SARA低表達(dá)對(duì)Activin A/Smads環(huán)路活性的影響
2019-08-02 08:39:54石曉花王姣琦劉洪雨紀(jì)秋野何金婷徐忠信
中國(guó)實(shí)驗(yàn)診斷學(xué) 2019年7期
石曉花,王姣琦,劉洪雨,董 玥,紀(jì)秋野,崔 楊,何金婷,莽 靖,徐忠信
(1.吉林大學(xué)臨床醫(yī)學(xué)院,吉林 長(zhǎng)春130021;2.吉林大學(xué)中日聯(lián)誼醫(yī)院)
激活素A(ActA)是目前較公認(rèn)的神經(jīng)保護(hù)因子[1,2]。研究表明ActA主要通過ActA/Smads線性跨膜信號(hào)轉(zhuǎn)導(dǎo)將胞外信號(hào)傳遞至胞內(nèi)[3-5]。近期研究發(fā)現(xiàn)ActA/Smads信號(hào)具有環(huán)路作用模式[6]。但該環(huán)路活性是否受其他因子調(diào)控尚不清楚。Smad錨捉蛋白(SARA)作為R-Smads的磷酸化輔助因子,其在經(jīng)典ActA/Smads線性通路中的功能已經(jīng)被證實(shí)[7],但在ActA環(huán)路的調(diào)控作用尚不明確。本研究利用靶向大鼠SARA基因的shRNA轉(zhuǎn)染下調(diào)SARA表達(dá),Western blot通過對(duì)Smad3總蛋白及磷酸化蛋白水平進(jìn)行檢測(cè)評(píng)估ActA環(huán)路活性,為進(jìn)一步明確ActA/Smads環(huán)路調(diào)控機(jī)制提供實(shí)驗(yàn)基礎(chǔ)。
1 材料和方法
1.1 主要材料
PC12細(xì)胞(大鼠腎上腺嗜鉻細(xì)胞瘤)由國(guó)家實(shí)驗(yàn)細(xì)胞資源共享服務(wù)平臺(tái)提供。ActA高表達(dá)的PC12細(xì)胞由本課題組提供[8]。G418購(gòu)自美國(guó)GIBCO公司。SARA及內(nèi)參GAPDH基因引物由上海生工公司合成。LipofectamineTM2000脂質(zhì)體轉(zhuǎn)染試劑購(gòu)自美國(guó)Invetrogen公司。Quantscript cDNA第一鏈合成試劑盒及Real Master Mix(SYBR Green)試劑盒購(gòu)自天根生化科技(北京)有限公司。兔抗大鼠Smad3、磷酸化Smad3抗體購(gòu)自美國(guó)ThermoFisher公司,小鼠抗大鼠β-actin抗體購(gòu)自美國(guó)Santa公司。辣根過氧化物酶標(biāo)記的羊抗小鼠、羊抗兔抗體購(gòu)自北京博奧森生物技術(shù)有限公司。
1.2 實(shí)驗(yàn)方法
1.2.1PC12細(xì)胞及ActA高表達(dá)細(xì)胞的培養(yǎng) PC12細(xì)胞常規(guī)培養(yǎng)于含10%馬血清、10%胎牛血清的高糖DMEM培養(yǎng)液。每2-3天根據(jù)細(xì)胞生長(zhǎng)情況及培養(yǎng)液顏色更換培養(yǎng)液,細(xì)胞接近80 %融合時(shí)用0.25 %的胰酶進(jìn)行消化傳代。構(gòu)建靶向大鼠INHBA基因的重組真核表達(dá)載體,脂質(zhì)體轉(zhuǎn)染PC12細(xì)胞后G418(終濃度300 μg/ml)篩選2個(gè)月,獲得ActA高表達(dá)的PC12細(xì)胞,隨后將G418的濃度降為150 μg/ml進(jìn)行維持篩選。除加入G418外,ActA高表達(dá)PC12細(xì)胞的培養(yǎng)方法同前。
