貴州白水牛ASIP、MC1R和TYR基因多態及其與白毛性狀的關聯分析
2019-08-05 01:17:26史穎超項門們汪富文李思華史芯蓓雷初朝黨瑞華
中國牛業科學 2019年3期
關鍵詞:貴州
史穎超, 項門們, 汪富文, 李思華, 史芯蓓, 雷初朝, 黨瑞華*
(西北農林科技大學動物科技學院,陜西 楊凌 712100)
水牛(Bubalus babulis)是世界牛遺傳資源的重要組成部分,其主要分布在亞洲地區且飼養數量占據該地區90%以上,根據亞洲水牛的外形特征、生活習性和生產用途將其分為沼澤型和河流型兩種類型,是熱帶和亞熱帶地區獨具特色的一個物種[1-2]。在中國,水牛在數量上僅次于黃牛,排名第二。作為貴州所特有的水牛品種,貴州白水牛主要產自黔北地區,具有被毛顏色獨特、抗病力強、役用性能好、屠宰性能高、繁殖性能良好等優點。據《貴州省畜禽品種志》記載貴州的水牛屬于沼澤型水牛,灰色被毛的水牛占到總數的73.19%,少部分為白色,占總數的5.7%[3],其中白色對灰色表現完全顯性。近年來,由于不科學的選育與保種不當、近親交配等原因,導致白色被毛水牛品種嚴重退化,群體數量不斷遞減[4]。皮、毛的顏色作為家畜最直觀的外部表型,不僅對其經濟價值有很大影響,還是品種的重要特征之一,與毛色性狀相關的候選基因多態性不僅是可以利用的良好遺傳標記,在確定雜交組合和品種純度等方面也有重要用途,此外還有研究表明動物的毛色與乳、肉等生產性能有直接的關系[5],對毛色與疾病的相關性研究也為診斷和治療相關疾病提供了理想的參考。本試驗旨在分析貴州白水牛的白色被毛形成的內在分子機制,以豐富對其品種資源遺傳特征的認識,為今后白水牛品種的保種和選育提供科學的指導。
牛的毛色主要是由皮膚和毛發中的真黑色素(褐與黑)與偽黑色素(紅與黃)的相對數量與分布情況所決定的[6]。許多研究表明,ASIP基因、MC1R基因和TYR基因是控制牛、羊等哺乳動物毛色的重要主效基因,這些基因的突變可能引起動物毛色性狀的改變,故本試驗通過對這3個基因的多態性研究,試圖從中分析得到貴州白水牛白色被毛形成的原因。
1 材料與方法
1.1 試驗材料
試驗在不同品種飼養管理條件基本相同的情況下選取健康無病、精神狀態良好、被毛光澤、顏色均一的水牛為樣本,40頭貴州水牛(20頭貴州白水牛和20頭貴州灰水牛)選自貴州省遵義市鳳崗縣養殖廠的水牛個體,剪取耳組織為樣本;40頭普通黑水牛采取隨機抽樣的方式從宜賓水牛育種場采集血樣,每頭牛采集10 mL頸靜脈血樣置于抗凝血采血管中,震蕩混勻后置于-80 ℃保存。
1.2 提取DNA和構建DNA池
血液樣本采用全血/組織/細胞基因組DNA快速提取試劑盒,提取血液基因組DNA;耳組織樣本采用磁珠法動物組織基因組DNA抽提試劑盒提取其DNA,用NanoDropTM 1000 Spectrometer(Thermo Fisher Scientific)檢測DNA濃度及純度(OD260/280,OD260/230),并將水牛DNA樣品的濃度調整至50 ng/μL。從10頭貴州白水牛的DNA樣品中各取5 μL混合在一起構建貴州白水牛的DNA池,從10頭貴州灰水牛耳部肌肉組織樣DNA中各取5 μL均勻混合在一起構建灰水牛的DNA池,從20頭宜賓水牛頸靜脈血樣DNA中各取5 μL混合在一起構建普通黑水牛的DNA池。
1.3 引物設計和PCR擴增
參照NCBI數據庫GenBank中提供的水牛(buffalo)ASIP基因序列(NC_037558.1)、MC1R基因序列(AC_000175.1)、TYR基因序列(AC_000186.1),用Primer 3.0 Plus軟件設計這3個基因的啟動子、外顯子、3′-UTR和5′-UTR的特異性引物,引物由上海生工Sangon Biotech生物公司合成(詳見表1~3)。
PCR反應總體系25 μL,其中ddH20 9.5μL,Mix(包括PCR穩定劑和增強劑、Mg2+、dNTPs、Taq DNA Polymerase、PCR Buffer))12.5 μL,上游引物F和下游引物R各1 μL,水牛DNA 1 μL。PCR反應條件:95 ℃預變性5 min;94 ℃變性30 s;設計不同的退火溫度30 s,72 ℃延伸30 s,共32個循環;最后72 ℃延伸10 min;4 ℃保存。PCR產物用1%瓊脂糖凝膠電泳30 min檢測,通過電泳檢測結果,判斷引物是否合適,并確定最適退火溫度。
1.4 序列測序及分析
以貴州白水牛、貴州灰水牛和普通水牛分別構建的DNA池作為DNA模板進行特定部分PCR擴增,將PCR擴增產物送去上海生工生物工程股份有限公司進行正、反測序。測序結果用SeqMan進行對比分析尋找突變位點。
1.5 擴大樣本驗證
針對以DNA池為模板檢測出來的突變位點,利用剩下的10頭貴州白水牛個體、10頭貴州灰水牛個體和20頭普通水牛個體的DNA樣本為模板對突變位點所在的區域進行PCR擴增,測序分析,對比篩選到的突變位點是否具有特異性,能否解釋品種間毛色的差異。

