湖南櫟類天然次生林幼樹更新特征及影響因子
2019-08-20 09:28:04朱光玉
生態學報 2019年13期
黃 朗,朱光玉,3,*,康 立,胡 松,劉 卓,盧 侃
1 中南林業科技大學林學院,長沙 410004 2 貴州省植物園,貴陽 550004 3 南方森林資源經營與監測國家林業局重點實驗室,長沙 410004
森林天然更新是森林生態系統自我繁衍的恢復手段,對未來森林群落結構的變化、功能的實現及生物多樣性的豐富具有重要影響[1]。森林天然更新主要受到環境因子(地形因子[2]、土壤因子[3]、枯落物[4]等)與林分因子(林分年齡[1]、林分密度[5]、林分郁閉度[6]、灌草蓋度[7]等)等生境因子的影響。康冰等[8]研究表明林分密度、海拔、坡度是影響秦嶺山地銳齒櫟次生林幼苗更新的主導因素。O′Brien等[9]認為林下灌木蓋度對幼齡植株的更新有阻礙作用,而林冠層蓋度卻對幼苗更新產生促進作用。李霄峰等[10]研究發現林分枯落物厚度的增加降低了林下幼苗的存活率。可以看出,影響林分天然更新的因子眾多,森林類型或更新樹種不同,影響林分天然更新的關鍵因子也不同[11-12],同時對于相同的林分其立地的差異也會導致其更新不一致[13-14]。有研究發現植株幼齡階段比成年個體更容易受生境因子影響,是決定天然更新成功與否的關鍵階段[15-16]。因此,了解林分幼齡植株的生長現狀,研究各影響因子與林分幼齡植株更新之間的關系,探究影響幼齡植株更新的關鍵因子、其不同樹種結構和復雜立地對天然次生林幼樹更新的影響,具有重要的理論與實際意義。
櫟類(Quercusspp.)面積占中國天然林總面積的14%,是我國天然林的主要組成部分,其在涵養水源、保持水土以及維持生態穩定等方面發揮了重要作用,具有很高的研究價值。櫟類林是湖南的一種典型林分類型,以中幼齡林為主。近十幾年來,由于對湖南櫟類天然林的更新特征、更新機制缺乏研究,經營方式不合理或不經營,大部分湖南櫟類天然林更新狀況不佳,部分櫟類次生林甚至有退化的趨勢。湖南是亞熱帶櫟類天然次生林的典型分布區,亞熱帶櫟類天然次生林,樹種組成結構與所處立地條件不盡相同,影響林分天然更新的關鍵因子也有差異。因此,本研究以湖南典型櫟類天然次生林為研究對象,進行標準地調查,采用k-means聚類分析,依據各樣地喬木優勢樹種的重要值以及各樣地經緯度劃分林分類型;分析不同類型櫟類次生林樹種組成和更新特征;采用單因素方差分析,分析各林分類型幼樹更新特征差異;采用Pearson相關分析,分別林分類型探討各生境因子對幼樹更新的影響,以期為湖南櫟類天然次生林的恢復與生態經營管理提供理論依據。
