利用微衛星和線粒體D-loop區聯合分析大尾寒羊的遺傳多樣性
2019-07-31 02:19:28劉澤民王巖超張淑二劉展生姜運良
山東畜牧獸醫 2019年7期
劉澤民 王巖超 張淑二 劉展生 姜運良*
利用微衛星和線粒體D-loop區聯合分析大尾寒羊的遺傳多樣性
劉澤民①王巖超①張淑二②劉展生②姜運良①*
(①山東農業大學動物科技學院 山東 泰安 271018 ②山東省畜牧總站 山東 濟南)
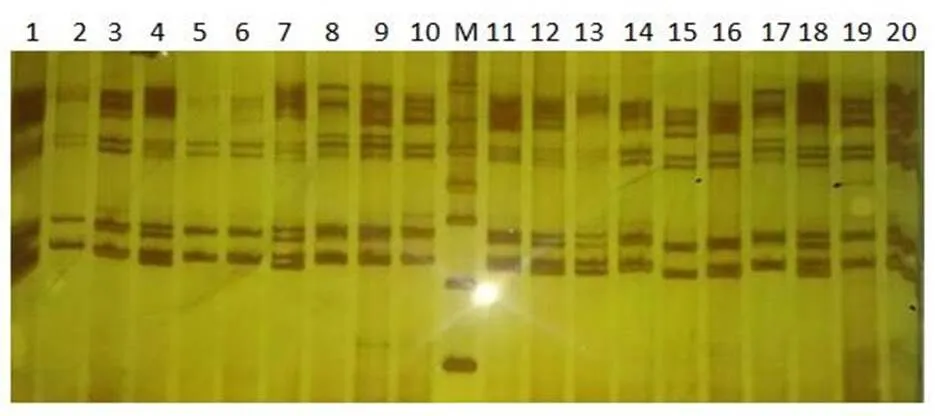
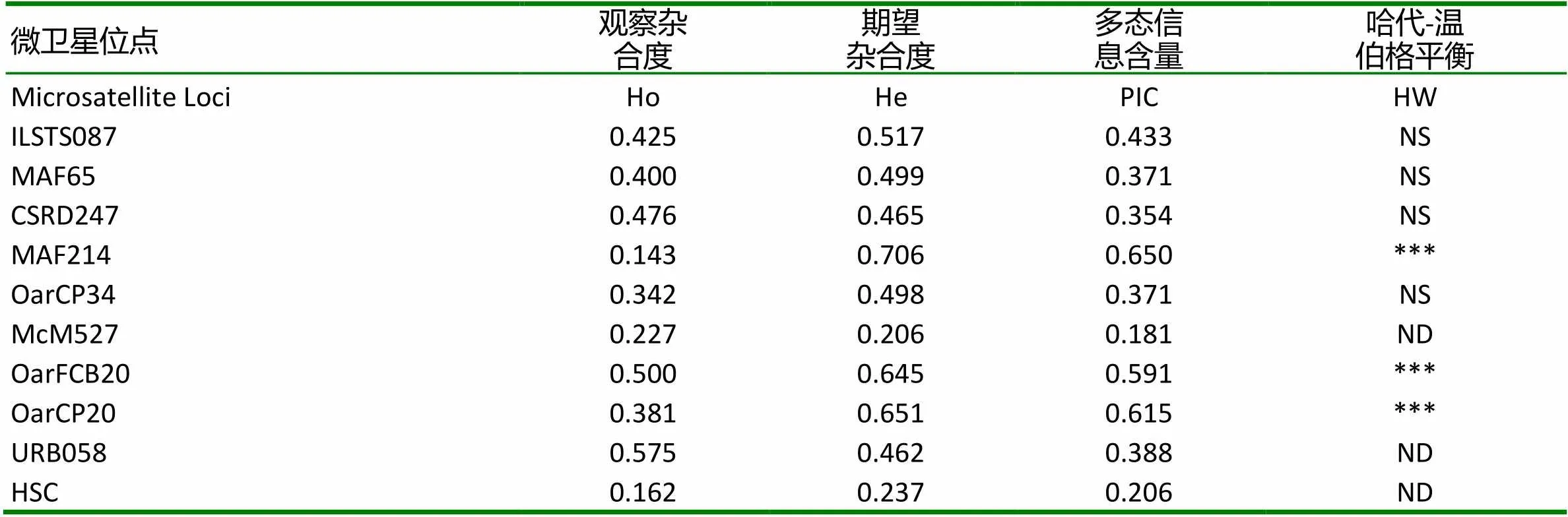
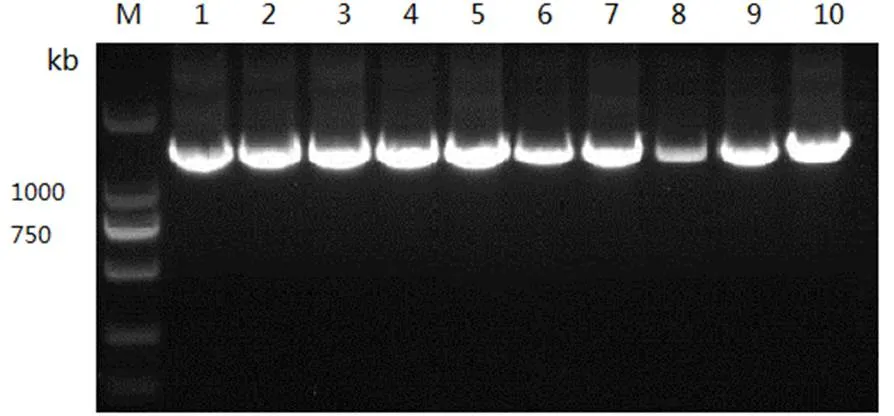
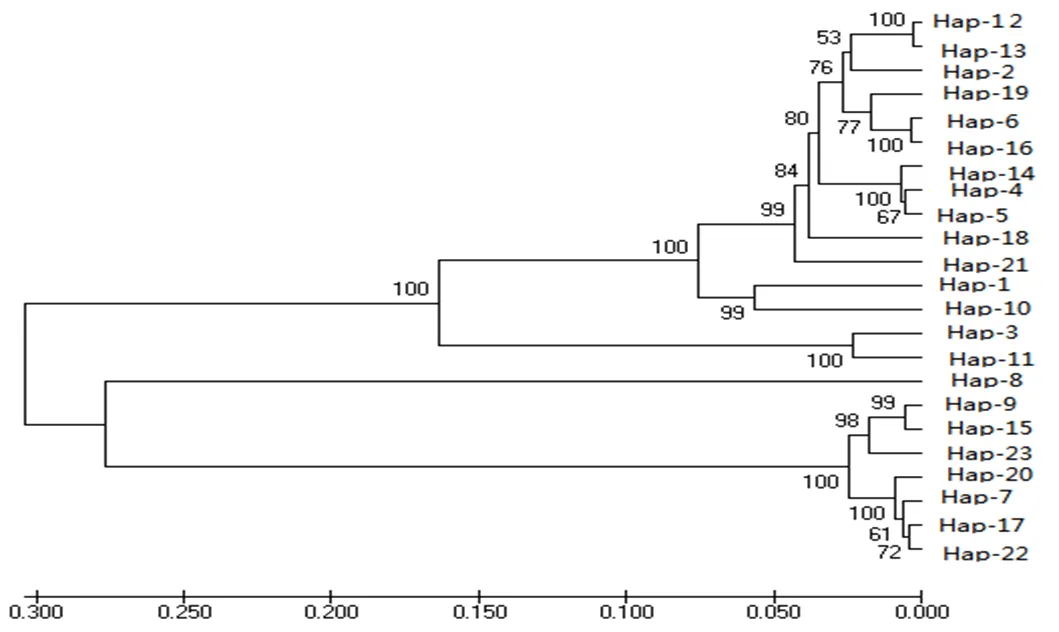
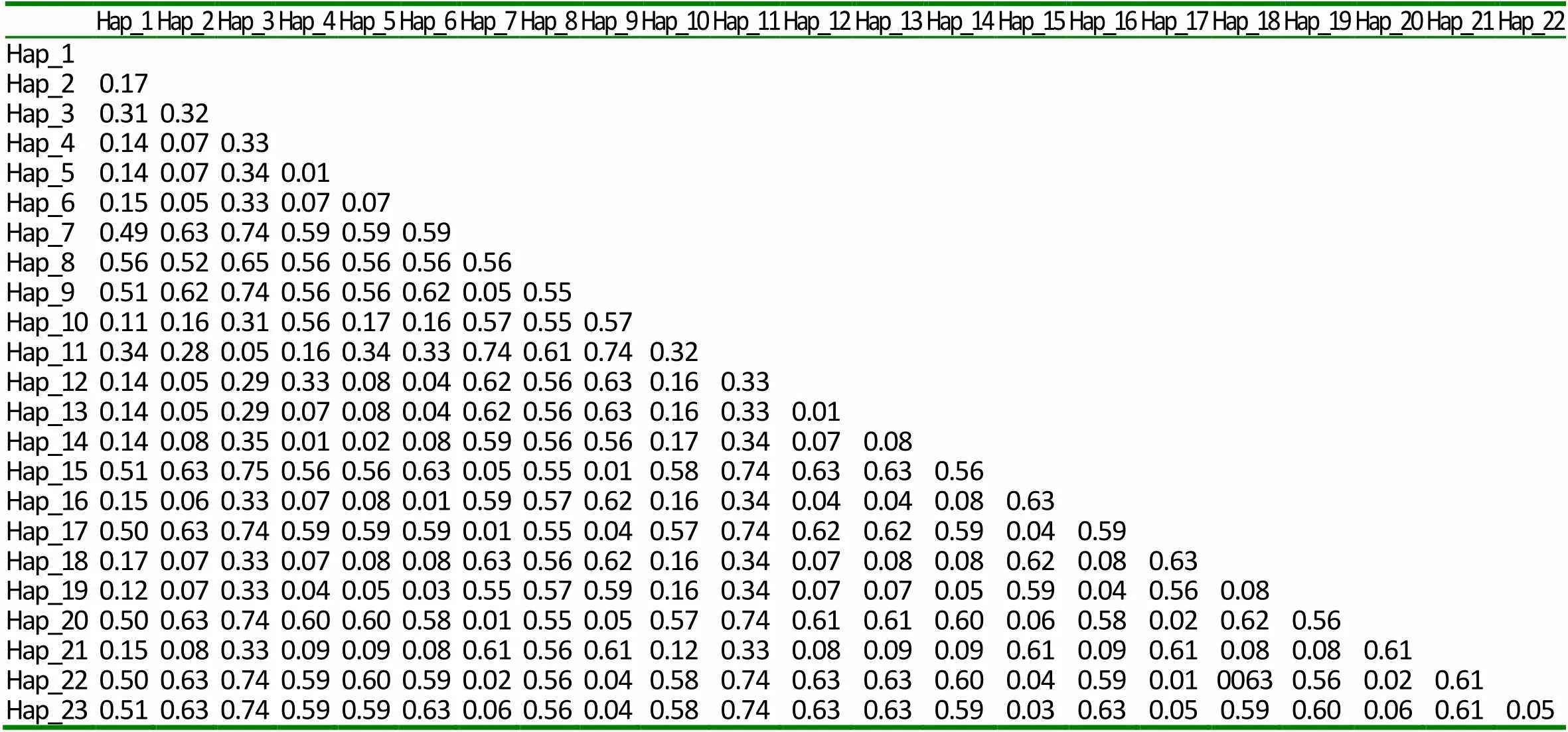
本研究對42只大尾寒羊的10個微衛星標記進行了遺傳多樣性分析,并對其中38只個體的線粒體DNA D-loop區的進行了測序和多態性分析。結果表明,10個微衛星位點共發現38個等位基因,平均觀察雜合度為0.3631,平均PIC值為0.416,FST、FIS和FIT平均值分別為0.0842、0.1784和0.2466,且該群體的HO 大尾寒羊 微衛星 線粒體D-loop區 遺傳多樣性 大尾寒羊是我國華北地方品種羊,公母羊的尾都過飛節,長者可接地或拖及地面,尾尖向上翻卷,形成明顯尾溝,屬大脂尾羊[1]。作為我國優良綿羊品種,其遺傳價值不容低估。由于保護開發投入不足,加之與其他品種的盲目雜交,導致大尾寒羊在部分地區瀕臨滅絕。妥善保存和利用大尾寒羊基因庫,制訂合理的保種方案,擴大其存欄數量已成為我省畜牧業生產中迫在眉捷的一項戰略任務[2, 3]。 微衛星DNA是可變數目串狀重復(VNTR)的一種[4]。微衛星具有多態信息含量豐富、雜合度高、共顯性遺傳、等位基因數目多、在畜群基因組DNA中分布廣泛等優點[5, 6]。線粒體DNA (mitochondrial DNA, mtDNA)是存在于線粒體基質中的、獨立于細胞核染色體外的基因組,與核基因相比,mtDNA 具有結構簡單、以母性方式遺傳、進化速度快和極少發生重組等特點[7-10]。其中D-loop區的進化速度最快,變異較大,適用于進行亞科內屬、種間的系統學研究[11, 12]。 本研究分析了微衛星和線粒體D-loop區的多態性,旨在為大尾寒羊的種質資源保護提供理論依據。 42只大尾寒羊(包括27只母羊和15只公羊)于2016年3月31日采自山東省臨清市大尾寒羊保種場,每只羊頸靜脈采血15ml于自帶肝素鈉的采血管中,保存在-20℃冰箱中以備提取基因組。 