表1 ASIP引物設計

表2 MC1R引物設計

表3 引物設計
2 結果與分析
通過對測序結果的分析,發現貴州白水牛的ASIP、MC1R和TYR基因都存在多態位點,以MC1R基因第一外顯子第一位堿基計數為1,貴州白水牛的SNPs位點為:exon 1-G617A、exon 1-T843C,這2個SNPs位點在貴州灰水牛和部分普通水牛個體中同樣存在,不具有特異性,詳見圖1;以TYR基因每一外顯子第一位堿基計數為1,貴州白水牛的SNPs位點為:exon 1-G826A、exon 3-A83G、exon 5-T70C和exon 4-C170A,這4個SNPs位點在貴州灰水牛和普通水牛中同樣存在,不具有特異性,詳見圖2;以ASIP基因第一外顯子第一位堿基計數為1,貴州白水牛的SNPs位點為:Promoter-A147T和Promoter-A235G,該突變在部分普通水牛個體樣本中同樣存在,故在白水牛品種中不具有特異性,不能解釋白色被毛形成的原因,詳見圖3。
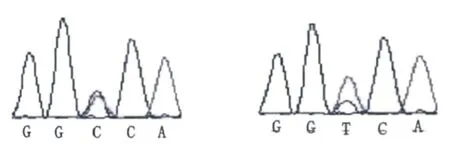
B:exon 1-T843 C Q:exon 1-T843 C