1 研究區概況
湖南省(24°38′—30°08′N,108°47′—114°15′E)地處中國中南部,長江中游,東西寬667 km,南北長774 km;土地總面積約為21.18萬hm2,其中林地面積1300萬hm2,森林覆蓋率59.57%,活立木蓄積5.05億m3;海拔24—2099 m,大部分地區海拔高度在100 m至800 m之間。湖南屬大陸性中亞熱帶季風濕潤氣候,全省年平均氣溫為16—18℃之間,年日照時數為1300—1800 h,無霜期長達260—310 d;年平均降水量在1200—1700 mm之間,雨量充沛,水熱充足。土壤主要以紅壤和黃壤為主,其次為紫色土和沖積土[17]。研究區內主要植被,喬木主要由甜櫧(Castanopsiseyrei)、錐栗(Castaneahenryi)、亮葉水青岡(Faguslucida)、多脈青岡(Cyclobalanopsismultinervis)、石櫟(Lithocarpusglaber)、枹櫟(Quercusserrata)、青岡櫟(Cyclobalanopsisglauca)、樟樹(Cinnamomumbodinieri)、厚皮香(Ternstroemiagymnanthera)、鵝耳櫪(Carpinusturczaninowii)等物種組成;灌木主要由鹿角杜鵑(Rhododendronlatoucheae)、山茶(Camelliajaponica)、箬竹(Indocalamustessellatus)、細枝柃(Euryaloquaiana)、尖連蕊茶(Camelliacuspidata)等物種組成;草本層主要由蘭草(EupatoriumfortuneiTurcz)、麥冬(Ophiopogonjaponicus)、淡竹葉(Lophatherumgracile)、芒萁(Dicranopterisdichotoma)等構成。
2 研究方法
2.1 數據來源
2015年12月—2016年8月分別在湖南省平江縣蘆頭林場、桑植縣八大公山自然保護區、益陽市龍虎山林場、郴州市五蓋山林場、寧鄉市青羊湖林場5個地方進行外業調查,運用典型抽樣技術在具有代表性的典型地段設立樣地進行調查。設置了20 m×30 m的以櫟類為主要優勢群落的固定樣地51塊,采用十分法計算各樣地樹種組成,其櫟類樹種組成系數的最小值為5、最大值為10、平均值為6.5、標準差為1.7,樣地均為櫟類次生林。