1.2.1 微衛星DNA的多態性分析 選取的10個微衛星來源于GeneBank,由生工生物工程(上海)股份有限公司合成。PCR反應總體系為25μl,包括10×Ex Taq Buffer 2.5μl,d NTPs 2μl,上下游引物(1μM)各0.5μl,基因組 DNA(50~100ng)2μl,Ex Taq 0.25μl,滅菌ddH2O 17.25μl。PCR反應條件為95℃5min;94℃30s,50~60℃30s,72℃ 30s,38個循環;72℃延伸10min。1%的瓊脂糖凝膠電泳檢測。選擇12%的非變性聚丙烯酰氨凝膠進行檢測。預電泳結束后,取5μl PCR產物與3μl 6× Loading Buffer混勻,上樣,160~180V電壓,電泳12~16h,硝酸銀染色,拍照。 1.2.2 線粒體DNA D-loop區的擴增與測序 根據NCBI數據庫提交的綿羊線粒體基因組序列(KR868678)設計擴增大尾寒羊線粒體D-loop區部分序列的特異引物。上下游引物序列分別為F:CCACTATCAACACCCAA,R:CGAAGGGCGT TACTCACC。引物由生工生物工程(上海)股份有限公司合成。 1.2.3 分型分析與序列分析 利用Cervus3.0計算大尾寒羊群體10個微衛星位點的觀察雜合度、多態信息含量(FIC)、哈代—溫伯格平衡等,對大尾寒羊多樣性進行評估。利用FSTAT3.9.2計算大尾寒羊總群體的近交系數(FIT)、不用亞群間的遺傳分化程度(FST)、群內近交系數(FIS)等遺傳多樣性參數。測得的線粒體D-loop區序列用DNA MAN6.0軟件進行整理對比,并進行人工校對。用DNAsp5.0軟件統計單倍型及變異位點、計算單倍型多樣性( Hd) 及核苷酸多樣度(Pi)等,評價群體遺傳多樣性水平。采用MEGA5.0軟件計算單倍型間的遺傳距離,并構建UPGMA分子系統樹。 表1 綿羊微衛星標記引物及退火溫度 2.1.1 微衛星位點的PCR擴增及多態性檢測 擴增結果顯示共有38個等位基因,平均每個位點有3.8個,其中在OarCP20位點檢測到的等位基因數最多(8個),在CSRD247、OarCP34、McM527和HSC四個位點檢測到的等位基因數最少(2個)。CSRD247位點的基因分析的結果如圖1所示。 圖1 微衛星CSRD247的PCR擴增產物PAGE膠圖 2.1.2 微衛星座位的遺傳參數 (1)所有微衛星座位上有效等位基因占85.11%,各位點平均有效等位基因數范圍在0.3306~0.9151之間,與有效等位基因數相差較大,表明等位基因分布不均勻,可能與人工選育有關。