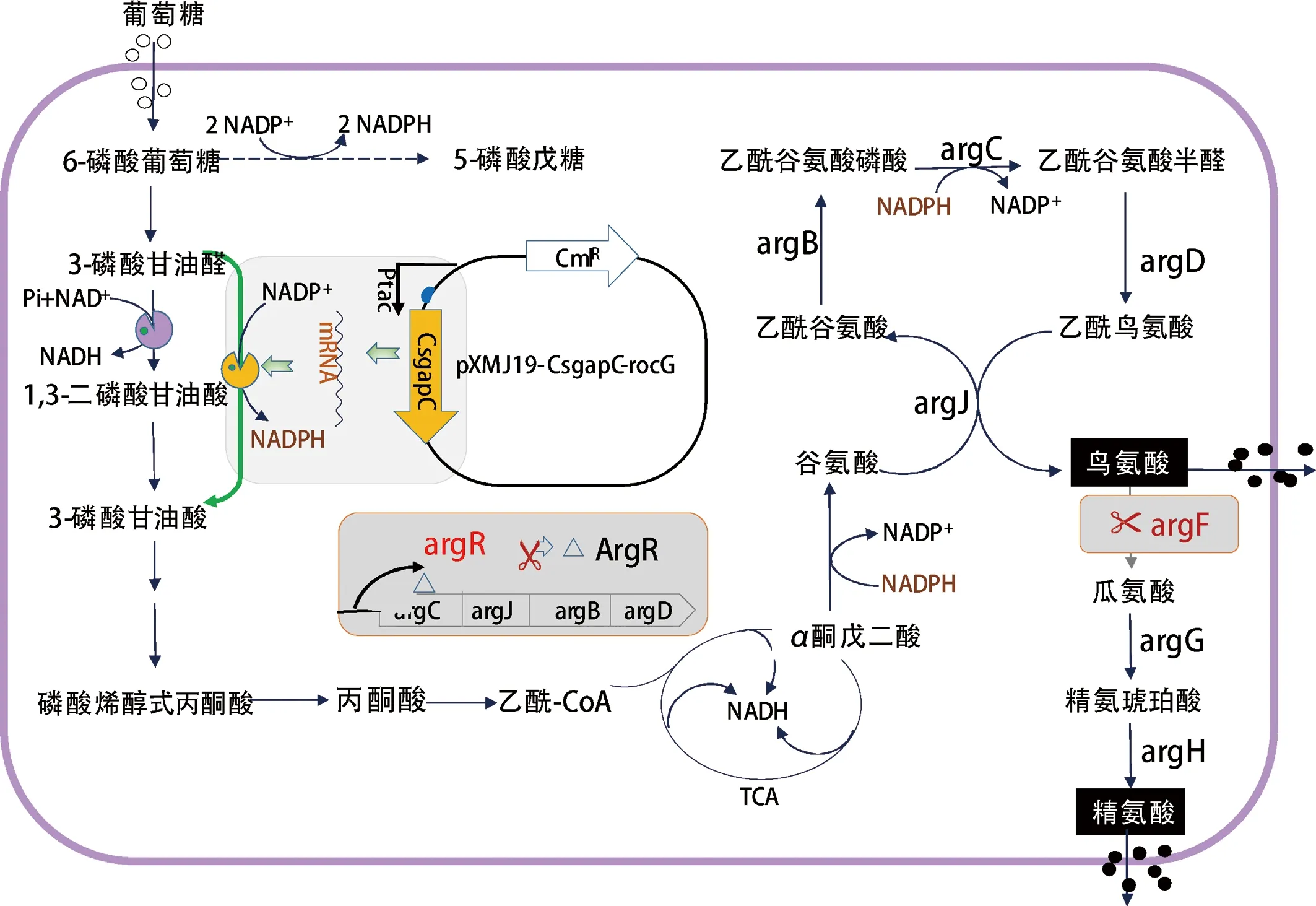
圖1 谷氨酸棒桿菌中L-鳥氨酸和L-精氨酸的合成途徑
Fig.1 The biosynthesis pathway of L-ornithine and L-arginine in C. glutamicum
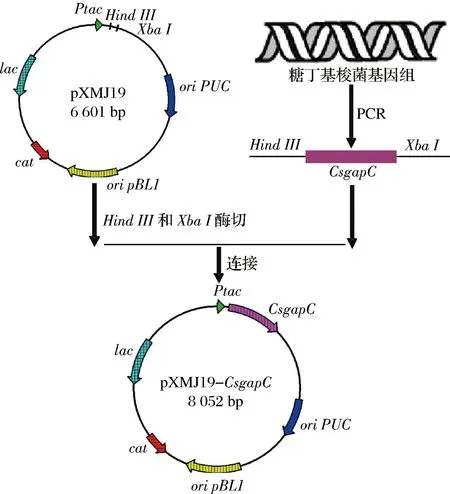
圖2 pXMJ19-CsgapC質粒構建
Fig.2 Construction of pXMJ19-CsgapC plasmid
1.2.2 CRISPR-Cpf1系統基因敲除質粒pJYS3_ΔargFΔargR的構建
基因argF和argR在谷氨酸棒狀桿菌基因組上的位置相距48 bp。首先以質粒pJYS3_ΔcrtYf作為模板,通過引物crRNA_argFR-F/crRNA_argFR-F引入定點突變,即通過全質粒PCR反應將crtYf基因的crRNA序列(GAATTTCTACTGTTGTAGATCAGGCAACCA-TAGGGCAGGAA)替換為argF基因的crRNA序列(GAATTTCTACTGTTGTAGATGTCCC-TCGAGTGGACGCTCCG)。DpnI消化掉PCR產物中的母本質粒,消化液轉入克隆宿主E.coliJM109。測序驗證正確后獲得pJYS3-I質粒。然后,以pJYS3-F/pJYS3-R為引物,pJYS3-I為模板,PCR擴增得到不包含crtYf基因上下游同源臂的pJYS3-H片段。同時以C.glutamicumSNK118基因組為模板,分別以引物argFR-D-F/argFR-D-R與argFR-U-F/argFR-U-R擴增出基因的下游D與上游U同源片段。膠回收純化以上3個目的片段,通過Gibson組裝法[16]將這3個片段進行組裝連接。構建成功獲得質粒pJYS3_ΔargFΔargR。質粒的構建流程如圖3所示。
