側柏母樹大小對種子特性和幼苗生長的影響
2019-07-13 07:45:14張金顥曹幫華毛培利山東英才學院山東濟南5004
種子 2019年6期
關鍵詞:生長
張金顥 曹幫華 毛培利(.山東英才學院 山東 濟南 5004;
2.山東農業大學林學院/黃河下游森林培育國家林業局重點實驗室/山東泰山森林生態系統國家定位觀測研究站, 山東 泰安 271018;3. 山東省林木種苗和花卉站, 山東 濟南 250014)
側柏(Platycladusorientalis)是我國重要鄉土樹種,具有適應性強、抗嚴寒、耐干旱瘠薄等特性,是我國荒山造林和水土保持的先鋒樹種。隨著側柏人工林的不斷發育,其天然更新逐漸受到了人們的關注,尤其是對于二十世紀五六十年代營造的林分。這是因為成功的天然更新對側柏人工林的穩定和生態功能持續發揮具有重要的意義。通過對側柏人工林林下幼苗分布的調查,認為郁閉度是影響側柏林下幼樹生存、生長的首要因子[1]。王斐和臧麗鵬認為,除了充足的光照,較高的土壤含水量也是側柏幼苗更新的重要限制因子[2]。除了更新的環境因子,種子質量也是側柏人工林天然更新的關鍵因子。張夢穎等發現,隨海拔增加,側柏人工林種子活力先增加再降低,在海拔300~500 m最高,種子發芽率超過60%[3]。但是關于母樹大小對側柏種子質量及幼苗生長的影響目前還沒有系統研究。
林木種子的質量直接影響著人工林的生長發育[4]和森林天然更新能否成功[5]。樹木個體大小對種子質量有著重要的影響。隨著母樹胸徑的增加,黑松種子的發芽率逐漸增加[6]。太白紅杉種子的千粒重隨母樹基徑的增加逐漸增加,但南坡是先增加再降低[7]。對于水青樹[8]、胖大海[9]、四合木[10]、Pinusechinata[11]和Sorbustorminalis[12]則是隨著母樹胸徑或基徑的增加,種子質量先增加再降低,在中間胸徑達到最大值。也有研究表明,香果樹[13]、Piceamariana[14]的種子發芽率不受母樹胸徑的影響。以上研究結果表明,隨母樹個體大小的增加,林木種子質量有逐漸增加、先增加再降低和變化不顯著3種情況。對不同胸徑黑松母樹種子特性的研究發現,胸徑大的母樹不僅種子質量好,而且形成的幼苗有著高的生物量和生長速率,這對早期幼苗建立有著重要意義[6]。