(2)我們選了檢測到的3個高度多態性位點和5個中度多態性位點來分析遺傳多樣性。這在這8個位點中有37.5%的位點偏離了哈代-溫伯格平衡,6個位點的期望雜合度都超過觀察雜合度(HO 表2 微衛星位點的觀察雜合度、期望雜合度、多態信息含量和哈代-溫伯格平衡 注:***為0.1%顯著性水平偏離哈代-溫伯格平衡,**為1%顯著性水平偏離哈代-溫伯格平衡,*為5%顯著性水平偏離哈代-溫伯格平衡,NS為不顯著偏離哈代-溫伯格平衡;ND 沒有做H-W檢測 FST值代表整個群體不同亞群間的遺傳分化程度,在8個所選的位點中,FST值在0.015到0.278之間,平均值為0.0842;群內近交系數(FIS)顯示大部分位點的近交系數為正值,平均值為0.1784;FIT值用來表示總群體的近交系數,其平均值為0.2466(表3)。 表3 群體的遺傳多樣性參數 2.2.1 mtDNA D-loop區序列擴增 根據普通綿羊線粒體DNA序列設計的引物在大尾寒羊基因組中得到較好的擴增(圖2)。PCR產物片段單一,擴增片段大小與預期的產物片段大小一致,可用于后續的測序分析。 圖2 大尾寒羊mtDNA D-loop區的擴增結果 2.2.2 mt DNA D-loop序列的遺傳多樣性 測序分析了其中38只D-loop區擴增結果較為理想的個體。結果顯示38個個體分別屬于23種單倍型(表4)。D-loop區堿基組成A+T的平均含量(62.0%)明顯高于G+C的平均含量(34.6%),轉換/顛換值為2.90,遺傳多樣性指數如表4所示。 表4 大尾寒羊mt DNA D-loop部分序列的遺傳多樣性指數 應用MEGA軟件,根據線粒體D-loop序列計算了23個單倍型之間的遺傳距離,單倍型的平均遺傳距離為0.37,其中有8組遺傳距離≤0.01,單倍型9、11的遺傳距離最大,為20.294。單倍型12、13為最小遺傳距離0.0055,絕大部分的遺傳距離在0~2之間(表5)。通過軟件構建了大尾寒羊單倍型間的UPGMA分子系統樹(Bootstrap檢驗重復為1000次)。由圖3可以看出,23種單倍型最終聚為3大分支。單倍型8與其他單倍型的同源性都有較遠距離。 圖3 大尾寒羊群體的UPGMA分子系統樹 表5 大尾寒羊mt DNA D-loop部分序列單倍型間的遺傳距離 (1)利用微衛星標記來探究種群的遺傳多樣性時,雜合度能夠反映群體在多個基因座位上的變異,它是衡量群體遺傳變異的一個最適參數[3~ 13]。在本研究中,10個標記的平均觀察雜合度為0.3631,期望雜合度均在0.3以上的座位有8個,觀察雜合度在0.3以上的座位有7個,與Takezaki和Nei(1996)認為微衛星計算出的雜合度范圍為0.3~ 0.8基本吻合[14]。