1.2.2shRNA質(zhì)粒的設(shè)計(jì)合成 從大鼠SARA基因(NM_001107952.1)編碼序列中選擇特異性靶序列,分別是第4324位 (shRNA1):5′-GATGACCACCGTACCTGCC-3′和第1081位 (shRNA2):5′-CAAATGGATGCTTTGAATAAA-3′。另設(shè)計(jì)2段同源的隨機(jī)對(duì)照序列作陰性對(duì)照。利用BLAST網(wǎng)站進(jìn)行靶序列及陰性對(duì)照序列的同源性分析。委托武漢市淅瑪生物技術(shù)有限公司使用pGenesil-3質(zhì)粒構(gòu)建shRNA表達(dá)載體。DNA測(cè)序驗(yàn)證成功后,將靶向SARA基因的質(zhì)粒命名為SARA-shRNA;對(duì)照質(zhì)粒命名為NC-shRNA。
1.2.3shRNA質(zhì)粒瞬時(shí)轉(zhuǎn)染及實(shí)驗(yàn)分組 將ActA高表達(dá)的PC12細(xì)胞接種于6孔板,細(xì)胞70-80%融和時(shí)按照shRNA轉(zhuǎn)染說明書進(jìn)行操作。轉(zhuǎn)染48 h后避光條件下,倒置熒光顯微鏡觀察細(xì)胞轉(zhuǎn)染效率并攝圖。以ActA高表達(dá)的PC12細(xì)胞作為正常對(duì)照,記為Control組;SARA-shRNA轉(zhuǎn)染的ActA高表達(dá)細(xì)胞作為陽性轉(zhuǎn)染組,記為SARA-shRNA組;NC-shRNA質(zhì)粒轉(zhuǎn)染的ActA高表達(dá)細(xì)胞作為陰性對(duì)照,記為NC-shRNA組。
1.2.4實(shí)時(shí)定量PCR檢測(cè)SARA基因mRNA的表達(dá) 收集上述三組細(xì)胞,Trizol法提取總RNA,逆轉(zhuǎn)錄制備cDNA。基因檢測(cè)引物如下:SARA F:5′-TACAGATCAGCCAACCTGAGGA-3′,R:5′-ATACA ATTGGGCTCTGCATTCC -3′,擴(kuò)增長(zhǎng)度192 bp;GAPDH F:5′-GGTTACCAGG GCTGCCTTCT-3′,R:5′-ATGGGTTTCCCGTTGATGAC-3′,擴(kuò)增長(zhǎng)度171 bp。按照實(shí)時(shí)定量PCR說明書步驟配置反應(yīng)體系,設(shè)定PCR運(yùn)行參數(shù)。PCR反應(yīng)使用ABI7500Fast 型實(shí)時(shí)熒光定量PCR儀,熒光收集及分析利用ABI7500Fast 程序軟件。應(yīng)用Relative Quantification (ddCt) Study進(jìn)行相對(duì)表達(dá)量(relative quantity,RQ)RQ分析,計(jì)算SARA基因的相對(duì)表達(dá)水平。檢測(cè)實(shí)驗(yàn)重復(fù)三次。
1.2.5Western blot檢測(cè)SARA及Smad3蛋白水平的表達(dá) 提取各組總蛋白,紫外分光光度計(jì)測(cè)定蛋白濃度并調(diào)整上樣量。每組蛋白100 ℃煮沸 5 min后上樣,經(jīng)電泳、轉(zhuǎn)膜、封閉后用含5%脫脂奶粉的TBST溶液稀釋抗體,分別為兔抗大鼠SARA(1∶10000),Smad3(1∶1000),磷酸化Smad3(1∶1000)和小鼠抗大鼠β-actin(1∶1000),一抗4 ℃ 過夜孵育。次日洗膜后二抗(1∶1000)室溫孵育1 h,ECL顯影壓膠片。凝膠圖像分析系統(tǒng)掃描膠片,使用吸光度分析計(jì)算目的條帶與相應(yīng)內(nèi)參條帶的灰度值,以兩者的比值表示目的蛋白的表達(dá)水平。
1.3 統(tǒng)計(jì)學(xué)分析
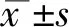