對樣地內胸徑5 cm以上的喬木進行每木檢尺,記錄樹種、樹高、胸徑、冠幅、郁閉度及坐標等;依據每木檢尺結果,每個樣地選取3株林分平均木,采用生長錐鉆取其髓心,將3株林分平均木年齡的均值記作林分年齡;利用GPS測定樣地的地理坐標和海拔,同時記錄坡位、坡向、坡度、土壤厚度、枯落物厚度、腐殖質厚度、灌木覆蓋度及草本覆蓋度等基本情況。記錄樣地內所有幼樹(闊葉樹種:1 m≤H≤3 m;針葉樹種:0.3 m≤H≤3 m)[18]種名、地徑、高、平均冠幅以及坐標。
2.2 數據處理
采用Excel 2013對實驗數據進行處理,計算各樣地喬木、幼樹重要值以及反應幼樹更新狀況的4個指標:幼樹密度、幼樹平均地徑、幼樹平均高以及幼樹平均冠幅[16,19];根據51個樣地各主要喬木樹種重要值以及各樣地經緯度,利用R語言對其進行k-means聚類分析,劃分林分類型;采用SPSS 23.0軟件對數據進行統計分析,采用單因素方差分析法(ANOVA)檢驗不同林分類型幼樹更新指標的差異(a =0.05);運用 Pearson相關分析法對不同林分類型幼樹更新指標與環境因子以及林分因子進行相關分析。
喬木物種重要值=[相對高度(%) +相對密度(%) +相對顯著度(%)]/3[20];
幼樹物種重要值=[相對頻度(%) +相對密度(%) +相對蓋度(%)]/3[8]重要值取值范圍在 0—100%。
3 結果與分析
3.1 林分類型劃分
本研究采用k-means聚類分析法,根據51塊樣地上的喬木優勢樹種的重要值以及各樣地經緯度在R軟件上對各樣地林分進行類型劃分。本文聚類分析的分類數標準為精度≥0.95,即合并后的因子水平信息要包含合并前的因子水平信息的95%,合并后因子水平信息損失要<5%[21-22]。聚類分析分類數對應的精度見表1。
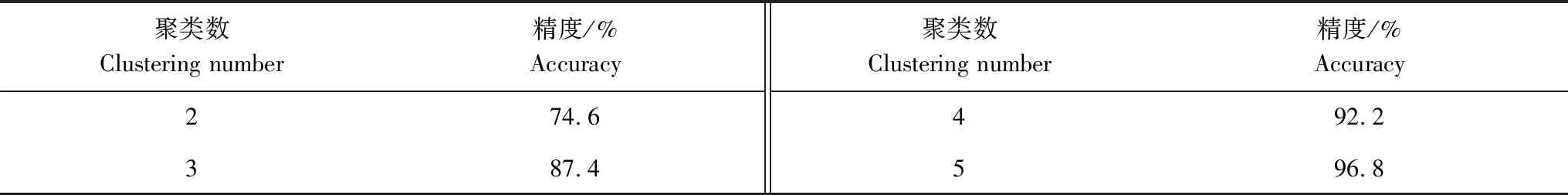
表1 聚類分類數對應精度
由聚類分析結果可知,將51塊樣地的林分聚為5類時,合并后的因子水平信息要包含合并前的因子水平信息的96.8%,滿足研究的要求。故將51塊樣地的林分共劃分為5個林分類型:甜櫧錐栗混交林(CC)、亮葉水青岡多脈青岡混交林(FC)、石櫟樟樹混交林(LC)、枹櫟甜櫧混交林(QC)、青岡櫟混交林(CG)。5個林分類型分布于湘西北(FC)、湘東北(CC)、湘中(LC、CG)以及湘南(QC)。5種林分類型樣地基本情況見表2。