多態信息含量(PIC)是基因豐富度的一個指標,多態信息含量的高低表明了品種遺傳基礎的豐富多樣性,用多態信息含量來描述種群在微衛星位點上的變異程度,當PIC>0.5時,該位點為高度多態;0.5>PIC>0.25時,該位點中度多態;PIC<0.25時,為低度多態[15, 16]。在所選用的10個微衛星多態位點的多態信息含量中HSC為和McM527低度多態,MAF214、OarFCB20和OarCP20為高度多態,其余基因座的多態信息含量為中度多態,所有位點的平均PIC值為0.416,說明所選擇的微衛星基因座位的多態性中等。(2)由于上述10個微衛星位點不論是PIC值還是雜合度,變化范圍都很大,不利于對大尾寒羊群體的遺傳多樣性進行評估。又分析了FIT、FST和FIS3個遺傳參數。FST值代表整個群體不同亞群間的遺傳分化程度,取值范圍從0~1,其值越大,表明亞群間遺傳分化越明顯。一般評價標準是FST<0.05,代表群體分化較小;0.05 本實驗由于保種場內樣品數量有限,品種間親緣關系混雜,抽樣效率低,繼而利用線粒體DNA D-loop區部分序列進行了分析,明確種群內個體的親緣關系及遺傳多樣性。應用mtDNA測序技術來研究物種的遺傳多樣性時,通常用兩個重要指標來衡量一個群體mtDNA的遺傳變異程度,單倍型間的平均遺傳距離(P)和核苷酸多樣性(π)[7, 20]。一般來說大多數哺乳動物的P值都在0.01以上時被認為變異大[21]。本研究顯示,群體單倍型間的平均遺傳距離為0.37,大于0.01,這說明中大尾寒羊的遺傳變異較大。核苷酸多樣性(π)是指群體內兩個隨機個體mtDNA序列間平均每個位點的核苷酸差異數目。Nei等定義,給定群體內兩個隨機選取的mtDNA序列間π值越小,表明群體的遺傳多樣性越低。本研究中,中大尾寒羊群體mtD NAD-loop區的核苷酸多樣性(π)為0.02815,高于Lan等認為的低遺傳多樣性指標(π值在0.0015 ~0.0047)[23],但低于山東其他綿羊品種,小尾寒羊(0.0324)與洼地綿羊(0.03172)[24],就其分子進化樹來看,三個分支內部的遺傳距離相對較近,存在近交現象。 總的來說該大尾寒羊種群雖然遺傳多樣性相對較高,但群體內部存在一定程度的近交現象,在大尾寒羊育種工作中,應避免遺傳距離較近的家系間的交配,進一步加大大尾寒羊保護、開發和利用力度,有效維持大尾寒羊這一國家保護品種的基本數量與質量。 [1] 王俊海, 姜永紅, 肖延光等. 大尾寒羊品種資源現狀與保護利用對策[J]. 山東畜牧獸醫, 2010, 31(5): 60-61+63. [2] 吳詩, 姜勛平. 