側柏林是泰山主要人工林之一,也是主要的風景林之一。大部分是上二十世紀五十年代營造,以其為主的林分總面積約為1 080 hm2,主要分布在海拔800 m以下的山坡上。由于是風景林,該林分的改造要求嚴格,極少進行撫育更新,造成林分結構不合理,生態穩定性和功能的發揮受到一定的影響。天然更新對林分結構改造效果好、成本低。本文以泰山20世紀50年代營造的側柏純林中不同胸徑母樹的種子為對象,研究種子萌發和幼苗生長動態,闡明母樹大小對幼苗建立階段的影響機制,為側柏人工林的天然更新管理提供理論依據。
1 材料與方法
1.1 研究區域概況
泰山位于山東省的中部,橫亙于濟南、泰安兩市之間。研究區屬于暖溫帶大陸性季風氣候區,多年平均氣溫為12.6 ℃,無霜期196 d,≥10 ℃的活動積溫3 821 ℃,多年平均降水量758 mm,降水集中在6—9月。土壤類型主要是棕壤土,pH值為6.0左右,土層厚度15~60 cm。泰山植被大部分為人工林,森林總面積9 490 hm2左右,森林覆蓋率81.5%。主要的樹種有側柏(P.orientalis)、麻櫟(Quercusacutissima)、刺槐(Robiniapseudoacacia)、黑松(Pinusthunbergii)、赤松(Pinusdensiflora)、油松(Pinustabuliformis)等。
1.2 研究方法
1.2.1種子采集
2017年10月,根據調查結果[3],在海拔為200 m的泰山普照寺周邊的側柏林內采集了3個胸徑徑級的側柏母樹種子,每個徑級采集母樹5~10株。徑級Ⅰ的胸徑為9.2~11.8 cm,徑級Ⅱ為14.2~16 cm,徑級Ⅲ為18.8~21.2 cm。通過自然風干待球果開裂后收集飽滿種子用于實驗。每個徑級種子用百粒法測定千粒重,并分別選擇30粒種子用游標卡尺測定最大長度、最大寬度。
1.2.2種子萌發試驗
種子萌發試驗在山東農業大學林學試驗站內進行。2018年4月24日,將取自側柏林內的土壤過篩、去除雜質,放入直徑20 cm、高20 cm的白色塑料盆內至預先標定的深度。澆水至盆底有水流出后,每個花盆擺20粒側柏種子于表面,再覆蓋0.5 cm土于表面蓋住種子。按照采種的3個母樹徑級水平(Ⅰ、Ⅱ、Ⅲ),每個水平9個重復,共計27盆,在室外進行萌發實驗。種子突破表土后視作萌發。種子萌發后,每天記錄萌發的種子數。試驗期間,定時澆水,使土壤保持濕潤。6月13日,萌發試驗結束觀察。統計發芽率(Germination rate,Gr)、發芽指數(Germination index,Gi)和平均發芽時間(Mean germination time,MGT)。