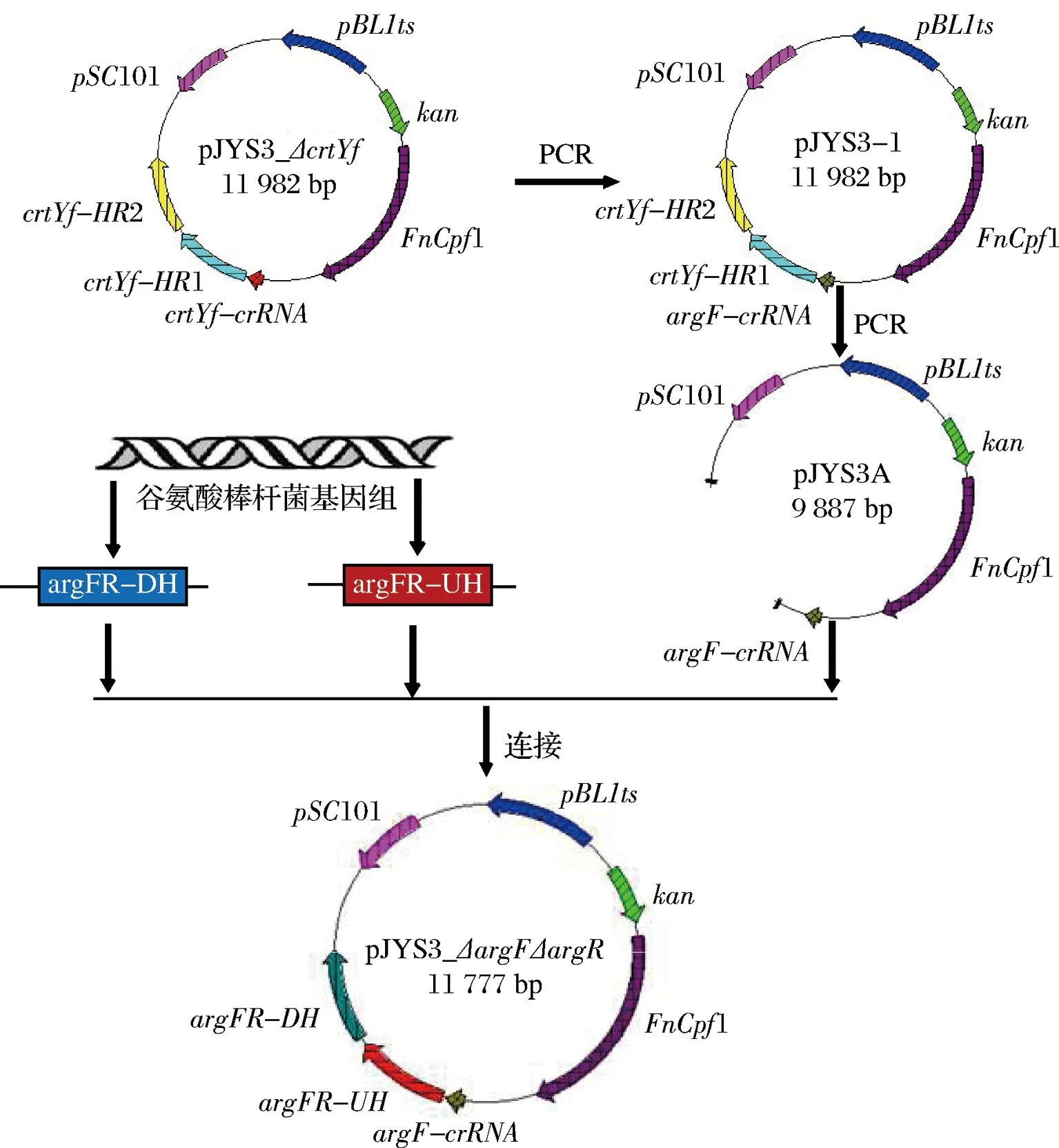
圖3 敲除質粒pJYS3_ΔargFΔargR的構建
Fig.3 Construction of knockout plasmid pJYS3_ΔargFΔargR
1.2.3 異源基因表達重組菌株的構建
(1)重組大腸桿菌的構建
參考SAMBROOK的方法[17],將構建成功的異源基因重組表達質粒轉化到感受態宿主表達細胞E.coliBL21(DE3)中,并涂布于30 mg/L氯霉素平板上,37 ℃培養12 h后菌落PCR驗證。
(2)重組谷氨酸棒狀桿菌的構建
將上述初篩到的2種重組質粒電轉到C.glutamicum感受態細胞中,獲得4株重組谷氨酸棒狀桿菌菌株SNK118/pXMJ19-CagapC、SNK118/pXMJ19-CsgapC、SNK118 ΔargFΔargR/pXMJ19-CagapC、SNK118ΔargFΔargR/pXMJ19-CsgapC。谷氨酸棒桿菌電轉方法與感受態制備見REST的方法[18]。
1.2.4 基因敲除菌SNK118ΔargRΔargF的構建
將成功構建的基因敲除質粒電轉至C.glutamicumSNK118感受態細胞中,均勻涂布于含卡那霉素平板,30 ℃靜置培養3~4 d,培養過程中Cpf1會對crRNA進行加工,成熟的crRNA會引導Cpf1結合到DNA的特異性位點上,對目標基因進行切割,在菌體內重組酶的作用下,通過同源重組將基因argF、argR進行敲除,培養結束后菌落PCR驗證篩選出成功敲除的突變株。對成功敲除的突變株劃線于無抗平板,34 ℃培養16 h以消除pJYS3_ΔargFΔargR。
1.2.5 GAPDH酶活力的測定