基于GIS的中國山羊品種資源管理系統研制[J]. 中國草食動物, 2011, 31(4): 71-75. [3] 吳詩, 姜勛平, 蔡忠亮. 基于GIS的中國山羊品種資源管理系統研制[J]. 中國草食動物, 2011, 31(4): 71-75. [4] Beck N R, Double M C, Cockburn A. Microsatellite Evolution at Two Hypervariable Loci Revealed by Extensive Avian Pedigrees[J]. Molecular Biology and Evolution, 2003, 20(1): 54-61. [5] Zeveren A V, Peelman L, Weghe A V D, et al. A genetic study of four Belgian pig populations by means of seven microsatellite loci[J]. Journal of Animal Breeding and Genetics, 1995, 112(1-6): 191-204. [6] Markowitz S D, Wang J Y, Myeroff L L, et al. Markowitz, S. et al. Inactivation of the type II TGF- receptor in colon cancer cells with microsatellite instability. Science 268, 1336-1338[J]. Science, 1995, 268(5215): 1336-1338. [7] 黃雪貞, 錢國英, 李彩燕. 中華鱉3個地理群體線粒體基因D-loop區遺傳多樣性分析[J]. 水產學報, 2012, 36(1): 17-24. [8] 李齊發, 李隱俠, 趙興波等. 牦牛線粒體DNA D-loop區序列測定及其在牛亞科中分類地位的研究[J]. 畜牧獸醫學報, 2008, 39(1): 1-6. [9] Cann R L, Brown W M, Wilson A C. Polymorphic sites and the mechanism of evolution in human mitochondrial DNA[J]. Genetics, 1984, 106(3): 479-499. [10] Glenn T C, Staton J L, Vu A T, et al. Low mitochondrial