大果衛矛和歐洲衛矛葉片呈色機制研究
2019-08-23 08:09:54丁彥芬王亞楠
河南農業科學 2019年8期
關鍵詞:植物
宋 鵬,丁彥芬,李 涵,王亞楠
(南京林業大學 風景園林學院,江蘇 南京 210000)
色葉植物是園林植物的重要組成部分。隨著園林綠化事業的迅速發展,人們對園林植物的色彩要求越來越高,傳統的色葉植物已經不能滿足人們的需求,因此,加強色葉植物的開發意義重大。衛矛屬植物是我國大部分地區都有分布的鄉土樹種,萌發力強,秋季葉色艷麗,對不利環境具有較強的抗性,是一類具有很高觀賞價值的園林綠化樹種[1]。但目前僅有衛矛(Euonymusalatus)、絲棉木(E.maackii)、扶芳藤(E.fortunei)、膠州衛矛(E.kiautschovicus)和冬青衛矛(E.japonicus)等[2-3]植物在園林綠地中有所應用,該屬大部分植物還處于野生狀態。
大果衛矛(E.myrianthus)是我國特有的植物,根系發達,耐修剪,葉色常綠,可用作整形植株材料。歐洲衛矛(E.europaea)原產歐洲,抗性較強,秋葉鮮紅,是一種值得推廣的優良觀葉植物,可用來制作綠籬。以往對該屬植物的研究主要集中在繁殖技術[4]上,有關該屬植物葉片結構和轉色期葉片生理變化的研究較少,鑒于此,以常綠植物大果衛矛和落葉植物歐洲衛矛為材料,觀測兩者葉片的形態結構,并對轉色期葉片的生理指標進行對比,綜合分析植物葉片的呈色機制,以期為提高該屬植物的觀賞價值和色葉植物的開發應用提供依據。
1 材料和方法
1.1 試驗地點和材料
試驗地點為江蘇省句容市乾景園林苗木有限公司基地(北緯32°05′,東經118°49′),位于長江中下游平原,屬亞熱帶季風氣候,年平均氣溫15.2 ℃,無霜期229 d,年降雨量1 058.8 mm。材料為生長健康、長勢一致的大果衛矛和歐洲衛矛3年生實生苗。
1.2 方法
1.2.1 葉片的采集 從2018年10月下旬歐洲衛矛葉色變化開始,選取生長健康、長勢一致、無明顯病蟲害的植株15株,每5株為一重復,重復3次。每8 d采一次樣,直至12月初全部葉落。每次采樣在10:00進行。選取樹冠向陽面中上部葉位相同、大小一致、健康成熟的葉片。按此法在相同時間采取大果衛矛的葉片。采后用冰盒帶回實驗室,洗凈干燥,剪去中脈,稱量完畢后置于-80 ℃冰箱保存待測。此外,選取完全變紅的歐洲衛矛和常綠的大果衛矛葉片,剪取若干 0.5 cm×0.5 cm的小塊,經 2.5%戊二醛液(北京中鏡科儀技術有限公司,分析純)固定帶回實驗室。