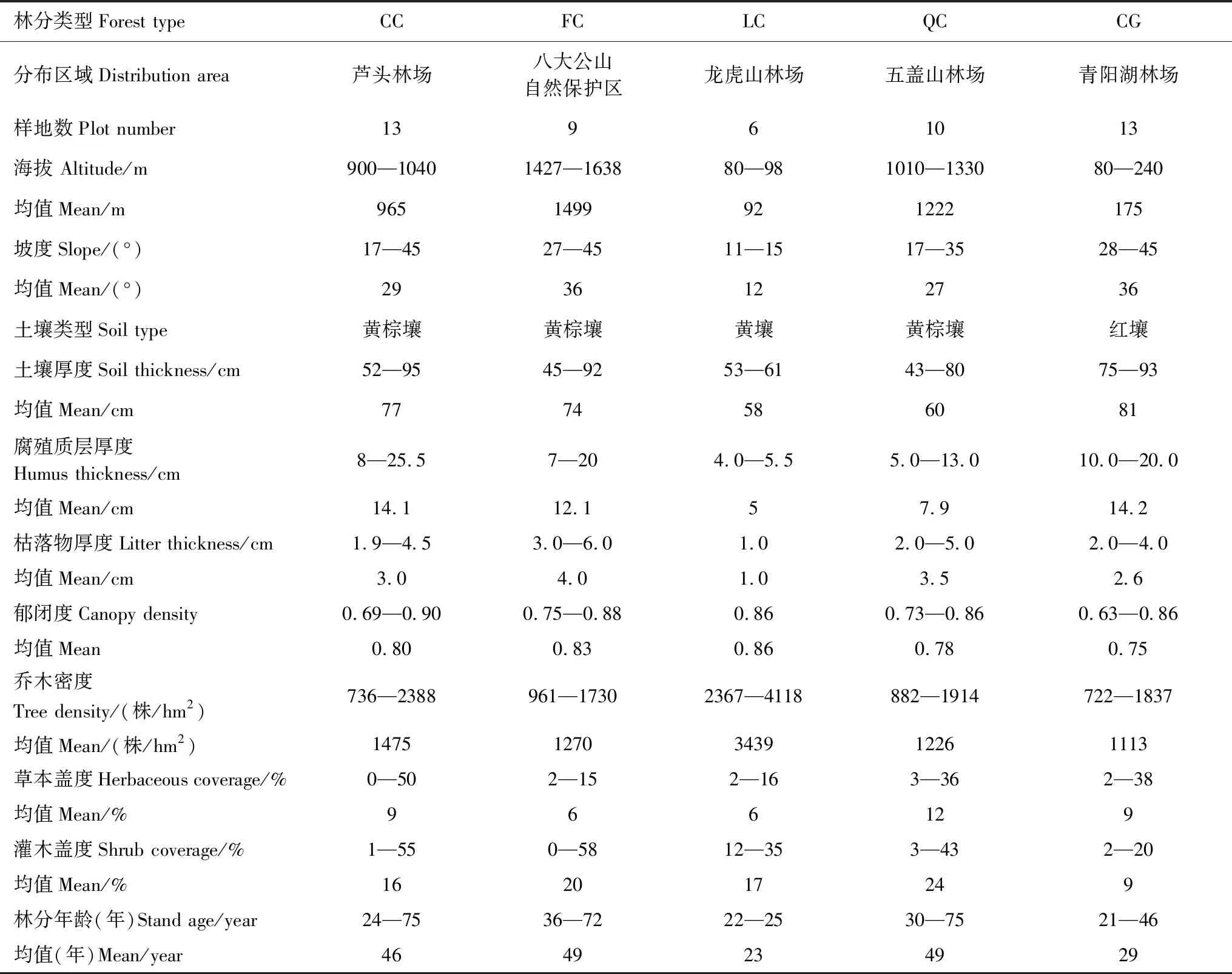
表2 5種林分類型樣地基本情況
CC:甜櫧錐栗混交林Castanopsiseyrie-Castaneahenryimixed forest;FC:亮葉水青岡多脈青岡混交林Faguslucida-Cyclobalanopsismultinervismixed forest;LC:石櫟樟樹混交林Lithocarpusglaber-Cinnamomumbodinierimixed forest;QC:枹櫟甜櫧混交林Quercusserrata-Castanopsiseyriemixed forest;CG:青岡櫟混交林Cyclobalanopsisglaucamixed forest
3.2 不同林型喬木幼樹更新特征
3.2.1不同林型喬木幼樹物種組成及特征值
根據51個樣地內更新幼樹調查數據,統計分析湖南5種不同林分類型櫟類天然混交林更新幼樹物種組成及物種特征值,見表 3。

表3 不同櫟類次生林喬木幼樹種類組成、特征值
從表3可以看出,不同類型櫟類次生林更新幼樹優勢種差異較大,物種豐富度差異顯著。CC林下更新幼樹有14種,分屬于6科11屬。優勢樹種為黃樟(Cinnamomumporrectum),重要值大于30%;其次為長葉石櫟(Lithocarpusharlandii)與甜櫧(Castanopsiseyrei),重要值均大于15%。FC林下更新幼樹有22種,分屬于12科15屬。優勢樹種為多脈青岡(Cyclobalanopsismultinervis),重要值為27.42%;其次為鵝耳櫪(Carpinusturczaninowii)、齒緣吊鐘花(Enkianthusserrulatus)和亮葉水青岡(Faguslucida),重要值均大于5%。LC林下更新幼樹有6種,分屬于3科4屬。優勢樹種為石櫟(Lithocarpusglaber(Thunb.) Nakai),在更新幼樹中占明顯優勢,重要值為60.77%;其次為杉木(Cunninghamialanceolata)和樟樹(CinnamomumbodinieriLevl.),重要值均大于10%。QC林下更新幼樹有18種,分屬于10科,16屬。優勢樹種為山礬(Symplocoscaudata)、吊鐘花(FuchsiahybridaVoss.)、黃樟和甜櫧,重要值均大于10%;其次為交讓木(DaphniphyllummacropodumMiq.)