在30 ℃條件下,通過分光光度法測量340 nm波長下NADP(或NAD+)還原的吸光值變化來計算GAPDH活力[19]。標準反應混合物含有300 μL 100 mmol/L Tris-HCl(pH 8.5),60 μL 10 mmol/L H3PO4,60 μL 10 mmol/L NADP+(或NAD+),60 μL 10 mmol/L甘油醛3-磷酸,50 μL的β-巰基乙醇和70 μL粗酶液。通過添加細胞破碎粗酶液引發催化反應。1單位GAPDH活力定義為在上述條件下1 min內還原1 μmol NAD(或NADP+)所需的酶量。
使用牛血清蛋白標準液,通過Bradford方法計算蛋白質濃度,按公式(1)計算:

(1)
1.2.6 胞內輔酶水平的測定
30 ℃培養46 h后收獲菌體細胞,并洗滌2次。使用NADP+/NADPH試劑盒(BioAssay Systems,Hayward,CA)抽提并測定胞內NADPH/NADP+的輔酶參數。
1.2.7 菌體濃度、氨基酸濃度、葡萄糖濃度的測定
菌體濃度的測定:取一定量的菌液用0.2 mol/L鹽酸稀釋50~100倍后,用分光光度計在660 nm波長下測定吸光值。
氨基酸濃度測定:采用高效液相色譜(high performance liquid chromatography, HPLC)分析儀進行鄰苯二甲醛(O-phthalaldehyde,OPA)在線衍生化法測定[20]。
葡萄糖濃度的測定:發酵上清液稀釋100倍,采用SBA-40C生物傳感器分析儀進行測定。
2 結果與分析
2.1 NADP+依賴型的GAPDH的克隆與表達篩選
在NCBI數據庫中,通過BLAST搜索,選擇不同梭菌的3個編碼NADP+依賴型的GAPDH基因:CagapC、CbgapC、CsgapC。將這3個基因克隆到穿梭載體pXMJ19中,并在大腸桿菌BL21(DE3)中誘導表達。之后測定細胞提取物中重組GAPDH的比活力,如表3所示。

表3 重組GAPDH在E. coli和C. glutamicum的酶活力