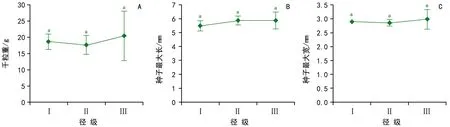
圖1 不同徑級側柏母樹種子大小特征
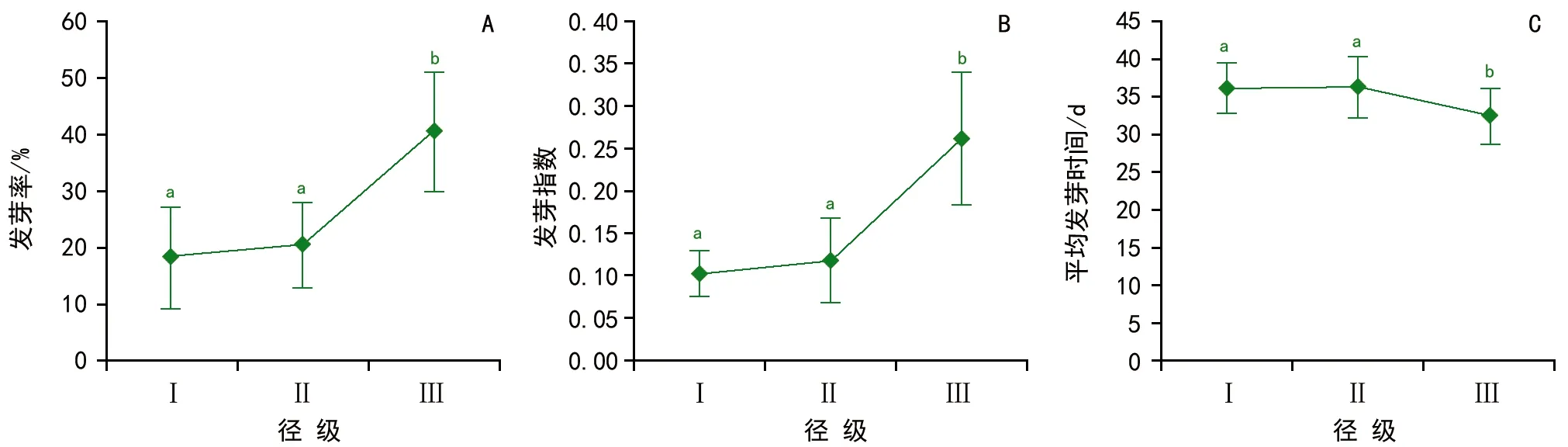
圖2 母樹徑級對側柏種子萌發的影響
Gr(%)=G1/N×100%
式中:G1為萌發數,N為供試種子總數。
MGT=∑(D×n) /∑n
式中:n為對應各日正常發芽的粒數,D為種子置床起的天數。
Gi=∑(Gt/Dt)
式中:Gt為各日凈發芽數;Dt為相應發芽日。
1.2.3幼苗生長試驗
種子萌發試驗結束后,每盆選取大小一致的幼苗3株,分別于7月12日、8月23日、9月23日和10月26日測定幼苗的高度和地徑。試驗期間,定期澆水。2018年10月26日,幼苗生長試驗結束,取出幼苗,并用清水沖洗干凈,分為地上部和根系。用HP Scanjet 8200掃描儀掃描根系,再用Delta-T Area Meter Type AMB 2根系參數分析軟件分析掃描的圖片獲取根系長度和根系表面積。然后將地上部和根系在120 ℃的烘箱中殺青30 min,再在80 ℃下烘干直至恒重,稱量干重并記錄。計算幼苗干重(Biomass,地上部重+根重)、根冠比(Root/shoot ratio,根重/生物量)、比根長(Specific root length,SRL,根長度/根系干重)和比根表面積(Specific root area,SRA,根表面積/根系干重)。計算相對苗高生長速率(Relative height growth rate, RGRH)和相對地徑生長速率(Relative ground diameter growth rate, RGRD)。
RGRH= (lnH2-lnH1)/(T2-T1)
RGRD= (lnD2-lnD1)/(T2-T1)
式中:H為幼苗高度,D為幼苗地徑,T1代表試驗開始時間,T2代表試驗結束時間。
1.3 統計分析
所有數據采用SPSS 17.0軟件(SPSS,Chicago,IL,USA)進行分析。采用單因子方差分析法(One-Way ANOVA)檢驗了不同母樹徑級之間在千粒重、種子最大長、種子最大寬、發芽率、發芽指數、平均發芽時間、幼苗干重、根冠比、比根長和比根表面積。用雙因子方差分析法(Two-Way ANOVA)比較了幼苗苗高、地徑生長指標。若差異顯著,再用最小顯著差異法(LSD)來進行多重比較。顯著性差異檢驗水平是p<0.05。
2 結果與分析
2.1 不同徑級母樹種子大小
