莧菜AtPAL基因克隆與表達分析
2019-08-15 11:36:44黃彬茹趙春麗賴鐘雄劉生財
園藝與種苗 2019年7期
曾 林 ,黃彬茹 ,王 曉 ,趙春麗 ,賴鐘雄 *,劉生財 *
(1.福建農林大學園藝學院,福建福州 350002;2.福建農林大學園藝植物生物工程研究所,福建福州 350002)
莧菜(Amaranthus tricolor L.)屬于石竹目莧科莧屬,為一年生植物,又名鳧葵、莕菜等。莧菜中富含多種營養物質,具有很高的營養價值[1],并且富含類黃酮[2]、甜菜色素[3]等多種具保健功能的次生代謝物質,在近20年來成為研究熱點,引起越來越多的關注[4]。
苯丙烷代謝通路是植物體中苯丙烷類物質代謝的一條重要途徑,其直接或間接產生了植物體中所有含苯丙烷骨架的物質,如木質素、黃酮類物質等[5-6],苯丙氨酸解氨酶(phenylalaniammonia-lyase)是苯丙烷途徑代謝中的第一種酶,也是關鍵酶和限速酶[7],是許多次級代謝產物的前體[8]。大量研究表明,PAL基因在維持植物正常生長發育及參與植物抗逆境作用等方面均發揮著重要作用[9-10],如菜心的抗逆性[11]、桂花類黃酮的積累[12]等。目前,研究PAL基因的表達調控機理對研究植物次生代謝物的產生具有重大意義,是現在研究的熱點之一。
光是植物體正常生長發育必不可少的環境因子,不僅直接影響到植物的光合作用,同時對植株形態建成、營養元素、化學成分等方面均有一定影響[13]。前人研究表明,光對PAL基因的表達以及甜菜色素、類黃酮等物質的積累也具有重要作用。謝靈玲等[14]研究發現,紅光、藍光、紫外光都能夠誘導大豆PAL基因的表達;劉洋等[15]研究發現,不同光質對青錢柳葉黃酮類化合物含量和PAL活性均有顯著影響,且兩者存在顯著的交互作用。
水楊酸(Salicylic acid,SA)是一種重要的植物激素,可調節植物體的一些生理活動,在植物體的抗病抗逆上具有重要作用[16]。前人研究發現,PAL基因參與植物體內水楊酸的合成[16]。趙亞婷[17]研究發現,水楊酸處理能增強杏果實PAL基因的表達,并且能顯著提升杏果實PAL的活性及類黃酮含量;韓秀麗[16]研究發現SA處理能夠誘導大麥PAL基因的表達。
1 材料與方法
1.1 材料
試驗所用種子材料是從南京金盛達種子有限公司購買的‘全紅’莧菜品種。選擇健康飽滿的種子經催芽后播種于營養缽中,放置于室外自然光照條件下,30 d后莧菜苗長到20 cm左右,取其葉片用于RNA提取,每次取樣均為3次重復。
1.2 總RNA的提取及cDNA的合成
莧菜總RNA提取使用Bio Teke的多糖多酚植物總RNA提取試劑盒(北京百泰克生物技術有限公司)。采用1%的瓊脂糖凝膠電泳檢測莧菜總RNA的可靠性,利用超微量分光光度計檢測RNA樣品的濃度和純度。cDNA第1鏈采用Fermentas公司的RevertAidTM First Strand cDNA Synthesis Kit試劑盒進行合成,用于 ORF 及 3′-RACE、5′-RACE 的擴增。
1.3 AtPAL基因cDNA全長序列克隆
1.3.1 引物設計與合成。從福建農林大學園藝植物生物工程研究所實驗室已構建的莧菜轉錄組數據庫中篩選PAL基因的序列片段,然后通過NCBI數據庫比對確定所要克隆的PAL基因序列,利用DNAMAN7.0軟件和NCBI在線比對分析,從開放閱讀框兩端設計1對特異引物,利用擴增出來的ORF區域,根據 3′-RACE、5′-RACE 的要求設計引物,引物由北京六合華大基因科技有限公司合成(表1)。