結果顯示,重組CaGapC和CsGapC在30 ℃下的比活力分別為2 000 mU/mg和5 263 mU/mg,而CbGapC沒有顯示出催化活性。基于以上結果,將重組質粒pXMJ19-CagapC、pXMJ19-CsgapC分別轉入SNK118菌株,評估這2個重組酶在谷氨酸棒桿菌中合成NADPH的催化活性。據報道,丙酮丁醇梭菌來源的編碼NADP+依賴型GAPDH的gapC的表達改善了谷氨酸棒桿菌胞內NADPH的供應[21]。然而,在甘油醛-3-磷酸和NADP+作為底物時,只有CsGapC顯示出77 mU/mg的酶活力(表3),表明CsGapC具有催化3-磷酸甘油醛生成3-磷酸甘油酸的活性,同時可以為C.glutamicumSNK118提供額外的NADPH來源。
2.2 重組菌發酵產L-精氨酸性能評價
將重組菌LA4(SNK118/pXMJ19-CsgapC)與對照菌LA1(SNK118/pXMJ19)的種子液接種于發酵培養基中進行發酵,考察CsgapC的重組表達對谷氨酸棒桿菌產L-精氨酸的影響。結果如圖4所示,發酵過程中菌濃并無明顯差異。在40 h時重組菌LA4的L-精氨酸濃度超過對照菌LA1;最終發酵至70 h,LA4的L-精氨酸產量為11.55 g/L,較LA1(9.17 g/L)對照菌株L-精氨酸產量提高了26%,糖酸轉化率提高了10%(見表4),說明CsgapC的表達能有效提高谷氨酸棒桿菌產L-精氨酸的能力。同時,通過測定對數生長期時胞內NADPH/NADP+參數,發現重組菌LA4中的NADPH/NADP+比值為0.31,顯著高于對照菌LA1(0.21),說明重組菌LA4的胞內NADPH水平得到了提高。KABUS等[22]和ZHAN等[12]在谷氨酸棒桿菌中異源表達煙酰胺核苷酸轉氫酶PntAB催化NADP+生成NADPH,分別提高了L-賴氨酸和L-精氨酸產量。YAMAMOTO等[23]在谷氨酸棒桿菌中過表達GAPDH編碼基因,增加了從3-磷酸甘油醛到1,3-二磷酸甘油酸代謝流的流量,從而提高了L-丙氨酸產量。因此,L-精氨酸積累量的提高可能是糖酵解途徑代謝流的增加和胞內NADPH水平的提高共同作用的結果。