隨母樹徑級的增加,千粒重、種子最大長和種子最大寬逐漸增加(圖1)。對于千粒重,徑級Ⅰ的種子變化幅度為21.31%,徑級Ⅱ的為30.49%,徑級Ⅲ的為49.49%。而方差分析結果表明,千粒重(F=0.30,p=0.75)、種子最大長(F=0.76,p=0.50)和種子最大寬(F=0.36,p=0.71)在不同徑級之間差異均不顯著。
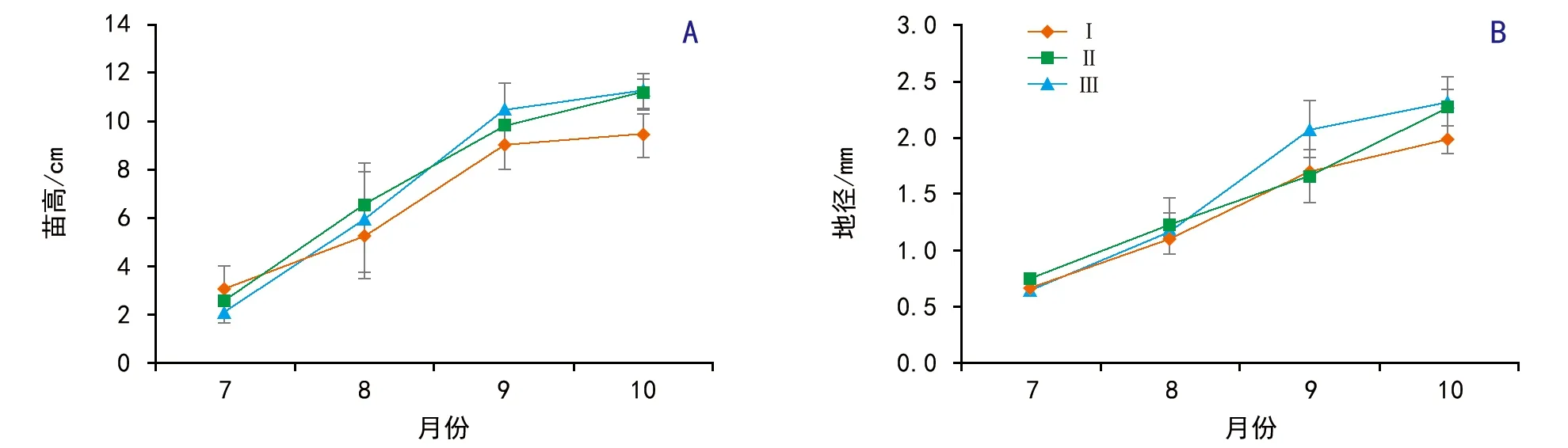
圖3 母樹徑級對側柏幼苗生長的影響

圖4 母樹徑級對側柏幼苗相對生長速率的影響
2.2 不同徑級母樹種子的萌發
隨母樹徑級的增加,發芽率、發芽指數逐漸增加,平均發芽時間逐漸降低(圖2)。方差分析結果表明,發芽率(F=9.49,p<0.01)和發芽指數(F=11.76,p<0.01)在不同徑級之間差異極顯著,平均發芽時間(F=3.58,p<0.05)在不同徑級之間差異顯著。發芽率和發芽指數都是在徑級Ⅰ和徑級Ⅱ之間差異不顯著(p>0.05),極顯著小于徑級Ⅲ(p<0.01);對于平均發芽時間,則是在徑級Ⅰ和徑級Ⅱ之間差異不顯著(p>0.05),顯著大于徑級Ⅲ(p<0.05)。
2.3 不同徑級母樹幼苗的生長動態
2.3.1苗高地徑生長動態
隨生長月份的增加,不同徑級側柏的幼苗苗高和地徑逐漸增加(圖3)。方差分析結果表明,母樹徑級大小對苗高影響不顯著(F=2.50,p=0.10),對地徑影響顯著(F=4.10,p<0.05);生長月份對苗高(F=128.42,p<0.01)和地徑(F=150.07,p<0.01)都影響極顯著;但交互作用在苗高(F=1.24,p=0.31)和地徑(F=2.03,p=0.0.09)上都影響不顯著。不同徑級之間,徑級Ⅰ的地徑極顯著低于徑級Ⅲ(p<0.01),徑級Ⅰ與徑級Ⅱ、徑級Ⅱ與徑級Ⅲ之間均差異不顯著(p>0.05)。在生長月份之間,對于苗高是7月<8月<9月和10月(p<0.01),而9月和10月之間差異不顯著(p>0.05);對于地徑則是7月<8月<9月<10月(p<0.01)。
2.3.2相對苗高和地徑生長速率動態
隨生長月份的增加,徑級Ⅰ和徑級Ⅲ的相對苗高生長速率與相對地徑生長速率都是先增加再降低,而徑級Ⅱ的則是逐漸降低(圖4)。方差分析結果表明,母樹徑級大小對相對苗高生長速率影響顯著(F=3.90,p<0.05),對相對地徑生長速率影響不顯著(F=0.31,p=0.74);生長月份對相對高度生長速率(F=82.22,p<0.01)和相對地徑生長速率(F=9.28,p<0.01)都影響極顯著;二者交互作用對相對苗高生長速率影響極顯著(F=6.54,p<0.01),對相對地徑生長速率影響不顯著(F=2.45,p=0.70)。