、長葉石櫟和枹櫟(QuercusserrataThunb.),重要值均大于5%。CG林下更新幼樹有6種,分屬于5科5屬。優勢樹種為青岡櫟(Cyclobalanopsisglauca),在更新幼樹中占明顯優勢,重要值為78.53%;其次為多脈青岡和杉木,重要值均大于5%。5種櫟類次生林更新幼樹的共有種為青岡櫟,出現頻率較高的樹種有杉木、山礬、細葉青岡以及長葉石櫟等。
3.2.2不同林型喬木幼樹數量特征和生長狀況
本文采用4個更新指標分別反映各林分類型幼樹數量特征(幼樹密度)和生長情況(幼樹平均地徑、平均高以及平均冠幅)。由圖1可以看出,5種不同櫟類次生林幼樹密度均未超過500株/hm2,更新情況較差。5種不同櫟類次生林幼樹密度大小順序為:亮葉水青岡多脈青岡混交林>石櫟樟樹混交林>青岡櫟混交林>枹櫟甜櫧混交林>甜櫧錐栗混交林,差異顯著。青岡櫟混交林幼樹平均地徑、平均高最大,其平均冠幅僅次于亮葉水青岡多脈青岡混交林,表明該林型幼樹生長情況較好,亮葉水青岡多脈青岡混交林次之,石櫟混交林最差。
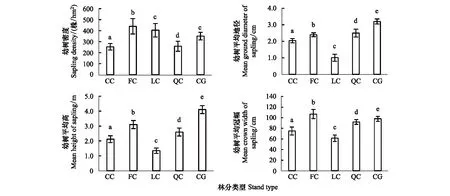
圖1 不同櫟類次生林幼樹更新指標特征(均值±標準誤)Fig.1 Features of saplings regeneration index in different Quercus secondary forests (mean±SE)柱狀圖頂部字母的不同表示不同林型間更新指標存在顯著性差異(P<0.05)
3.3 不同櫟類次生林幼樹更新影響因子相關性分析
分別以不同類型櫟類次生林的海拔(x1)、坡度(x2)、坡向(x3)、坡位 (x4)、土壤厚度(x5)、腐殖質厚度(x6)、枯落物厚度(x7)等環境因子和林分年齡(x8)、郁閉度(x9)、喬木密度(x10)、草本蓋度(x11)、灌木蓋度(x12)等林分因子為自變量,以幼樹密度(y1)、幼樹平均高(y2)、幼樹平均地徑(y3)、幼樹平均冠幅(y4)為因變量進行相關性分析,所得結果如表4。
表4結果顯示,5種林分類型中除了林分年齡與4個更新指標均無顯著關聯外,其余7個環境因子、4個林分因子與4個更新指標的部分指標有顯著相關性,但各個林分類型間與4個更新指標顯著相關的因子存在差異。CC中幼樹密度與腐殖質厚度顯著負相關,其余各環境因子對幼樹生長影響不大;幼樹平均高與灌木蓋度呈顯著正相關;幼樹平均地徑與草本蓋度、灌木蓋度呈顯著正相關。