1.3.2 AtPAL基因ORF序列的獲得。以合成的cDNA第1鏈為模板進行PCR擴增,擴增產物利用瓊脂糖凝膠回收試劑盒(Gel/PCR Extraction Kit)進行回收純化,以pMD18-T為載體進行連接,利用大腸桿菌DH5α感受態細胞進行轉化,而后進行活化、涂板,37℃恒溫培養12 h、挑菌,37℃搖床中培養12 h,菌檢PCR,檢驗后菌液送至北京六合華大基因科技有限公司進行測序驗證。
1.3.3 AtPAL基因3′RACE和5′RACE擴增。從獲得的ORF片段靠近3′端及5′端處分別設計基因序列的2條3′-RACE正向特異引物及2條5′-RACE反向引物(表1),以逆轉錄合成的cDNA為模板,進行2輪槽式PCR擴增,擴增出3′端帶PloyA的UTR序列及5′端UTR序列。擴增后進行測序拼接驗證。

表1 引物序列
1.4 AtPAL基因的生物信息學分析
利用DNAMAN7.0進行基因序列及氨基酸序列比對;采用NCBI的BLAST檢索分析基因序列,并從GenBank中挑選出甜菜、菠菜等27個物種的PAL基因的氨基酸序列,采用MEGA7.0構建系統進化樹;采用 ProtScale(http://web.expasy.org/protscale/)進行AtPAL蛋白基本理化性質分析;采用SOPM(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopm.pl)進行蛋白二級結構分析;采用SWISS.MODEL(http://swissmode1.Expasy.org/interactive)進行蛋白三級結構預測;采用 Wolf Psort Prediction(http://www.gen—script.com/psort/wolf—psort.htm1)進行亞細胞定位分析 ;采用 TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)進行蛋白跨膜結構分析;采用SignalP(http:/www.Cbs.Dtu.dk/services/SignalP/)進行蛋白信號肽分析;采用 NetPhos(http:/myhits.isb—sib.ch/cgibin/motif—scan)進行翻譯后蛋白修飾位點分析。
1.5 非生物脅迫條件下AtPAL基因表達分析
1.5.1 不同光質處理下。選取健康飽滿的‘全紅’莧菜種子,依次用體積分數75%乙醇、0.1%升汞進行消毒處理1、8 min,消毒后用無菌水對種子進行5次沖洗,在無菌的條件下將消毒后的‘全紅’莧菜種子播種在MS培養基中,然后放置在恒溫培養室中暗培養7 d。7 d暗培養后將材料移至不同光質(紅、藍、綠、白、黑暗)下進行培養,在處理后的3 h時取莧菜幼苗的根上部分,隨機取樣,重復3次,凍存,之后進行qRT-PCR定量分析。
1.5.2 不同濃度SA處理下。根據日本園試配方配制所需的水培營養液,將‘全紅’莧菜種子放置于營養液上的水培海綿塊中全光照培養20 d(水培盆周邊進行遮光),20 d后將材料移入不同濃度SA(0.25、0.5、0.75、1.00、1.25 mmol/L)條件下進行培養,以不含SA的組為對照,處理12 h后取莧菜幼苗的根上部分,隨機取樣,重復3次,凍存,之后進行qRT-PCR定量分析。