1.2.2 指標測定和方法 戊二醛液固定的葉片經乙醇(南京化學試劑股份有限公司,分析純)逐級脫水、臨界點干燥儀(EMITECH-K850,英國Quorum公司)干燥、離子濺射儀(HITACH E-1010,日本松中實業有限公司)真空噴鍍金膜,使用環境掃描電鏡(FEI Quanta200,荷蘭FEI公司)觀察葉片表皮和切面并拍照。
光合色素含量的測定參照張憲政[5]的方法;花色素苷相對含量的測定采用鹽酸甲醇浸提法,以每克鮮質量在10 mL提取液中變化1個OD值為1個花色素苷單位;可溶性糖和淀粉含量的測定采用蒽酮比色法[6];苯丙氨酸解氨酶(PAL)和查爾酮異構酶(CHI)活性的測定參照姜琳[7]的方法;過氧化物酶(POD)和多酚氧化酶(PPO)活性的測定參照卓啟苗等[8]的方法,以每克鮮質量在相應波長下變化0.01個OD值為1個酶活性單位。所有指標的測定均為3次重復,取平均值。
1.3 數據分析
采用Excel 2007 和 SPSS 17.0軟件對數據進行統計分析,并使用 Excel 2007作圖,折線圖中的值以平均值±標準差表示,采用Photoshop對電鏡圖進行處理。
2 結果與分析
2.1 2種植物葉片形態結構
由圖1可知,兩者的表皮形態特征相似,表皮細胞相互之間排列緊密,未發現有明顯變異的細胞,也未發現皮毛之類的附屬物。兩者的氣孔均位于下表皮,上表皮未發現氣孔。這樣的分布特點符合陸生植物的氣孔分布,既避免了干旱脅迫下水分的散失,又保證了植物光合作用的進行[9]。2種植物葉片橫切面形態結構基本相同,均是典型的異面葉,葉片組織結構從上往下,依次為上表皮、柵欄組織、海綿組織和下表皮。上表皮為單層細胞,排列緊密,細胞呈規則的方形。柵欄組織由數層規則的長柱形薄壁細胞組成,其長軸與葉片表皮垂直,相鄰細胞之間排列緊密。靠近下表皮的是海綿組織,海綿組織細胞的大小和形狀不規則。下表皮也由單層細胞構成,排列緊密。2種植物的柵欄組織細胞和上表皮之間排列緊密規整,未發現氣室。

A、B、C分別為大果衛矛葉片的上表皮、下表皮和橫切面;D、E、F分別為歐洲衛矛葉片的上表皮、下表皮和橫切面;S:氣孔;U:上表皮;PT:柵欄組織;ST:海綿組織;L:下表皮A,B,and C are the upper epidermis,lower epidermis,and cross-section of the leaves of E.myrianthus respectively;D,E,and F are the upper