DNA variation among American alligators and a novel non-coding region in crocodilians[J]. Journal of Experimental Zoology, 2002, 294(4): 312-24. [11] Sankoff D, Bryant D, Deneault M, et al. Early eukaryote evolution based on mitochondrial gene order breakpoints[C]// International Conference on Computational Molecular Biology. ACM, 2000: 254-262. [12] Saccone C, Gissi C, Lanave C, et al. Evolution of the mitochondrial genetic system: an overview[J]. Gene, 2000, 261(1): 153-9. [13] 包文斌, 束婧婷, 許盛海等. 樣本量和性比對微衛星分析中群體遺傳多樣性指標的影響[J]. 中國畜牧雜志, 2007, 43(1): 6-9. [14] Takezaki N, Nei M. Genetic Distances and Recons-truction of Phylogenetic Trees From Microsatellite DNA[J]. Genetics, 1996, 144(1): 389-99. [15] Botstein D, White R L, Skolnick M, et al. Construction of a genetic linkage map in man using restriction fragment length polymorphisms.[J]. American Journal of Human Genetics, 1980, 32(3): 314. [16] 張愛玲, 馬月輝, 李宏濱等. 利用微衛星標記分析6個山羊品種遺傳多樣性[J]. 農業生物技術學報, 2006, 14(1): 38-44. [17] Balloux F, Lugon‐Moulin N. The estimation of population differentiation with microsatellite markers.[J]. Molecular Ecology, 2002, 11(2): 155-65. [18] 孫俊麗, 張冰, 潘天彪等. 