B:exon 1-G826A exon 3-A83G exon 5-T70C

Q:exon 1-G826A exon 3-A83G exon 5-T70C
3 結 論
利用在線軟件https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html預測貴州白水牛MC1R基因及TYR基因突變前后蛋白質二級結構的變化,發現在MC1R基因中exon 1-T843C為錯義突變,導致纈氨酸(Val)變成丙氨酸(Ala);TYR基因中exon 1-G826A和exon 5-T70C為錯義突變,分別導致精氨酸(Arg)突變為組氨酸(His)、亮氨酸(Leu)突變為異亮氨酸(Ile),exon 3-A83G為同義突變;在ASIP的啟動子中發現了2個SNPs位點:Promoter-A147T和Promoter-A235G。擴大個體樣本進行測序驗證這些SNPs位點,發現它們在貴州白水牛中不存在特異性,在其他非白水牛的個體中也有同樣突變,故不能解釋貴州白水牛白色被毛形成的分子機制,對其毛色形成原因的分析,有待下一步試驗探討。
4 討 論
ASIP和MC1R是哺乳動物體內調控毛色的2個重要候選基因,ASIP的表達會引起褐黑素的產生,MC1R的表達會引起真黑素的產生,許多研究表明,MC1R基因與ASIP基因共同通過控制紅/黃褐黑素和黑/棕真黑素之間的相對數量,從而控制動物的毛色[7]。一方面ASIP基因編碼Agouti信號蛋白(agouti signaling protein,ASIP)[8],ASIP是一種旁分泌信號分子,對G蛋白共軛型受體有天然的拮抗作用,以競爭方式對黑色素細胞刺激激素受體(MSHR)起拮抗作用,引起環腺苷酸水平下降而產生棕黑素;另一方面,ASIP還可以下調MC1R信號,通過氨基末端,降低TYR和Dct(多巴色素異構酶)水平[9],通過以上這2種方式,ASIP使真黑素合成過程停止,轉為合成褐黑素,從而使皮毛表現為黃色或紅色。此外,ASIP基因還編碼一種Agouti基因相關蛋白(agouti-related protein, AgRP),AgRP的N末端可以除去與ASIP具有高親和性的MC4R,使ASIP產生生理作用,從而達到與ASIP共同調節動物毛色的功能[10]。結合國內和國外已有的研究表明,MC1R基因是調控牛毛色的重要候選基因之一,具有不同MC1R基因型的牛,其毛色存在差異。早期的研究發現,牛MC1R基因有ED、E+和e 3種基因型,Francois等在2000年通過COP-PCR的方法在牛中還檢測到一種新的基因型E1[11]。TYR基因是編碼酪氨酸酶的基因,酪氨酸酶是黑色素形成過程中的關鍵酶,其表達量和活性決定了黑色素生成的速度和多少。黑色素是多巴醌、吲哚-5,6醌和多巴色素的聚合物,酪氨酸酶正是把酪氨酸轉化成多巴的中介,多巴再被氧化成多巴醌,進一步轉變為各種衍生物組合構成黑色素。鑒于酪氨酸酶對黑色素形成的重要性,TYR基因也被列為與毛色性狀有關的重要研究基因之一。許多研究都表明,這3種基因的遺傳多態性與動物的毛色性狀顯著相關。
研究一方面在貴州白水牛MC1R基因和TYR基因中篩選到突變位點,驗證了敖啟燕等[12]對貴州水牛毛色研究的結果,同時擴大了水牛品種分析的范圍,但發現這些SNPs位點在其他黑水牛品種中也存在,不具有特異性,因此不能解釋其特殊白色被毛形成的原因。接下來,筆者準備擴大群體分析這些突變位點在貴州白水牛和非白水牛品種之間的突變頻率是否有顯著差異,因為毛色也可能是多基因控制。另一方面,毛色相關基因很多,本研究初步分析了3個毛色相關基因的突變情況,可以為下一步尋找其他可能的因果基因奠定基礎,試驗結果豐富了我們對貴州白水牛毛色性狀遺傳機理的認識,也為貴州白水牛的分子選育工作提供了一定的理論依據。
猜你喜歡
今日農業(2021年21期)2021-11-26 05:07:00
鄉村地理(2019年2期)2019-11-16 08:49:28
鄉村地理(2018年3期)2018-11-06 06:51:08
鄉村地理(2018年3期)2018-11-06 06:51:06
鄉村地理(2018年3期)2018-11-06 06:50:10
貴茶(2018年6期)2018-05-30 09:53:50
貴茶(2018年6期)2018-05-30 09:53:36
鄉村地理(2018年4期)2018-03-23 01:54:04
鄉村地理(2016年2期)2016-06-15 20:29:24
貴州師范學院學報(2016年8期)2016-02-27 12:48:14