FC中幼樹密度與海拔、腐殖質厚度、枯落物厚度呈顯著正相關,與草本蓋度呈極顯著正相關;坡位對幼樹平均高、幼樹平均冠幅產生顯著正影響;幼樹平均地徑與郁閉度呈顯著負相關。LC中幼樹密度與坡向、土壤厚度呈顯著正相關,其余因子對幼樹生長無顯著影響。QC中幼樹冠幅、幼樹密度分別與坡度、坡位呈顯著負相關;幼樹密度與郁閉度、喬木密度呈極顯著正相關。CG中幼樹平均地徑與土壤厚度呈顯著正相關,與喬木密度呈極顯著正相關;幼樹平均冠幅與灌木蓋度呈顯著正相關。
4 結論與討論
4.1 林分類型劃分
櫟類林是湖南的一種典型森林類型,其在湖南的分布具有明顯的地帶性差異與地域性特點[23]。為了揭示湖南不同地區、不同類型櫟類次生林的天然更新規律,本研究采用k-means聚類分析,將51個樣地櫟類次生林劃分為5個林分類型,分別為:甜櫧錐栗混交林(CC)、亮葉水青岡多脈青岡混交林(FC)、石櫟樟樹混交林(LC)、枹櫟甜櫧混交林(QC)、青岡櫟混交林(CG)。聚類精度達到96.8%,說明聚類分析劃分的林分類型與野外調查所觀察到的類型非常一致。
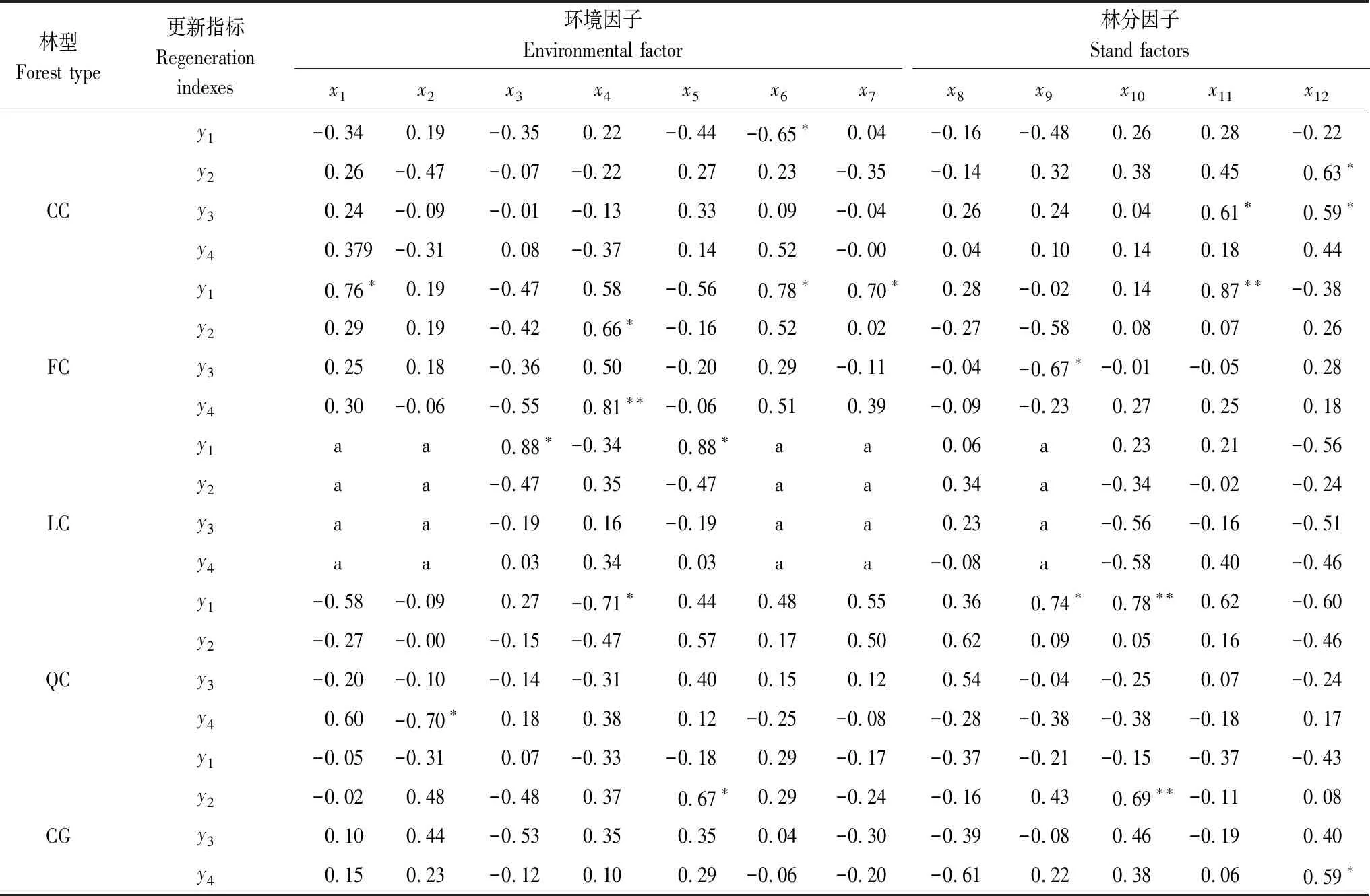
表4 不同櫟類次生林幼樹更新指標與影響因子間的相關系數
x1:海拔Altitude;x2:坡度Slope;x3坡向Slope aspect;x4坡位Slope position;x5土壤厚度Soil thickness;x6腐殖質厚度Humus thickness;x7枯落物厚度Litter thickness;x8林分年齡Stand age;x9郁閉度Canopy density;x10喬木密度Tree density;x11草本蓋度Herbaceous coverage;x12灌木蓋度Shrub coverage;y1幼樹密度Sapling density;y2幼樹平均高Mean height of sapling;y3幼樹平均地徑Mean ground diameter of sapling;y4幼樹平均冠幅Mean crown width of sapling ;*:P<0.05(雙尾);**:P<0.01(雙尾);a:由于至少有一個變量為常量,因此無法進行計算
4.2 櫟類次生林喬木幼樹物種組成及特征值