不同徑級之間,徑級Ⅰ的相對苗高生長速率顯著低于徑級Ⅲ(p<0.05),徑級Ⅰ與徑級Ⅱ、徑級Ⅱ與徑級Ⅲ之間均差異不顯著(p>0.05)。生長月份之間,相對苗高生長速率和相對地徑生長速率都是7—8月與8—9月之間差異不顯著(p>0.05),二者極顯著高于9—10月(p<0.01)。
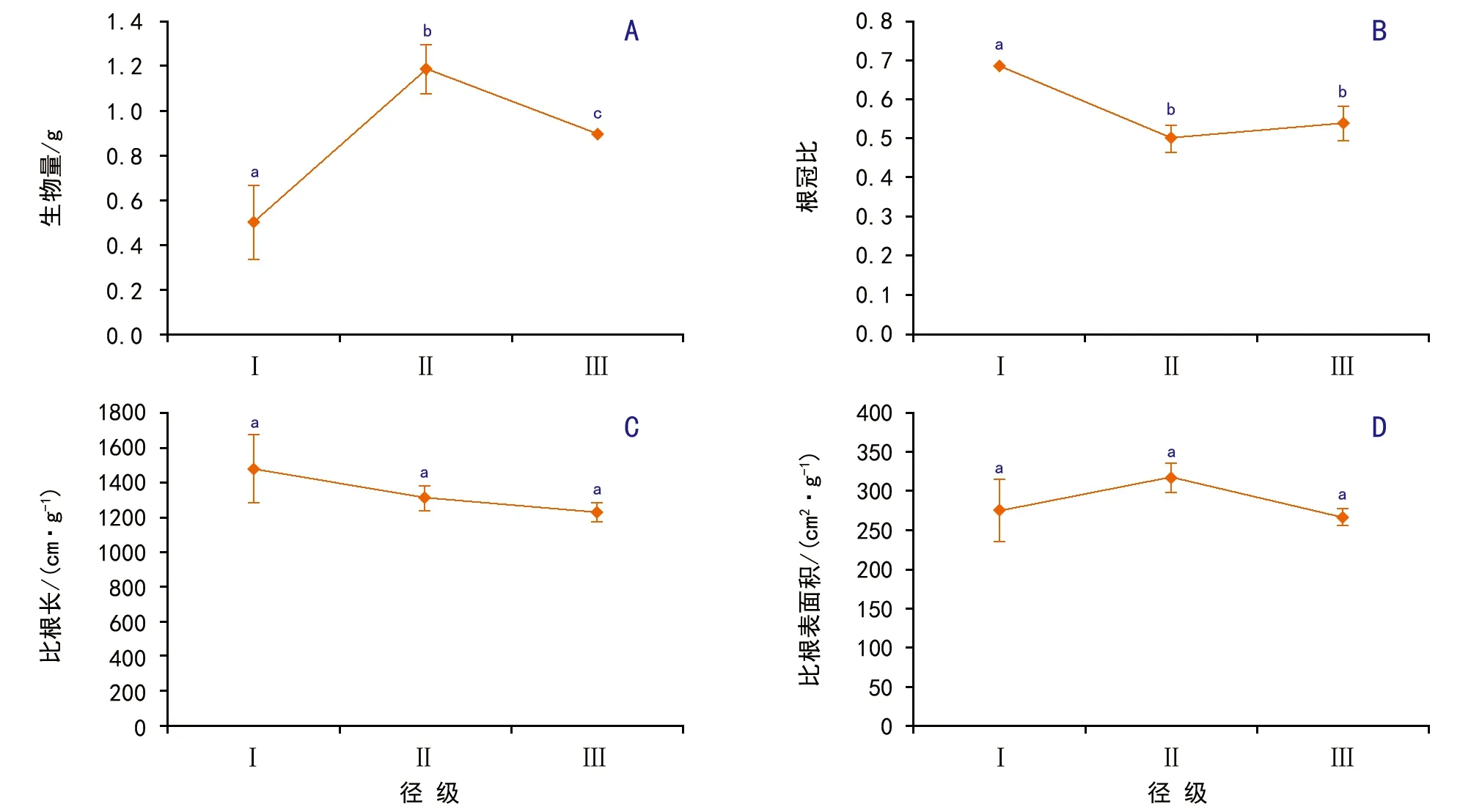
圖5 母樹徑級對側柏幼苗的生物量及其分配的影響
2.4 不同徑級母樹幼苗的生物量及分配
隨母樹徑級的增加,幼苗干重和比根表面積先增加再降低,根冠比和比根長則是逐漸降低(圖5)。方差分析結果表明,幼苗干重(F=27.00,p<0.01)和根冠比(F=27.30,p<0.01)在不同徑級之間差異極顯著,而比根長(F=3.20,p=0.11)差異不顯著。對于幼苗干重是徑級Ⅰ<徑級Ⅲ<徑級Ⅱ(p<0.05);對于根冠比,徑級Ⅰ極顯著大于徑級Ⅱ與徑級Ⅲ(p<0.01),而徑級Ⅱ與徑級Ⅲ差異不顯著(p>0.05)。
3 結論與討論
種子大小是反映其質量的重要特征。本研究發現,隨母樹徑級的增加,側柏種子千粒重逐漸增加,但變化不顯著。馬紹賓等也發現,多年生草本植物桃兒七在同一種群內單粒種子重量變化不大[15]。但水青樹種子的干重[8]和四合木果實的重量[10]則隨母樹胸徑(或基徑)增加先增加再降低。太白紅杉種子千粒重隨隨母樹胸徑增加而增加,王孝安等認為,與植株用于生殖的投資增多有關,而隨后的降低則是因為植株變大引起樹木間相互干擾加大導致生殖投資減少造成的[7]。本文中側柏種子的千粒重在不同徑級母樹之間變異很大,在徑級Ⅲ達到了49.49%,而種子萌發和幼苗生長中依然是徑級Ⅲ的最高。呂文芳等發現,種子千粒重并不能代表種子的發芽能力和隨后的幼苗生長水平[16]。而李懷春等也發現,水青樹種子千粒重和發芽能力最大在母樹胸徑30~40 cm最大,而種子的長、寬、厚在母樹胸徑40~50 cm最大,二者表現不一致[8]。種子大小的變化可能主要與母樹生長的環境[17]和生長狀況[18]有關。而種子質量可能受到其它因素的影響,如母樹遺傳因素,所以有時種子大小與種子質量不一定完全一致。