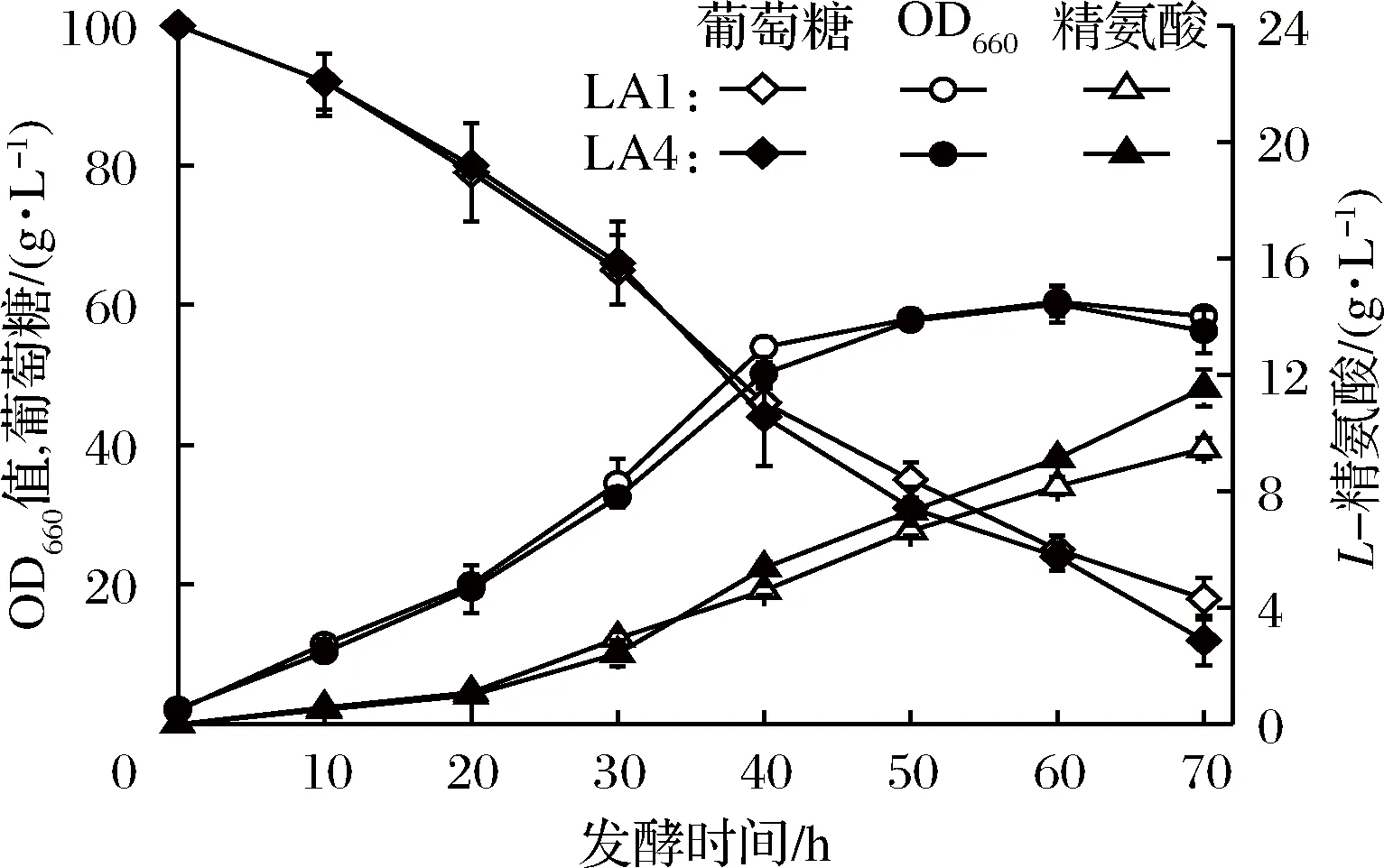
圖4 對照菌LA1(SNK118/pXMJ19)和重組菌LA4(SNK118/pXMJ19-CsgapC)發酵產L-精氨酸的比較
Fig.4 Fermentative production of L-arginine by SNK118/ pXMJ19 and SNK118/pXMJ19-CsgapC
2.3 構建L-鳥氨酸生產菌株SNK118ΔargFΔargR
L-鳥氨酸是L-精氨酸合成的中間代謝產物。C.glutamicumSNK118可用作合成L-鳥氨酸代謝改造的出發菌株。將菌落PCR驗證成功敲除argF和argR的C.glutamicumSNK118ΔargFΔargR進行L-鳥氨酸搖瓶發酵,結果如圖5所示。經過68 h的發酵,菌株LO1(SNK118ΔargFΔargR)的L-鳥氨酸產量達到22.3 g/L,L-精氨酸產量為0.14 g/L。對照菌株LR1(SNK118ΔargR)L-鳥氨酸產量僅為0.23 g/L,而L-精氨酸積累了8.62 g/L。說明敲除編碼鳥氨酸氨基甲酰基轉移酶的基因argF可以阻斷L-鳥氨酸降解為L-瓜氨酸和L-精氨酸,從而有效積累L-鳥氨酸。此外,argF與精氨酸操縱子阻遏蛋白基因argR在基因組上毗鄰,可同時被敲除,以解除反饋抑制。
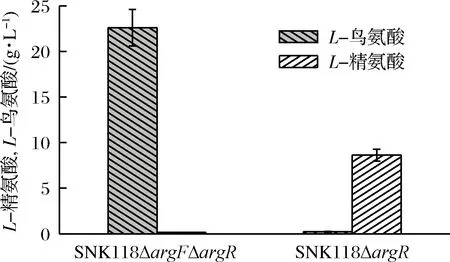
圖5 SNK118ΔargR與SNK118ΔargFΔargR的L-鳥氨酸和L-精氨酸產量的比較
Fig.5 Production of L-arginine and L-ornithine by SNK118ΔargR and SNK118ΔargFΔargR