1.5.3 AtPAL基因熒光定量PCR分析。使用Bio Teke多糖多酚植物總RNA提取試劑盒提取樣本總RNA,采用1%瓊脂糖凝膠電泳檢測總RNA的可靠性,利用超微量分光光度計檢測RNA濃度和純度,利用SYBR ExScriptTM試劑盒合成cDNA。根據AtPAL基因全長設計qRT-PCR定量表達特異引物,Actin為內參基因(表1),采用Roche Lightcycler480進行定量表達分析,計算基因相對表達量。
2 結果與分析
2.1 AtPAL基因的克隆結果
以‘全紅’莧菜為材料進行基因序列擴增獲得的結果如圖1所示,通過NCBI的比對和DNAMAN的拼接驗證,確認最終獲得了莧菜PAL基因cDNA的全長序列(GenBank登錄號:MK574874),將其命名為AtPAL,該基因cDNA全長為2 476 bp,包含一個長為2 142 bp的ORF,預測編碼713個氨基酸,5′端UTR 序列為 112 bp,3′端 UTR 序列為 222 bp,poly A尾巴長為29 bp,終止密碼子為TAA。
2.2 AtPAL基因的生物信息學
2.2.1 AtPAL蛋白基本理化性質。莧菜AtPAL蛋白基本理化性質分析結果顯示,該蛋白理論編碼蛋白分子量為(Mw)77.469 2 kD,理論等電點(pI)為6.10,分子式為C3408H5455N957O1048S27,原子總數為10 895,不穩定系數(II)為34.96,小于40,推測莧菜AtPAL蛋白為不穩定蛋白,脂肪系數(AI)為90.08。AtPAL中帶正、負電氨基酸數量分別為81個和72個。
2.2.2 AtPAL蛋白序列二級及三級結構。使用SOPM預測的莧菜AtPAL蛋白的二級結構(圖2)顯示,序列中α螺旋、β轉角、延伸片段和隨機卷曲分別占51.05%、7.57%、10.80%和30.58%,說明AtPAL蛋白二級結構以α螺旋為主。使用SWISS-MODEL預測的AtPAL蛋白三級結構如圖3所示。

圖1 莧菜AtPAL基因PCR擴增產物

圖2 莧菜AtPAL蛋白二級結構分析結果

圖3 莧菜AtPAL蛋白三級結構

圖4 27個物種 PAL蛋白系統進化樹
2.2.3 AtPAL蛋白信號肽、亞細胞定位及跨結構域。TMHMM分析結果表明,AtPAL蛋白屬于非跨膜蛋白,不含跨膜螺旋區。SignalP分析結果顯示,AtPAL蛋白不含信號肽序列。使用Wolf Psort預測表明,其亞細胞定位于內質網。NetPhos 2.0 Serve分析結果顯示,AtPAL蛋白含有59個磷酸化修飾位點,絲氨酸(S)、酪氨酸(Y)、蘇氨酸(T)磷酸化修飾位點分別為31個、8個和20個,表明AtPAL蛋白主要在絲氨酸和蘇氨酸殘基上磷酸化。
2.2.4 不同物種PAL蛋白序列對比與進化樹分析。AtPAL氨基酸序列比對結果顯示,AtPAL與甜菜、藜麥、菠菜3種甜菜色素代謝類型植物的PAL同源性均在90%以上,并且與花青素代謝類型的康乃馨、葡萄、杏、毛白楊、番茄的同源性也都在84%以上,這進一步證實該次克隆獲得的是莧菜的AtPAL基因。利用MEGA7.0構建的系統進化樹如圖4所示,結果顯示,單子葉植物與雙子葉植物被分成了2個不同的分支,莧菜AtPAL蛋白與同為石竹目的甜菜、菠菜、康乃馨、藜麥4個物種的PAL蛋白聚成一類,其中同屬藜科的甜菜、菠菜、藜麥單獨聚成一小類,表明莧菜AtPAL蛋白與藜麥、甜菜、菠菜、康乃馨的PAL蛋白親緣關系較近,而與水稻、葡萄、番茄等的PAL蛋白親緣關系較遠。
2.3 非生物脅迫條件下AtPAL基因實時定量PCR(qRT-PCR)分析
