β一胡蘿卜素一15,15’一加氧酶在利魯牛各組織部位的表達
2019-08-03 07:16:06萬發春魏晨張相倫譚秀文劉桂芬游偉趙紅波王星凌劉曉牧靳青
山東農業科學 2019年3期
關鍵詞:檢測
萬發春 魏晨 張相倫 譚秀文 劉桂芬 游偉 趙紅波 王星凌 劉曉牧 靳青

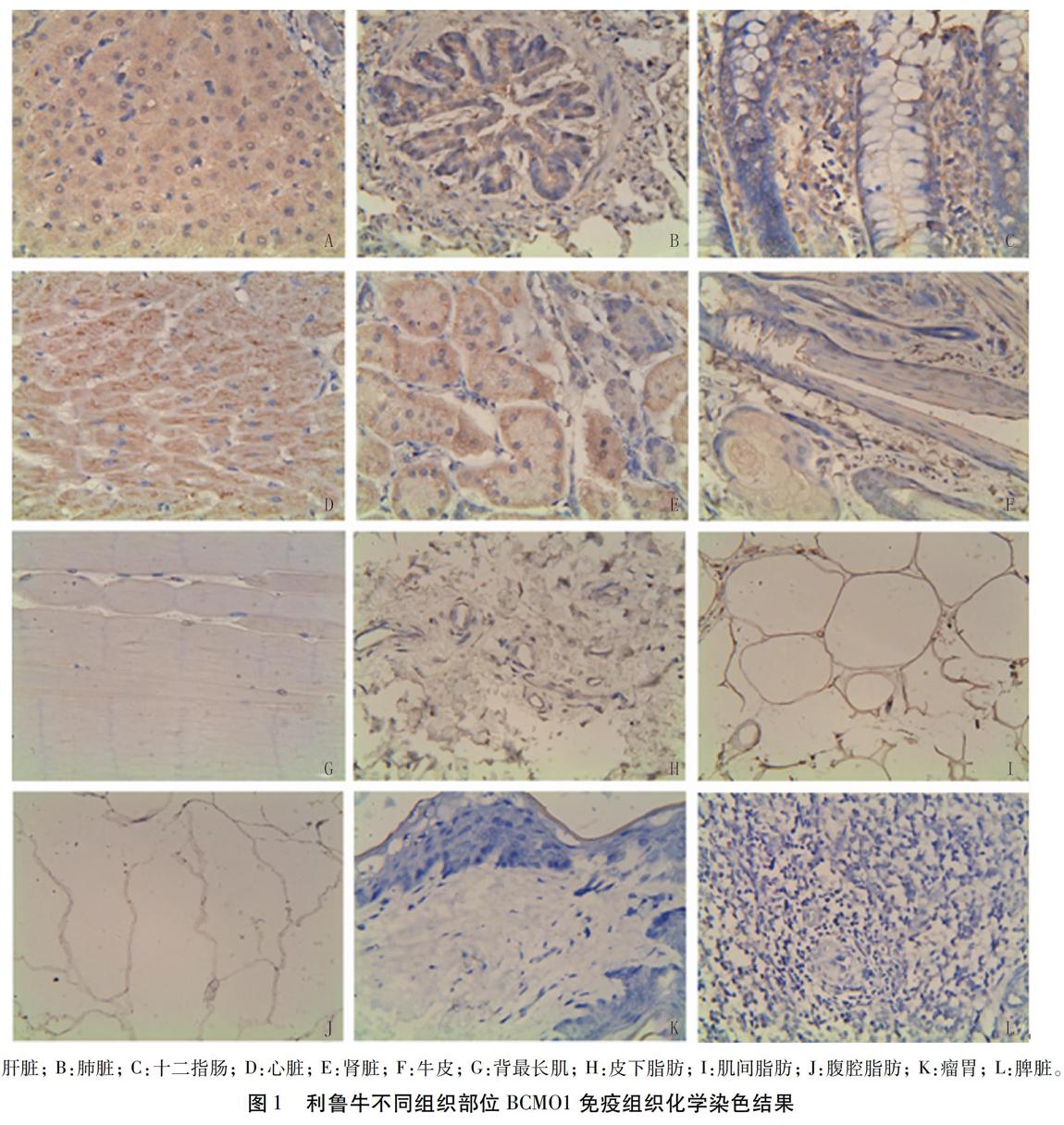

摘要:本試驗以利魯牛(利木贊牛×魯西黃牛)為研究對象,通過免疫組織化學、蛋白質印跡法( Westernblot)和熒光定量PCR等分子手段檢測了BCMOI在利魯牛肝臟、十二指腸、不同脂肪組織等部位的表達。免疫組織化學和Westem Blot檢測結果均顯示,BCM01在肝臟、十二指腸、心臟、腎臟和肺臟中具有較高的表達量,在脾和瘤胃內未表達;不同部位脂肪組織內BCMOI相對表達量差異較大,其中在Western Blot檢測中BC-MOI在皮下脂肪僅有極少量表達,但免疫組織化學結果顯示BCMOI在不同脂肪組織中的表達量相當。熒光定量PCR檢測結果顯示,BCM01除在十二指腸具有極高的表達量外,在腎臟、心臟、牛皮的表達量均高于肝臟。以上結果對深入研究p一胡蘿卜素在肉牛體內的代謝機制提供了科學支撐。
關鍵詞:利魯牛;B一胡蘿卜素- 15,15' -加氧酶;免疫組織化學;蛋白質印跡法;熒光定量PCR;表達量
Expression of [3 - Carotene - 15 ,15' - Monooxygenase(BCM01) in Tissues of Lilu Cattle Wan Fachun, Wei Chen, Zhang Xianglun, Tan Xiuwen, Liu Guifen, You Wei,Zhao Hongbo, Wang Xingling, Liu Xiaomu, Jin Qing
Abstract
The study analyzed the expression of BCM01 in twelve tissues (liver, duodenum, adipose tis-sues and so on) of Limusin X Luxi( Lilu) bull by immunohistochemistry, Western - blot and qRT - PCR. Im-munohistochemical and Western - blot results showed that BCM01 was highly expressed in liver, duodenum,heart and lung, but no expression was detected in spleen and rumen, and the relative expression levels of BC-M01 in different adipose tissues were different. The Western - blot results showed that BCMOI was only ex-pressed in a very small amount in subcutaneous fat tissue, but the immunohistochemical results showed thaithe expression levels of BCM01 in the three fat tissues were similar. Extremely higher BCM01 mRNA relativeexpression was detected in duodenum by qRT - PCR, and the levels in kidney, heart, and skin were all high-er than that in liver. These results provided scientific supports for further research on the metabolic mechanismof beta carotene in beef cattle.