2 結(jié)果
2.1 shRNA瞬時(shí)轉(zhuǎn)染下調(diào)SARA基因表達(dá)
利用RNA干擾技術(shù),構(gòu)建靶向SARA基因的shRNA質(zhì)粒。該質(zhì)粒因攜帶DsRed2序列而在554 nm的激發(fā)光下產(chǎn)生紅色熒光。瞬時(shí)轉(zhuǎn)染Act A高表達(dá)的PC12細(xì)胞。轉(zhuǎn)染48 h,倒置熒光顯微鏡下觀察轉(zhuǎn)染效率,結(jié)果如圖1-A-D,明暗場(chǎng)對(duì)比,SARA-shRNA及NC-shRNA質(zhì)粒的轉(zhuǎn)染效率均超過70 %。實(shí)時(shí)定量PCR及Western blot檢測(cè)SARA蛋白水平的表達(dá),結(jié)果如圖1-E-F,靶向SARA基因的shRNA質(zhì)粒轉(zhuǎn)染可顯著下調(diào)SARA轉(zhuǎn)錄及蛋白水平的表達(dá)。
2.2 SARA低表達(dá)下調(diào)Act A/Smads環(huán)路活性
SARA低表達(dá)下調(diào)Smad3總蛋白及磷酸化蛋白水平。如圖2,SARA-shRNA組,細(xì)胞換液3 h,ActA環(huán)路自發(fā)活化后,與NC-shRNA組比較,Smad3總蛋白及磷酸化蛋白分別下調(diào)44%和57%。
3 討論
激活素A作為轉(zhuǎn)化生長(zhǎng)因子(TGF-β)超家族的成員之一,是目前較公認(rèn)的神經(jīng)保護(hù)因子,在包括缺血性腦血管病、神經(jīng)系統(tǒng)退行性變?cè)趦?nèi)的多種神經(jīng)系統(tǒng)疾病中發(fā)揮保護(hù)作用[1,2]。它是由兩個(gè)βA亞基通過二硫鍵連接成的二聚體結(jié)構(gòu)蛋白。β亞基是它的基本單位,由INHBA基因編碼。經(jīng)典研究認(rèn)為,ActA作為細(xì)胞外配體,主要通過ActA/Smads線性跨膜信號(hào)轉(zhuǎn)導(dǎo)將胞外信號(hào)傳遞至胞內(nèi),經(jīng)Smads級(jí)聯(lián)反應(yīng)完成膜受體激活的Smads (R-Smads),如Smad2、Smad3的磷酸化活化及核內(nèi)遷移,從而調(diào)控靶基因的表達(dá),發(fā)揮相應(yīng)的生物學(xué)功能[3-5]。課題組近期研究發(fā)現(xiàn),ActA/Smads信號(hào)還具有環(huán)路作用模式[6]。利用ActA基因高表達(dá)的PC12細(xì)胞進(jìn)行的研究發(fā)現(xiàn),隨著外源INHBA基因的高表達(dá),ActA合成增加并分泌到細(xì)胞外,其細(xì)胞外積累可觸發(fā)ActA信號(hào)短時(shí)活化,并反過來上調(diào)內(nèi)源性INHBA基因的表達(dá),從而構(gòu)成ActA/Smads 正反饋環(huán)路[8]。利用該細(xì)胞模型,實(shí)現(xiàn)了在非病理情況下模擬損傷性刺激介導(dǎo)的INHBA基因表達(dá)上調(diào)及ActA信號(hào)活化的生物學(xué)過程,并去除了外界損傷對(duì)細(xì)胞狀態(tài)及基因表達(dá)的影響,更利于探討ActA信號(hào)環(huán)路及其相關(guān)因子的表達(dá)變化及生物學(xué)功能。因此,本研究延續(xù)課題組前期工作,繼續(xù)利用ActA高表達(dá)的PC12細(xì)胞探討可能影響ActA環(huán)路活性的調(diào)控因子。
圖1 shRNA質(zhì)粒轉(zhuǎn)染下調(diào)SARA轉(zhuǎn)錄及蛋白表達(dá)