epidermis,lower epidermis,and cross-section of the leaves of E.europaea respectively; S:Stomata; U:Upper epidermis;PT:Palisade tissue;ST:Spongy tissue;L:Lower epidermis圖1 大果衛矛和歐洲衛矛葉片掃描電鏡照片(比例尺=200 μm)Fig.1 The scanning electron micrograph of the leaves of E.myrianthus and E.europaea(Bar=200 μm)
2.2 2種植物葉片光合色素含量和花色素苷相對含量變化
高等植物光合作用過程中吸收光能的色素統稱為光合色素,主要有葉綠素和類胡蘿卜素,其中葉綠素主要有葉綠素a和葉綠素b[10]。大果衛矛和歐洲衛矛的光合色素含量隨時間推移均呈下降趨勢,但不同色素下降幅度不一樣。12月3日大果衛矛的葉綠素含量比10月24日下降47%,葉綠素a含量的下降趨勢與葉綠素含量的下降趨勢高度一致,而葉綠素b含量下降幅度較小,據此可認為葉綠素含量的下降主要是因為葉綠素a的分解,大果衛矛花色素苷相對含量在整個時期變化不大(圖2A)。歐洲衛矛葉綠素含量12月3日比10月24日下降82%,類胡蘿卜素含量下降76%,下降的幅度明顯高于大果衛矛,說明歐洲衛矛作為落葉植物,其光合色素比常綠植物更容易分解。歐洲衛矛的花色素苷相對含量在前期上升幅度較小,但后期呈明顯上升趨勢,且高于大果衛矛的花色素苷相對含量(圖2B)。

A.大果衛矛; B.歐洲衛矛 A.E.myrianthus; B.E.europaea圖2 2種植物葉片光合色素含量和花色素苷相對含量變化Fig.2 The changes of photosynthetic pigment content and anthocyanin relative content in leaves of two Euonymus species
2.3 2種植物葉片色素含量占比變化
由于花色素苷相對含量的單位(U/g)與光合色素的單位(mg/g) 不同,故將花色素苷相對含量縮小10倍,再計算各類色素在色素總量中的占比,繪制成百分比含量堆積柱形圖(圖3)。在整個時期大果衛矛的葉綠素含量占比顯著高于花色素苷和類胡蘿卜素含量,因而葉片一直呈現綠色(圖3A)。歐洲衛矛的葉綠素含量占比由最初的65%下降到16%,類胡蘿卜素含量占比由12%下降到2%,花色素苷含量占比由最初的24%上升到83%,花色素苷含量占比升高是葉片由綠轉紅的主要原因(圖3B)。