Keywords
Lilu cattle;β- carotene - 15.15' - momoxygenase; Immunohistochemistry; Western -blot; RT - qPCR; Expression quantity
β-胡蘿卜素- 15,15' -加氧酶(β-carotene -15,15-momoxygenase 1,BCM01)是類胡蘿卜素向維生素A轉化過程中的關鍵酶,催化β-胡蘿卜素的中間斷裂,生成2分子視黃醛,進一步轉化為不同形式的維生素A。BCM01對類胡蘿卜素中15,15位置的雙鍵有高度的特異性,在大鼠上β-胡蘿卜素的這種中心裂解是產生維生素A的主要途徑[1]。人類吸收的35%~90%全反式β-胡蘿卜素是在BCM01催化下裂解進而合成維生素A。Hessel等在敲除小鼠BCM01的試驗中發現,BCM01功能的缺失能夠導致維生素A缺乏,當進一步喂食富含β-胡蘿卜素的日糧后,小鼠的肝臟、肺臟和脂肪組織均有大量的β-胡蘿卜素積累[2]。可見BCM01在動物體內維生素A合成過程中發揮著不可替代的作用。
目前,BCM01基因已在人、鼠、豬、牛等哺乳動物以及果蠅、雞、海洋細菌、斑馬魚等物種內被克隆[3],其中Lindqvist等[4]首次獲得完整的人類BCM01的分子克隆。早期研究只在人、小鼠、大鼠、豚鼠、兔子、猴子等哺乳動物的腸黏膜中檢測到BCM01的活性[5,6],后續發現其在人多數器官組織的上皮細胞中表達[7,8]。上皮細胞通常是對維生素A缺乏非常敏感的結構,盡管BCM01的體外功能尚不清楚,但其在被檢測組織的上皮細胞中均有表達,暗示BCM01對維生素A的局部合成非常重要。當膳食中維生素A攝人量不足時,BCM01構成了維生素A合成的一條備用途徑。
在牛上,BCM01對胡蘿卜素的催化活性首先在卵巢和黃體上被發現[9,10],隨后在肝臟和腸道不同部位及性腺組織內被陸續報道[ll]。但BC-M01在肉牛上述組織之外的細胞類型中的表達情況和可能起到的作用仍有待于進一步探索。基于此,本試驗對BCM01在肉牛體內12個組織部位的表達情況進行檢測,以期為深入研究肉牛體內β-胡蘿卜素的代謝機制提供科學支撐。
1 材料與方法
1.1 樣品采集
選取4頭利魯牛(利木贊牛×魯西黃牛)成年育肥公牛作為試驗用牛。采集心臟、肝臟、腎臟、肺臟、脾臟、瘤胃、十二指腸、牛皮、背最長肌、皮下脂肪、肌間脂肪、腹腔脂肪共12個組織部位樣品,一部分分裝于冷凍管中并立即投入液氮中帶回實驗室-80℃冰箱保存(用于mRMA和蛋白表達檢測),另一部分浸泡于含4%多聚甲醛的樣品瓶中(用于免疫組織化學檢測)。
1.2 主要試劑
超純RNA提取試劑盒(Ultrapure RNA Kit)購自康為世紀生物有限公司;逆轉錄試劑盒( HiS-cript⑧ⅡReverse Transcriptase)和熒光定量檢測試劑盒( ChamQ'TM SYBR Color qPCR Master Mix)均購自南京諾唯贊生物科技有限公司。
1.3 引物設計與合成
根據GenBank公布的牛BCM01基因的mR-NA序歹0號,利用Primer 5.0和NCBI中BLAST工具設計實時熒光定量檢測引物,以β-actin作為內參基因。引物由鉑尚生物技術(上海)有限公司合成,具體信息如下:
1.4 BCM01蛋白水平表達的免疫組織化學檢測
組織部位樣品用4%多聚甲醛固定,酒精和二甲苯脫水,石蠟包埋,再經切片、制片后室溫脫蠟、水化,后對內源酶進行滅活,加入山羊血清封閉非特異位點,再加I抗4℃過夜,然后經Ⅱ抗、鏈霉菌抗生物素一過氧化物酶溶液孵育,后用DAB溶液顯色,蘇木素染核,再經鹽酸酒精分化,石蠟脫水,最后用中性樹膠封片,置于顯微鏡下進行觀察、拍照。