基于微衛星位點研究7個廣西地方豬種的遺傳關系[J]. 西南農業學報, 2016, 29(6): 1475-1480. [19] 毛永江, 常洪, 楊章平等. 3個中國地方黃牛品種遺傳結構及其遺傳分化的研究[J]. 畜牧獸醫學報, 2007, 38(2): 125-132. [20] Neigel J E, Avise J C. Application of a random walk model to geographic distributions of animal mitochondrial DNA variation.[J]. 1994, 135(4): 1209-1220. [21] Lan H, Shi L. The origin and genetic differentiation of native breeds of pigs in southwest China: an approach from mitochondrial DNA polymorphism.[J]. Biochemical Genetics, 1993, 31(1): 51-60. [22] Neigel J E, Avise J C. Application of a random walk model to geographic distributions of animal mitochondrial DNA variation.[J]. 1994, 135(4):1209-1220. [23] Lan H, Shi L. The origin and genetic differentiation of native breeds of pigs in southwest China: an approach from mitochondrial DNA polymorphism.[J]. Biochemical Genetics, 1993, 31(1): 51-60. [24] 黃慶華, 張果平, 王金文等. 山東省地方綿羊品種的mtDNA D-loop序列多態性及系統進化研究[J]. 山東農業科學, 2012(5): 5-8. (2019–04–09) S813.3 A 1007-1733(2019)07-0001-051 材料和方法
1.1 試驗動物
1.2 方法

2 結果
2.1 微衛星DNA的多態性分析

2.2 mtDNA D-loop區多態性分析

2.3 單倍型之間的遺傳距離及分子系統樹
3 討論
3.1 微衛星DNA的多態性分析
3.2 mtDNA D-loop區多態性分析