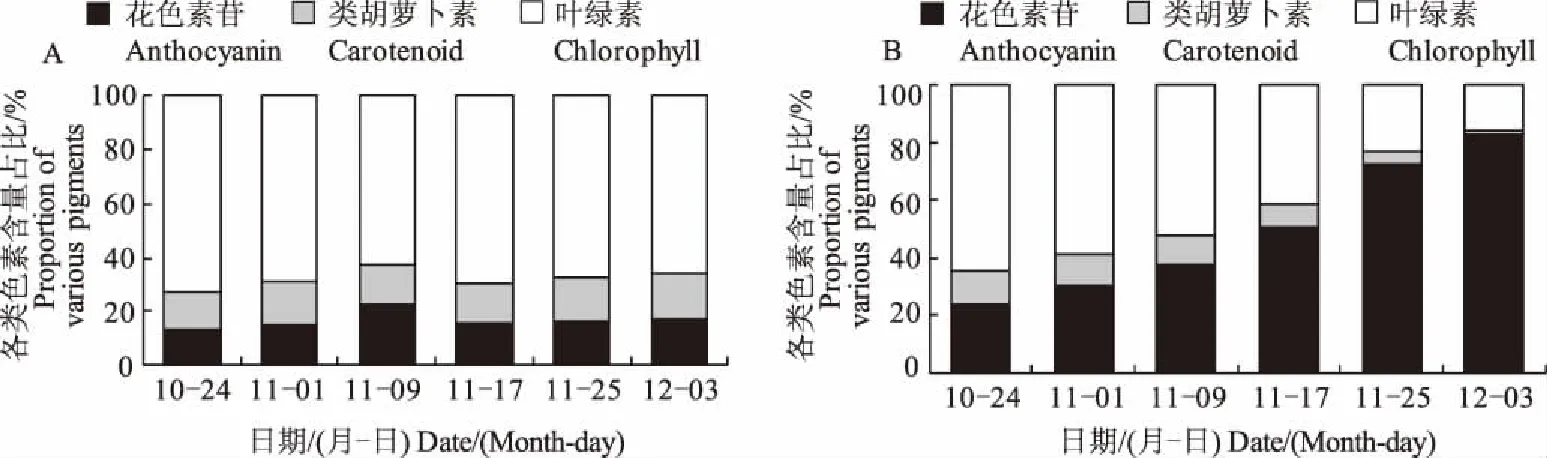
A.大果衛矛; B.歐洲衛矛 A.E.myrianthus; B.E.europaea圖3 2種植物葉片色素含量占比變化Fig.3 The changes of proportion of various pigments in leaves of two Euonymus species
2.4 2種植物葉片可溶性糖和淀粉含量變化
如圖4所示,大果衛矛和歐洲衛矛葉片可溶性糖含量變化趨勢不一致。大果衛矛的可溶性糖含量變化不大,在70 mg/g左右。歐洲衛矛的可溶性糖含量呈先上升后下降的單峰曲線,在11月1日達到峰值(97 mg/g),且大部分時期都高于大果衛矛的可溶性糖含量,結合兩者葉色變化情況,初步推測可溶性糖含量可能與葉色有一定聯系(圖4A)。兩者淀粉含量變化沒有呈現出一定規律(圖4B),這表明淀粉含量可能與葉色變化沒有關系,但大果衛矛可能通過分解淀粉以獲取更多的能量來保持葉色常綠。

圖4 2種植物葉片可溶性糖和淀粉含量變化Fig.4 The changes of soluble sugar and starch contents in leaves of two Euonymus species
2.5 2種植物葉片花色素苷相關酶活性變化
如圖5A和B所示,大果衛矛葉片的PAL和CHI活性保持平穩;歐洲衛矛葉片的PAL活性變化沒有呈現一定規律,CHI活性呈現先上升后下降的單峰曲線,峰值在11月9日出現,為346 U/g。歐洲衛矛的PAL和CHI活性顯著高于大果衛矛,表明這2種酶對花色素苷的合成可能有促進作用。大果衛矛葉片的POD和PPO活性在整個時期呈下降趨勢,但總體變化不大(圖5C、D)。歐洲衛矛的POD活性呈下降趨勢,到后期已經沒有活性,PPO活性在前期有所升高,后期下降。大果衛矛POD和PPO活性高于歐洲衛矛,表明這2種酶可能會抑制花色素苷的合成。
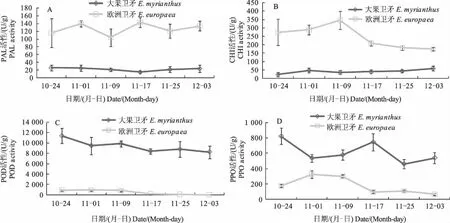
圖5 2種植物葉片花色素苷相關酶活性變化Fig.5 The changes of anthocyanin-related enzyme activities of leaves of two Euonymus species
2.6 2種植物各生理指標間的相關性分析