1.5 BCM01蛋白表達水平Western Blot檢測
取適量肉牛組織加100 μL含PMSF的RIPA蛋白裂解液,4℃裂解30 min,期間EP管要經常晃動以促進細胞充分裂解;4℃下12000 r/min離心10min,將上清液轉移至新離心管中,得組織蛋白樣品。BCA法檢測蛋白濃度,根據濃度計算30μg不同蛋白樣品的上樣體積,將不同上樣體積的蛋白樣品分別與5×十二烷基硫酸鈉(SDS)緩沖液混勻,沸水中煮沸5 min,于冰浴中迅速冷卻,后上樣進行SDS-PAGE電泳(25 mA恒流電泳至溴酚藍跑完濃縮膠層,時間大約為30 min。分離膠電泳電流為30 mA,當溴酚藍遷移至分離膠下緣時,關掉電源,停止電泳);電泳后轉PVDF膜,電流為200 mA;將PVDF膜浸入含5%脫脂奶粉封閉液中室溫搖床封閉1h;取出已封閉的PVDF膜,浸于TBST緩沖液中洗滌后移人含有一抗的小袋中,4℃孵育過夜,TBST緩沖液搖床洗滌3次,每次15min;將PVDF膜轉移到含二抗的玻璃平皿中室溫搖床孵育2h,TBST緩沖液搖床洗滌3次,每次15min;將PVDF膜置于保鮮膜上,取適量ECL試劑盒中等體積的A液和B液混合,混勻后加在膜的表面,移人凝膠成像分析儀中,化學光敏模式曝光顯影。
1.6 BCM01基因實時熒光定量PCR
取各組織樣品約50mg,液氮研磨后加入1mL Trizol,提取總RNA,微量分光光度計檢測總RNA濃度,1%瓊脂糖凝膠電泳檢測RNA完整性。按照逆轉錄試劑盒進行反轉錄,反應體系20μL: RNA 3μL,4×gDNA wipe Mix 4μL, RNAase -free ddH20 9μL,5×HiScriptⅡqRT SuperMixⅡ4μL;反應程序:42℃ 2 min,50℃ 15 min,85℃ 5s。逆轉錄所得cDNA模板按照熒光定量試劑盒進行實時熒光定量PCR檢測。反應體系20μL:cDNA lμL,上下游引物各0.4μL,2×ChamQTMSYBR Color qPCR Master Mix 10μL,ddH20 8.2μL。反應程序:95C預變性30 s;95qC變性10 s,60℃退火30 s,40個循環;95℃延伸15s,60℃收集熒光1min。每個樣品測定重復3次。
1.7數據處理與分析
將免疫組織化學染色結果和Western Blot膠片掃描圖片用Quantity One圖像分析軟件分析目標條帶的光密度值,所得數據利用SAS 9.1軟件的一般線性模型進行單因素方差分析進行統計,利用Duncan,s法進行多重比較,P<0.05表示差異顯著。
2 結果與分析
2.1 免疫組織化學檢測結果
免疫組織化學檢測結果顯示,在所有檢測的組織部位中,BCM01在十二指腸、心臟和腎臟中表達量相當(P>0.05),顯著高于其它組織部位(P<0.05),其次為肺臟;在牛皮內的表達量略高于肝臟;在不同部位脂肪組織中表達量差異不大;在背最長肌中表達量顯著低于其它組織(P<0.05);在瘤胃和脾臟中未檢測到BCM01的表達(圖l、圖2)。
2.2 BCM01蛋白Western Blot檢測結果
蛋白印跡檢測結果顯示,在所有檢測的組織部位中,BCM01在肺臟內的表達量最高,其次為十二指腸和心臟,在肝臟和腎臟內的表達量相當(P>0.05);在不同部位脂肪組織中,肌間脂肪組織內表達量最高,腹腔脂肪中的略低,而在皮下脂肪組織中表達量顯著低于其它部位(P<0.05);瘤胃和脾臟中未檢測到BCM01的表達(圖3)。
2.3 BCM01熒光定量PCR檢測結果