2.4 重組菌發酵產L-鳥氨酸性能評價
NADPH的供給被認為是L-精氨酸、L-瓜氨酸和L-鳥氨酸等氨基酸合成的至關重要因素[9]。GAPDH是糖酵解途徑中的關鍵酶,其催化的反應是糖酵解途徑的限速步驟[11]。因此,為了評估CsgapC的表達對發酵生產L-鳥氨酸的影響,構建了重組菌株LO3(SNK118ΔargFΔargR/pXMJ19-CsgapC)和空白對照菌株LO2(SNK118ΔargFΔargR/pXMJ19)。在L-鳥氨酸發酵培養基中比較了菌株LO2和LO3的發酵性能。結果如圖6所示,發酵30 h后重組菌LO3的L-鳥氨酸產量高于對照菌LO2。發酵70 h后對照菌株LO2的L-鳥氨酸產量為23.11 g/L,糖酸轉化率為0.237 g/g葡萄糖。LO3的L-鳥氨酸產量為27.76 g/L,糖酸轉化率為0.274 g/g葡萄糖(表4),較對照菌株LO2分別提高了20.1%和15.6%。在細胞生長方面,菌株LO2和LO3相差不大,最終OD660均達到41。同時重組菌LO3胞內NADPH/NADP+比值較對照菌株LO2提高了60%。說明重組表達NADP+依賴型的GAPDH基因CsgapC可提高輔酶NADPH的含量,增強糖酵解途徑的代謝流量,從而提高重組谷氨酸棒桿菌的L-鳥氨酸產量。
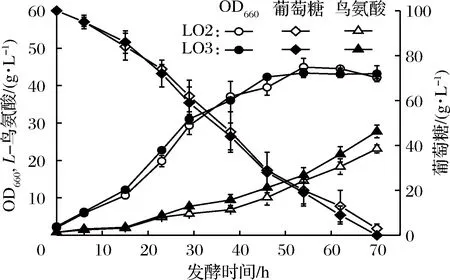
圖6 菌株LO2(SNK118ΔargFΔargR/pXMJ19)和LO3(SNK118ΔargFΔargR/pXMJ19-CsgapC)發酵產L-鳥氨酸的比較
Fig.6 Fermentative production of L-ornithine by SNK118 ΔargFΔargR/pXMJ19 and SNK118ΔargFΔargR/ pXMJ19-CsgapC