種子萌發是植物開始天然更新的關鍵階段之一。本文發現,隨母樹徑級的增加,側柏種子的發芽率、發芽指數逐漸增加,平均發芽時間逐漸降低,表明種子質量逐漸提升,這與Mao等[6]對黑松種子的研究結果一致。發芽時間的增加會使種子儲藏養分過多消耗,導致不能萌發或幼苗細弱,影響幼苗成活率[19]。因此,側柏大徑級的母樹形成的種子在萌發階段具有明顯的優勢。但水青樹[8]、胖大海[9]、Pinusechinata[11]和Sorbustorminalis[12]的種子發芽率則是在中間胸徑的母樹達到最大值。這可能與本文側柏母樹胸徑的范圍(9.2~21.2 cm)較小有關。對于林木種子發芽率的降低可能與母樹對生殖分配減少有關[8-9]。香果樹[13]、Piceamariana[14]的種子發芽率不受母樹胸徑的影響。侯聰等認為,不同水平密度疏伐的紅松母樹林內種子生活力沒有顯著差異,與遺傳控制有關[20]。因此,樹木在種子萌發上的差異除了與生長環境密切相關,也與母樹相關的遺傳因素有關。
生長是植物對外界不同脅迫的綜合體現,反映了植物的適應能力[21]。本研究發現,不同徑級側柏的幼苗中,徑級Ⅰ的地徑、相對苗高生長速率和幼苗干重都顯著低于徑級Ⅲ。Mao等也發現,母樹徑級最大的黑松種子形成的幼苗有著最大的生物量和相對生長速率最大[6]。高的相對苗高生長速率有利于幼苗快速高生長,獲得更多的光資源,提高與周邊植物競爭的能力[22]。側柏母樹徑級Ⅰ的根冠比極顯著大于徑級Ⅱ與徑級Ⅲ。減少對地上部的投入,尤其是對葉片的投入,會降低植物的光合能力、生長速率[23]及在林下的光截獲能力[24]。側柏是喜光植物,快速的高生長有利于其在林窗或林緣快速生長進入冠層。因此,生長速率低、根冠比高不利于側柏徑級Ⅰ種子形成的幼苗在林下的建立。
母樹大小對種子的質量有著顯著的影響。本文研究隨著徑級的增加,側柏種子的質量逐漸提升,發芽率和發芽指數逐漸增加,平均發芽時間逐漸降低,形成幼苗的生物量和生長速率也是大徑級母樹的最高,這對側柏林下的幼苗建立具有重要意義。雖然側柏不同徑級母樹之間種子的大小變異較大,但并沒有影響種子的萌發和幼苗生長,說明種子的大小不能完全反映種子質量。這可能與種子在形成過程中受到母樹生長環境、自身營養狀況、遺傳因素等多方面的共同影響有關。因此,在今后的研究中,深入開展側柏母樹生長環境和本身的生長狀況對種子形成的影響,有助于準確判斷種子質量,有利于提出合理有效的人工促進天然更新措施。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