BCM01基因mRNA在十二指腸表達水平極高,為肝臟的378倍,其次為腎臟、心臟、牛皮,表達量分別為肝臟的130倍、16倍和3.7倍,牛皮內的表達量為肝臟的3.7倍,其它組織部位BC-M01相對表達量均低于肝臟,但差異不顯著(P>0.05),瘤胃內檢測到微量的表達,脾臟內未檢測到BCM01的相對表達(圖4)。
3 討論與結論
20世紀60年代,Lietz等[12]通過大鼠試驗研究發現,β-胡蘿卜素轉變為維生素A是由一種小腸粘膜和肝臟中的可溶性β-胡蘿卜素- 15,15'-加氧酶通過一種假定的橫跨B-胡蘿卜素分子中間15,15雙鍵上的雙氧四環結構媒介物完成。目前對這種酶的性質已經了解,它是一種水溶性胞漿酶,最適pH值稍偏堿性,裂解中需使用分子氧[13]。由于在最早的研究中沒有發現任何其它的反應產物,因此稱這種酶為β-胡蘿卜素- 15,15' -加氧酶,主要存在于小腸粘膜和肝臟細胞的胞漿中[14]。
小腸是β-胡蘿卜素在動物體內的主要吸收場所,具有較強的轉化維生素A的能力;肝臟是β-胡蘿卜素在肉牛體內的主要存儲和轉化場所,因此BCM01在肝臟和小腸中均具有較高的表達量[15],與本研究檢測結果一致。BCM01也是動物體脂肪代謝的重要調控者,通過BCM01基因敲除小鼠試驗發現,血清脂質水平發生改變并且引起其肝臟脂肪變性[2]。本團隊早期研究也發現,日糧添加B -胡蘿卜素可顯著降低肉牛背部皮下脂肪厚度,但對肌間脂肪含量并無顯著影響,在此過程中BCM01可能發揮著重要作用[16],暗示β-胡蘿卜素或BCM01對脂質代謝具有重要的調節作用。
Lindqvist等[7]通過免疫組織化學染色發現,BCM01在人多數部位的上皮細胞中均有表達,包括胃粘膜和腺細胞、小腸、結腸、肝臟實質細胞、胰腺外分泌腺細胞、前列腺腺細胞、子宮內膜、乳腺組織、腎小管細胞、皮膚鱗狀上皮細胞的角質形成細胞,此外,在睪丸、卵巢、腎上腺以及骨骼肌細胞中的類固醇生成細胞中也有表達。除上述組織部位之外,BCM01在視網膜色素上皮細胞中大量表達[8,17]。β-胡蘿卜素除為動物提供機體所需的維生素A、參與脂肪代謝調控外,還在保護皮膚、提高免疫力、延緩衰老、預防心血管疾病以及防治白內障、關節炎、糖尿病、腎小球炎等慢性疾病方面具有一定的功效,這可能是本研究中在肉牛體內除十二指腸、肝臟和脂肪組織外,在心臟、腎臟、肺臟、皮膚等組織器官內檢測到BCM01表達的重要原因之一,具體機制有待進一步研究。
反芻動物的瘤胃微生物無法合成維生素A,只能通過魚粉、血粉等動物性飼料補充或通過飼喂青草、青貯料等富含胡蘿卜素的粗飼料在肝臟內轉化合成維生素A,這可能是本研究中未檢測到BCM01在肉牛瘤胃內表達的原因之一。脾臟是動物機體最大的免疫器官,是機體細胞免疫和體液免疫的中心,本研究未檢測到BCMOI在肉牛脾臟內表達,暗示β-胡蘿卜素可能通過其它途徑提高肉牛免疫力,與畢宇霖等[18]的研究結果(β-胡蘿卜素通過提高機體抗氧化能力從而提高肉牛免疫力)相吻合。
綜上,BCMOI在肉牛體內的普遍表達不僅能為各組織器官提供正常生理功能所需的維生素A,在維持與調節心、肺、腎功能方面也具有重要作用,而其具體作用和調控機制有待進一步研究。
參考文獻:
[1] Paik J,During A,Harrison E H,et al.Expression and char-acterization ofa murine enzyme able to cleave beta - carotene.The formation of retinoids[J].Journal()f Biological Chemistry,2001,276(34): 32160 - 32168.