在L-鳥氨酸生產菌SNK118ΔargFΔargR中,L-鳥氨酸產量可達22.3 g/L(168.7 mmol/L),在L-精氨酸生產菌SNK118ΔargR中,L-精氨酸產量為8.62 g/L(49.5 mmol/L),可以推斷在C.glutamicumSNK118中,L-精氨酸合成途徑中由ArgF、ArgG、和ArgH組成的代謝途徑是限制步驟,后續研究在以上的基礎上圍繞提高ArgG和ArgH的酶活力來提高L-精氨酸的產量。

表4 谷氨酸棒狀桿菌生產L-精氨酸、L-鳥氨酸的發酵參數
3 結論
本文以C.glutamicumSNK118為出發菌株,考察了糖丁基梭菌來源的NADP+依賴型的GAPDH基因CsgapC的表達對發酵產L-精氨酸和L-鳥氨酸的影響。重組菌SNK118/pXMJ19-CsgapC的L-精氨酸產量比對照菌株SNK118/pXMJ19提高了26%,糖酸轉化率提高了10%。采用CRISPR-Cpf1基因編輯工具成功敲除了SNK118的基因argF和argR,構建了L-鳥氨酸生產菌株SNK118ΔargFΔargR,有效實現了L-鳥氨酸的積累,產量可達22.3 g/L。進一步將糖丁基梭菌來源的CsgapC引入SNK118ΔargFΔargR得到重組菌LO3,評估了CsgapC的重組表達對谷氨酸棒桿菌產L-鳥氨酸的影響。與對照菌相比,重組菌LO3的L-鳥氨酸產量為27.76 g/L,糖酸轉化率為0.274 g/g,較對照菌株LO2分別提高了20.1%和15.6%。說明重組表達梭菌來源的NADP+依賴型的GAPDH編碼基因可增強糖酵解途徑的代謝流量,并提高胞內NADPH的含量,從而提高L-精氨酸和L-鳥氨酸的產量。本文首次報道了通過異源表達糖丁基梭菌來源的CsgapC改造策略實現谷氨酸棒狀桿菌更高效合成L-精氨酸及L-鳥氨酸,也為提高其他氨基酸產量提供了依據。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