(A)NC-shRNA質(zhì)粒轉(zhuǎn)染48 h的明場(chǎng)圖;(B)NC-shRNA質(zhì)粒轉(zhuǎn)染48 h相應(yīng)的暗場(chǎng)圖;(C)SARA-shRNA轉(zhuǎn)染48 h的明場(chǎng)圖;(D)SARA-shRNA轉(zhuǎn)染48 h相應(yīng)的暗場(chǎng)圖。(標(biāo)尺=50 μm);(E)SARA轉(zhuǎn)錄水平的表達(dá)檢測(cè);(F)SARA蛋白表達(dá)及灰度分析。(*與Control組及NC-shRNA組比較P<0.05,n≥3/組)
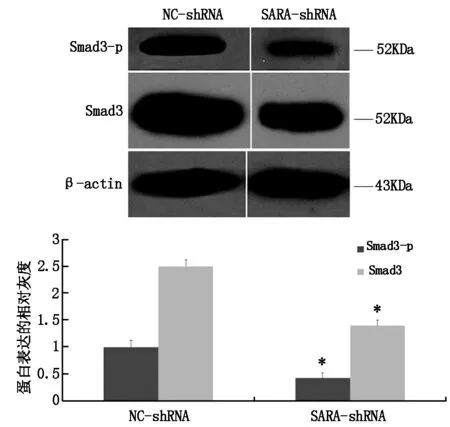
圖2 SARA低表達(dá)對(duì)Act A/Smads環(huán)路活性的影響
SARA作為R-Smads向膜受體募集的重要輔助因子,通過調(diào)控Smads與TGF-β受體的定位影響Smad2、Smad3的平衡和分布[9]。在缺血性腦損傷中,課題組前期研究發(fā)現(xiàn),SARA具有輔助R-Smads磷酸化活化,維持ActA/Smads線性通路活化的作用[7]。但在ActA信號(hào)環(huán)路中,SARA是否調(diào)控ActA環(huán)路活性尚不清楚。
本研究利用RNA干擾技術(shù)(RNAi),構(gòu)建靶向SARA基因的shRNA表達(dá)載體。該載體由啟動(dòng)子、發(fā)卡結(jié)構(gòu)序列和終止子組成,編碼出具有類似miRNA前體的莖環(huán)結(jié)構(gòu),經(jīng)核糖核酸酶Drosha酶切成shRNA前體(pre-shRNA),再在Exportin 5的作用下出核。出核后的pre-shRNA經(jīng)Dicer酶切后與RNA誘導(dǎo)的基因沉默復(fù)合物(RISA)結(jié)合,通過完全或不完全地與靶基因mRNA互補(bǔ)達(dá)到降解該基因 mRNA 或抑制 mRNA 翻譯蛋白質(zhì)的作用[10]。通過脂質(zhì)體瞬時(shí)轉(zhuǎn)染shRNA載體,本研究成功下調(diào)了ActA高表達(dá)細(xì)胞內(nèi)SARA基因mRNA及蛋白水平的表達(dá)。與正常的PC12細(xì)胞不同,ActA高表達(dá)細(xì)胞隨著外源性INHBA基因的轉(zhuǎn)錄翻譯,ActA表達(dá)升高并在細(xì)胞外積累,繼而觸發(fā)ActA信號(hào)活化,激活下游Smads級(jí)聯(lián)反應(yīng)。前期研究發(fā)現(xiàn)ActA高表達(dá)的PC12細(xì)胞換液3 h時(shí)ActA環(huán)路活化,表現(xiàn)為Smad3總蛋白及磷酸化蛋白表達(dá)升高,Smad4的核內(nèi)熒光強(qiáng)度增強(qiáng),內(nèi)源性INHBA 基因表達(dá)上調(diào)[8]。本研究細(xì)胞換液3 h時(shí)對(duì)Smad3總蛋白及磷酸化蛋白進(jìn)行檢測(cè)后發(fā)現(xiàn),SARA低表達(dá)組(SARA-shRNA組)Smad3總蛋白及磷酸化蛋白的水平較陰性轉(zhuǎn)染組(NC-shRNA組)明顯減低,說明SARA低表達(dá)降低了環(huán)路的活化水平,提示在ActA正反饋環(huán)路中SARA的表達(dá)水平仍是環(huán)路活性的一個(gè)調(diào)控因子。作為潛在干預(yù)位點(diǎn),進(jìn)一步探討SARA高表達(dá)對(duì)Act A/Smads環(huán)路活化時(shí)間及生物學(xué)功能的影響將為抗缺血性腦損傷位點(diǎn)的選擇提供新思路。