[2]Hessel S,Eichinger A,Isken A,et al.CM01 defiCiencv abol-ishes vitamin Aproduction from beta - Carotene and alters lipidmetaholism in miCe[J].Joumal of Biologcal Chemistry.2007.282(46):33553 - 33561.
[3] 靳青,畢宇霖,劉曉牧,等.類胡蘿卜素代謝及功能研究進展[J].動物營養學報,2014,26(12):3561 - 3571.
[4] Lindqvist A,Andersson S. Biochemical properties of purifiedrecombinant human beta - carotene 15, 15- monooxygenase[J].Journal of Biological Chemistry, 2002, 277 (26) :23942 - 23948.
[5]von Lintig J. Colors with functions : elucidating the biochemicaland molecular basis of carotenoid metabolism[J] . Annual Re-view of Nutrition , 2010 . 30 : 35 - 56.
[6]During A, Nagao A, Hoshino C. et al. Assay of β- carotene15 .15 ' - dioxygenase activity by reverse - phase high pressureliquid chromatography [J] .Analytical Biochemistry , 1996 , 241(2) : 199 -205.
[7]Lindqvist A, Andersson S. Cell type - specific expression ofβ- carotene 15,15 ' - mono - oxygenase in human tissues [J] .Journal of Histochemistry & Cytochemistry, 2004 . 52 (4) :491 - 499.
[8]Yan W, Jang C F, Haeseleer F, et al. Cloning and character-ization of a human beta , heta - carotene - 15 , 15 ' - dioxygenasethat is highly expressed in the retinal pigment epithelium [J] .Genoruics . 2001 , 72 (2) : 193 - 202.
[9]Sklan D. Carotene cleavage activity in the corpus luteum of cat-tle[J] Intemational Journal for Vitamin and Nutrition Re-search.1983 .53 (l) : 23 - 26.
[10] Schweigert F, Wierich M. Rambeck W. et al. Carotene cleav-age activity in bovine ovarian follicles [J] Theriogenology,1988. 30(5) : 923 -930.
[11] Morales A, Rosas A. Gonzalez A. et al. Cloning of the bovinebeta - carotene - 15 . 15 ' - oxygenase and expression in gonadaltissues [J] . Infemational Joumal for Vitamin and Nutrition Re-search.2006 .76(l) : 9 - 17.
[12] Lietz C, Lange J. Rimbach G. Molecular and dietary regula-tion of beta, heta - carotene 15. 15' - monooxygenase I(BCM01) [J] Archives of Biochemistry and Biophysics,2010 . 502(1) :8 - 16.
[13] von Lintig J, Vogt K. Filling the gap in vitamin A research.Molecular identification of an enzyme c:leaving beta - caroteneto retinal [J] .Journal of Biological Cheruistry ,2000 .275 (16) :11915 - 11920.
[14] Redmond T M. Centleman S, Duncan T, et al. Identification,expression . and substrate specificity of a mammalian heta - car-otene 15 . 15 - dioxygenase [J] . Journal of Biological Chemis-try, 2001 .276 (9) : 6560 - 6565.
[15] Shmarakov I O.Yuen J J. Blaner W S. Carotenoid metabolismand enzymology [M]//TANUMIHARDJO S A. Carotenoidsand human health. Nutrition and health. New York: HumanaPress. 2013 :29 - 56.
[16] Jin Q, Cheng H, Wan F. et al. Effects of feeding β- caroteneon levels of β- carotene and vitamin A in blood and tissues ofheef cattle and the effects on heef quality [J] . Meat Sciem:e,2015 . 110 : 293 - 301.
[17] Chichili G, Nohr D. Schaffer M. et al. β - carotene c:onver-sion into vitamin A in huruan retinal pigment epithelial cells[J] Investigative Ophthalruology & Visual Science. 2005 .46 : 3562 - 3568.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