有研究表明,氣候條件是決定植物類型或生活型分布的主要因素[24]。本研究顯示,5種櫟類次生林林下更新幼樹物種豐富度與幼樹物種組成差異較大。這與康冰[25]的研究結果相似,主要是因為5種櫟類次生林空間分布區域不同,各區域氣候條件、立地因子存在差異,各林分主要建群種的種類、林分結構以及微環境不同,從而導致更新物種產生較大的分化。在石櫟樟樹混交林(LC)和青岡櫟混交林(CG)中,更新幼樹石櫟、青岡櫟占明顯優勢,重要值均占其所在林分類型的60%以上,且二者皆為喬木層優勢樹種,可能是由于,其喬木層物種組成相對簡單,主要優勢櫟類樹種占絕對優勢,其余樹種沒有與優勢種形成明顯的競爭關系。其余3種櫟類次生林,喬木層物種組成相對復雜,物種豐富度較大,更新幼樹的優勢種不明顯。尤其是枹櫟甜櫧混交林(QC),更新幼樹的優勢樹種有山礬、吊鐘花、黃樟和甜櫧,重要值均大于10%,形成了明顯的競爭關系。
4.3 櫟類次生林喬木幼樹數量特征及生長狀況
從喬木幼樹數量特征與生長狀況來看,5種不同櫟類次生林幼樹密度均未超過500株/hm2,更新情況較差;不同櫟類次生林喬木幼樹數量及生長情況差異顯著。亮葉水青岡多脈青岡混交林(FC)更新幼樹的株數密度最大,幼樹平均地徑、平均高以及平均冠幅長勢較好,說明該林分類型處于較為健康的演替階段。甜櫧錐栗混交林(CC)和枹櫟甜櫧混交林(QC)處于演替后期,更新能力不足,導致兩種櫟類次生林的幼樹更新密度較小。石櫟樟樹混交林(LC)與青岡櫟混交林(CG)處于演替初期,幼樹更新密度較大,但石櫟樟樹混交林(LC)幼樹平均地徑、平均高以及平均冠幅均明顯小于其他櫟類次生林,這主要是因為石櫟樟樹混交林(LC)的喬木株數密度過大,導致其林下更新幼樹生長空間不足,難以存活。
4.4 不同櫟類次生林幼樹更新影響因子相關分析
幼樹的生長發育受林分的光照、溫度、水分和土壤條件等因子的影響,而環境因子(地形因子、土壤因子、枯落物)、林分因子(年齡結構、密度因子、灌草蓋度)與這些生境因子密切相關,間接的影響更新幼樹的生長[26]。相關分析結果顯示,櫟類次生林幼樹更新是各個影響因子相互作用的結果,不同類型次生林幼樹更新的主要影響因子存在差異。
甜櫧錐栗混交林(CC)中幼樹密度與腐殖質厚度呈顯著負相關,說明腐殖質對該林分類型的幼樹生長有阻礙作用,這與董麗[27]的研究結果一致。幼樹平均高與灌木蓋度呈顯著正相關,幼樹平均地徑與草本蓋度、灌木蓋度呈顯著正相關。這是由于灌木蓋度與幼樹密度呈微弱的負相關,隨著灌木蓋度的增大,幼樹數量減少,更新幼樹間對有限資源的競爭減弱,有利于幼樹株高和地徑的生長。