由表1可知,大果衛矛葉片的葉綠素和類胡蘿卜素含量呈顯著正相關,說明秋冬季常綠植物葉片的光合色素會同時發生分解。淀粉含量與POD活性呈顯著正相關,與PPO活性呈極顯著正相關。CHI活性與淀粉含量呈極顯著負相關,與POD活性呈顯著負相關。
由表2可知,歐洲衛矛葉片的葉綠素與類胡蘿卜素含量和POD活性呈極顯著正相關。CHI活性與POD和PPO活性呈顯著正相關。

表1 大果衛矛葉片各生理指標之間的相關性Tab.1 Correlation coefficients between indexes of E.myrianthus
注:*和**分別表示相關系數達到0.05和0.01的顯著水平,下同。
Note:* and **denote that correlation coefficients are significant at 0.05 and 0.01 levels,respectively.The same below.
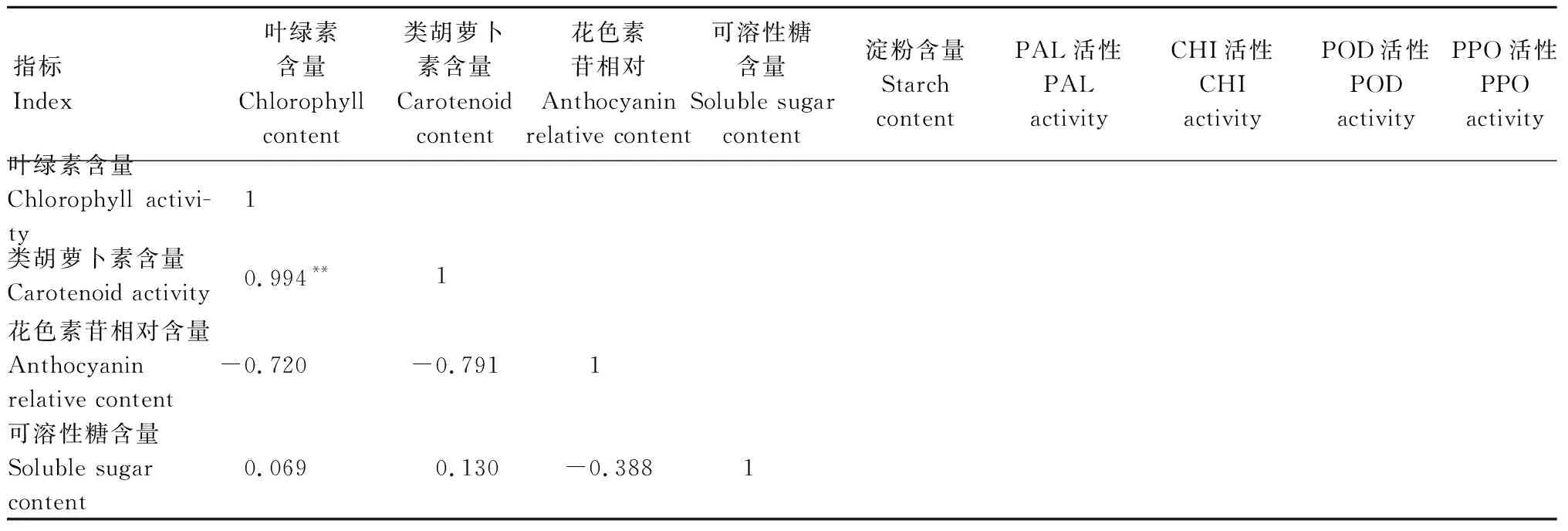
表2 歐洲衛矛葉片各生理指標之間的相關性Tab.2 Correlation coefficients between indexes of E.europaea
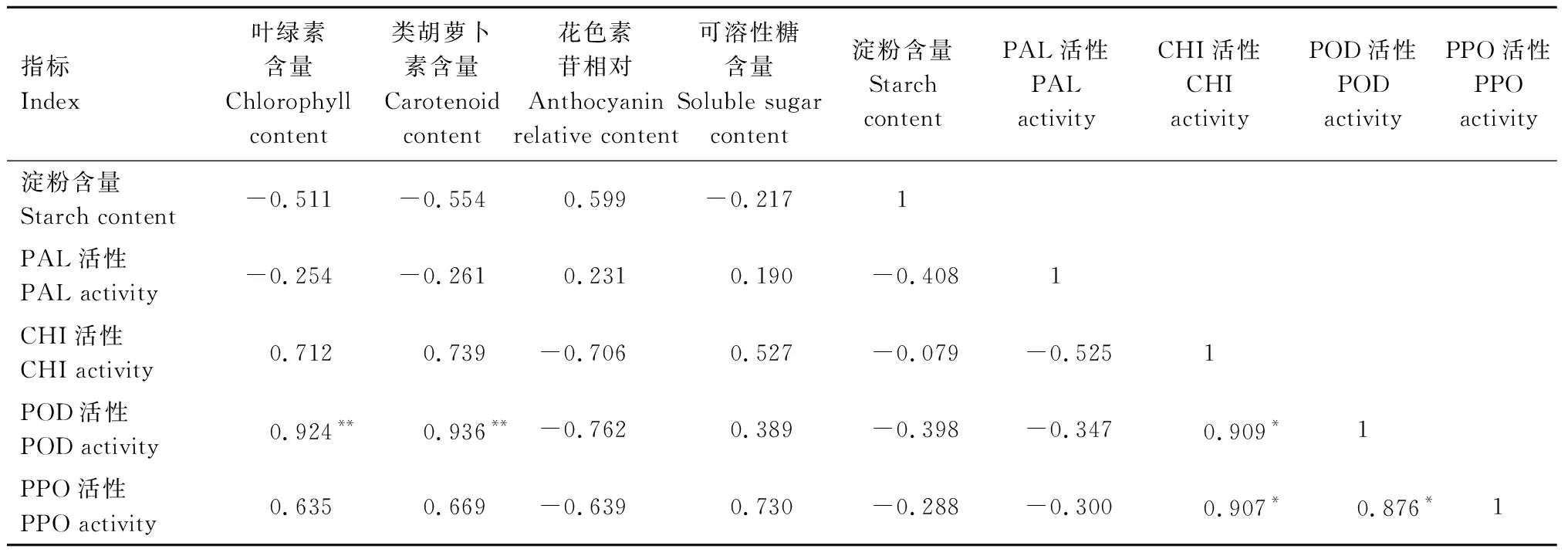
續表2 歐洲衛矛葉片各生理指標之間的相關性Tab.2(Continued) Correlation coefficients between indexes of E.europaea
3 結論與討論
植物葉片的表皮細胞和柵欄組織細胞形狀、排列以及氣室分布會影響葉片的呈色[10],這些因葉片結構發生變化而使葉色發生變化的植物被稱為結構類彩葉。狗棗獼猴桃的白色葉片的柵欄組織中比正常綠葉存在更多的氣室[11],使葉色看上去發白。金魚草花色的深淺是由花冠裂片表皮是否有圓錐狀突起的細胞決定的[12]。本試驗結果表明,通過掃描電鏡觀察,大果衛矛和歐洲衛矛的葉片均是異面葉,葉片表皮不存在圓錐形細胞,相鄰柵欄組織細胞排列緊密,不存在氣室,可以認定這2種植物葉片的結構對其呈色沒有影響。結合前人的研究,可推測結構類彩葉主要存在于陰生植物中,這類植物的葉肉組織中沒有明顯的柵欄組織和海綿組織的分化,細胞之間存在氣室,入射光在葉肉細胞氣室間會形成漫反射[10],從而使肉眼看到的葉色發生改變。