2.3.1 不同光質處理對AtPAL基因表達量的影響。AtPAL基因的定量分析結果顯示,在紅光、白光、綠光、藍光4種不同光質處理下,除白光處理外,AtPAL基因在’全紅’莧菜中的表達量相較于CK對照(黑暗)都呈現上調變化(圖5),而白光處理下則呈現下調變化。在藍光處理下,AtPAL基因表達量最高,且與對照組相比出現極顯著差異(P<0.01),但紅光、白光、綠光處理下與對照組相比AtPAL基因的表達量沒有出現顯著差異,說明藍光能夠明顯促進AtPAL基因的表達。

圖5 不同光質處理下AtPAL基因表達情況
2.3.2 不同濃度SA處理對AtPAL基因表達量的影響。AtPAL基因的定量分析結果顯示,在不同濃度SA的處理下,AtPAL基因在‘全紅’莧菜中的表達量相較于對照組都出現上調的變化(圖6),呈現出“升—降—升”的變化趨勢,且不同濃度SA處理下AtPAL基因的表達量相較于對照組都出現極顯著差異(P<0.01),當 SA 濃度為 0.75 mmol/L時,表達量達到最高。
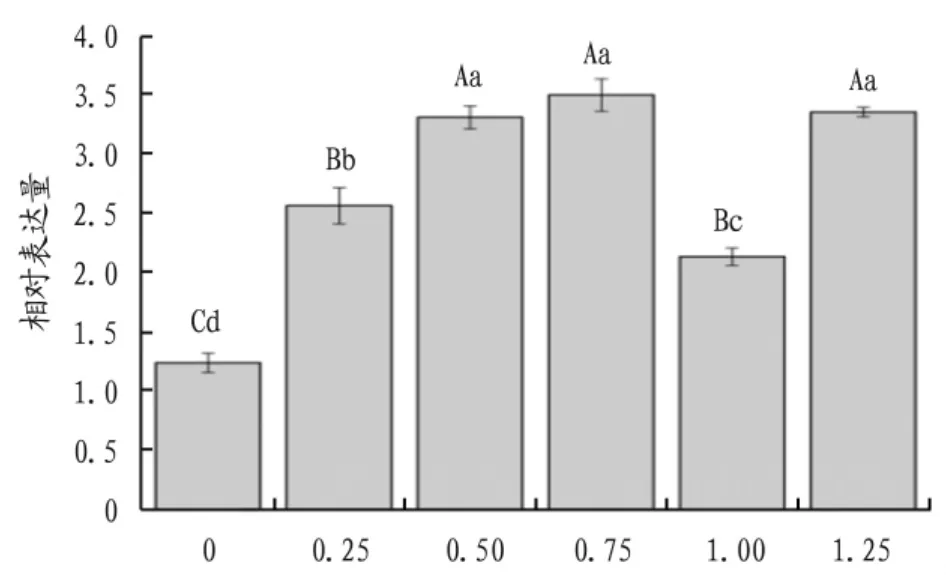
圖6 不同濃度水楊酸處理下AtPAL基因表達情況
3 討論
目前,PAL基因已在石榴、夏枯草等物種中成功克隆[18-19],前人研究表明,目前所報道物種的PAL同源基因cDNA序列長度為2 096~2 650 bp,編碼628~730個氨基酸[20],該研究克隆得到的莧菜AtPAL基因cDNA序列全長為2 476 bp,編碼713個氨基酸,與前人研究所得結果接近。
細胞內蛋白質磷酸化與信號轉導密切相關,其磷酸化部位主要有蘇氨酸、酪氨酸和絲氨酸結合位點[11],PAL磷酸化后其蛋白質活性也會改變,這種改變對于植物生理生化應答反應具有重要意義[21]。該研究發現AtPAL蛋白主要是在絲氨酸和蘇氨酸殘基上磷酸化,少數在酪氨酸位置上被磷酸化,與馮立娟等[18]研究所得結論相符。系統進化樹結果表明,AtPAL氨基酸序列與藜麥、菠菜、葡萄等植物的PAL氨基酸序列具有很高的同源性,可見PAL作為植物苯丙烷代謝途徑中第1個關鍵酶,在進化的過程中有著較強的演化趨同性與遺傳穩定性,與前人的研究結果吻合[22]。分析結果可以為生物進化研究提供一定的科學參考。
謝靈玲等[14]研究發現,紅光、藍光、紫外光都能促進大豆PAL的基因表達,該研究發現不同單色光處理,都能夠上調AtPAL基因的表達,并且藍光最有利于AtPAL基因的表達,與前人研究所得結論一致。韓秀麗[16]研究發現水楊酸可以促進大麥PAL基因的表達,在該研究中也發現水楊酸能夠上調AtPAL基因的表達,結果與前人研究結論相符。AtPAL基因在不同非生物脅迫條件下的應答反應暗示其在莧菜的抗逆防御反應過程中發揮著重要作用。此外,大量研究表明PAL基因的表達與光質、水楊酸、以及黃酮類物質的含量都存在明顯的相關性[15,23-24],謝靈玲等[14]發現,紅、藍光在促進大豆PAL基因表達的同時還能促進異黃酮的積累,趙亞婷[17]發現SA能夠顯著促進杏果實PAL基因的表達及類黃酮的積累,提升果實抗病性,因此推測莧菜很有可能是通過調節AtPAL基因的表達來控制黃酮類物質的合成從而提升其對逆境的抵抗能力,但具體是否存在這樣的分子調控機制還有待進一步的探究。
該研究首次對莧菜AtPAL基因進行克隆并進行生信分析和表達分析,為探討莧菜次生代謝的調控機理奠定基礎,也在提高植物類黃酮含量、增強作物抗逆等方面有一定的指導價值。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