亮葉水青岡多脈青岡混交林(FC)中幼樹密度與海拔、腐殖質層度、枯落物厚度呈顯著正相關,這是由于海拔高度的變化導致林分中光熱條件也發生了變化,隨著海拔高度的增加林分溫度降低濕度加大,土壤微生物活動受阻,有機質分解較慢,導致腐殖質、枯落物厚度增加,更適合林分中幼樹的生長,使其數量增加[28]。坡位對幼樹株高產生顯著正影響,對幼樹冠幅產生極顯著正影響。坡位對于樹木的生長的影響不是孤立的,常常和坡向、坡度等因子綜合在一起起作用[29],亮葉水青岡多脈青岡混交林所處的研究地點山坡坡度較大(平均坡度大于36°,表2),而山脊或靠近山脊的上坡相對而言比較平坦開闊,山脊的土壤較山坡土壤深厚肥沃,使得山脊的幼樹比山坡的幼樹生長的更好。幼樹平均地徑與郁閉度呈顯著負相關,這是因為高郁閉度森林的林下光照不足和光質改變,不利于林下幼樹的生長,Denslow[30]、于飛等[31]研究也證實了這一點。草本蓋度與幼樹密度呈極顯著正相關,亮葉水青岡多脈青岡混交林的草本蓋度在2%—15%之間(表2),說明在這個范圍內,草本蓋度的增加對幼樹的生長起促進作用。
石櫟樟樹混交林(LC)中幼樹密度與坡向、土壤厚度呈顯著正相關外,其余各個影響因子對幼樹更新未產生顯著影響,說明坡向、土壤厚度是影響該林分類型幼樹生長的主要因子,陽光越充足且土壤越深厚肥沃的環境條件下更適合林分中幼樹的生長。
枹櫟甜櫧混交林(QC)中幼樹密度、幼樹冠幅分別與坡位、坡度呈顯著負相關,說明林分中幼樹適合生長在地勢平坦土壤肥沃的下坡位。郁閉度與幼樹密度呈顯著正相關、喬木密度與幼樹密度呈極顯著正相關,在882—1914株/hm2范圍內(表2),隨著喬木密度的增加,林分郁閉度也隨之增加,林分中幼樹密度呈增加趨勢。這主要是因為不同林分喬木密度、郁閉度所產生的森林群落內光熱條件的異質性,直接影響到林下幼樹的生長發育[32]。
青岡櫟混交林(CG)幼樹平均地徑與土壤厚度呈顯著正相關,這是由于隨著土壤厚度的增加,土壤中有機質含量越多,土壤越肥沃,更適合林下幼樹的生長發育。喬木密度對幼樹平均地徑產生極顯著正影響,幼樹平均地徑隨喬木密度的增加而增長。由于喬木密度與幼樹密度呈微弱的負相關,隨著喬木密度的增加,幼樹數量減少,更新幼樹間對有限資源的競爭減弱,有利于更新幼樹的生長[33]。幼樹平均冠幅與灌木蓋度呈顯著正相關,說明在一定范圍內,灌木蓋度對林下幼樹的生長有促進作用。但幼樹密度與灌木蓋度呈微弱的負相關,這與甜櫧錐栗混交林(CC)中研究結果一致,說明林下幼樹與灌木之間存在著競爭,隨著灌木蓋度的增加,幼樹生長空間減少,導致幼樹數量減少。