植物葉色主要是由葉片細胞中色素的種類、分布、含量以及各類色素含量的比例決定的[13]。植物葉片細胞的色素主要有4類,分別是葉綠素、類胡蘿卜素、花色素苷和甜菜色素。葉綠素合成的最低溫度為2~4 ℃,早春低溫限制了葉綠素的合成,秋冬季的低溫又加速了葉綠素的分解,這兩段時間花色素苷與類胡蘿卜素含量較多,所以植物的葉片呈現出非綠色。花色素苷可以調控葉片對光能的吸收,減少強光對葉片的破壞。甜菜色素主要分布于火龍果、雁來紅和紫茉莉等石竹目的10個科的植物中[14]。本試驗發現,大果衛矛葉片的葉綠素雖然分解了一部分,但在葉片中仍占據絕對優勢,所以葉片仍為綠色,而歐洲衛矛的葉片在初期葉綠素占比較高,葉片呈現綠色,但隨著花色素苷的不斷合成、葉綠素和類胡蘿卜素的不斷分解,花色素苷占比不斷升高,葉色從綠轉紅。
研究表明,可溶性糖的積累有助于花色素苷的合成[15-16]。對元寶楓葉片噴施質量分數4%的蔗糖溶液可以提高其觀賞性[17]。4種槭屬植物葉片的可溶性糖含量在轉色期表現為先上升后下降的單峰曲線[18],本試驗結果與之有些類似,大果衛矛的可溶性糖含量總體變化不大,歐洲衛矛的可溶性糖含量呈先上升后下降的單峰曲線,對比兩者發現,可溶性糖參與了花色素苷的合成,但是可溶性糖與花色素苷的合成并不是簡單的線性關系,因為可溶性糖不僅為花色素苷的合成提供能源,同時也為其他色素合成提供底物和能源,能在一定范圍內影響著色,但并不是決定性因素[19]。淀粉是葉片光合作用產物,主要為植物代謝提供能量,由相關性分析可知,淀粉與花色素苷的合成無顯著關系。
PAL和CHI參與花色素苷的合成,POD和PPO參與花色素苷的降解[20]。本試驗中,大果衛矛和歐洲衛矛葉片PAL和CHI活性與花色素苷相對含量呈不顯著相關,這表明PAL和CHI并不是花色素苷合成的關鍵酶。此外,大果衛矛POD和PPO活性高于歐洲衛矛,這表明POD和PPO在一定程度上會抑制花色素苷的合成[21]。花色素苷是黃酮類化合物,具有清除自由基的功能[22],與POD在植物體內共同起抗氧化的作用,兩者可能存在某種互補關系[23]。
綜上所述,大果衛矛和歐洲衛矛葉色的變化主要是因為葉片色素含量的比例發生了變化,與葉片結構沒有關系。可溶性糖促進花色素苷的合成,POD和PPO抑制花色素苷的合成。在實際環境中可以通過對葉片噴施糖溶液和提高相關酶活性來提高觀賞價值。但要更深一步地揭示植物呈色機制,可以探究植物葉片色素的合成途徑[24],也可以探索外界環境對葉片呈色的影響[25]以及葉片細胞的超微結構在轉色期的變化[26]。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
