沙地樟子松天然林南緣分布區林木競爭、空間格局及其更新特征
2019-07-05 10:59:48潘磊磊KWONSEMYUNG劉艷書楊曉暉朱媛君時忠杰
生態學報 2019年10期
潘磊磊,KWON SEMYUNG ,劉艷書,張 曉,楊曉暉,山 丹,朱媛君,時忠杰,*
1 中國林業科學研究院荒漠化研究所,北京 100091 2 北京林業大學水土保持學院,北京 100083
林木的生長與更新受到許多生物和非生物因素的影響,其中,競爭和空間格局是影響林木生長、林分結構及其動態的重要因素,在種群更新與演替中發揮著重要的作用,是生態學和林學研究的重要問題之一[1- 2]。競爭普遍存在于森林生態系統中,鄰近的林木個體之間通過競爭獲取更多的光照、水分和養分等資源,從而影響鄰近個體對資源的利用[3-5]。而林木空間格局作為種群分布的外在表現形式,是其分布格局及其屬性在空間上的排列方式[6],表達了林木間樹種、大小、分布等空間關系,決定了樹木之間的競爭優勢及其空間生態位,是種群結構、動態和生態學過程、種內種間關系及外界環境相互作用的結果[7-8]。
近年來,許多學者對于林木的競爭[9-10]、空間格局[11-12]和天然更新[13]等進行了較為深入的研究。在林木競爭方面主要集中于種內競爭[14-15]、種間競爭[16-17]、種間差異[14]及其資源利用特性[18]、外界干擾的影響[19]等,而林木空間格局研究主要分析種群生物學特性[20]、種內和種間關系[21]以及植物與環境之間的相互作用和空間分布狀態[22-23]等。而林分的更新可能受林木的競爭與空間格局等因素的影響,但對于林木競爭及空間格局與群落更新的關系研究卻甚少,制約了對群落更新機制的認識。
我國的樟子松天然林主要分布在大興安嶺山地北部和呼倫貝爾森林草原過渡帶的沙地上,其地理分布的最南緣位于呼倫貝爾沙地的南端。由于樟子松具有耐寒、耐旱、根系發達及不苛求土壤水分等特征,也被廣泛應用到我國北方沙地造林中,對于防風固沙、農田防護等方面發揮著重要的作用。國內對于沙地樟子松的空間格局、競爭與更新等方面已有較多研究,特別是火干擾對競爭、格局與更新的影響方面[19,24- 25],如喻泓等[19]發現樟子松的競爭壓力主要來源于種內,幼樹多呈聚集分布特征,大樹多呈隨機分布特征,而火干擾是樟子松林分天然更新的主要驅動因子之一[24]。但對于天然沙地樟子松林純林林木競爭、空間格局與更新的關系研究并未深入。為此,本研究針對天然沙地樟子松地理分布最南緣區的純林,旨在充分認識林木之間的競爭關系、空間格局及其天然更新特征,厘清林木競爭與空間格局對天然更新的影響機制,為樟子松天然林的經營與保護及人工林的營造與管理等提供科學的指導。
1 研究區概況
研究區位于呼倫貝爾沙地東南端、新巴爾虎左旗罕達蓋蘇木中國蒙古邊界附近(47°38′—47°39′N,119°09′—119°12′E)。呼倫貝爾沙地位于我國東北西部,東西長約270 km,南北寬約170 km,面積約1萬km2,本區屬中溫帶半干旱大陸性氣候,夏季溫暖,冬季嚴寒、日照充足,晝夜溫差大,年平均氣溫0.2 ℃,年降水量280—400 mm左右,多集中在夏秋季,年蒸發量1400—1900 mm,無霜期90—100 d,積雪期長達140 d左右[26]。地貌類型以固定沙丘和半固定沙丘為主,土壤以風沙土為主[27]。植被以森林和草原為主,該區是沙地樟子松(Pinussylvestrisvar.mongolica)種群在我國地理分布的最南緣。本區的樟子松林多以純林為主,伴生種有白樺(Betulaplatyphylla)、榛子(Corylusheterophylla)等,以及線葉菊(Filifoliumibiricum)、大針茅(Stipagrandis)、羊草(Leymuschinensis)、日陰菅(Carexpediformis)、地榆(Sanguisorbaofficinalis)等雜類草[28]。

表1 樣地基本情況
2 研究方法
2.1 數據獲取
2017年7—9月,在研究區選取人為干擾較小的沙地樟子松天然林,建立2個密度(DBH>5 cm林木株數/hm2)相似的50 m×50 m的樣地,并每木檢尺,對所有樹高2 m以上的樹木統一編號,測定其胸徑、樹高、枝下高、冠幅(按東、南、西、北4個方位)等林分指標,而樹高小于2 m的樹木則測定其基徑,并利用全站儀(NTS 340,南方測繪儀器有限公司)測定所有林木空間位置坐標。樣地基本信息如表1,調查樣地內的林木空間坐標信息見圖1。
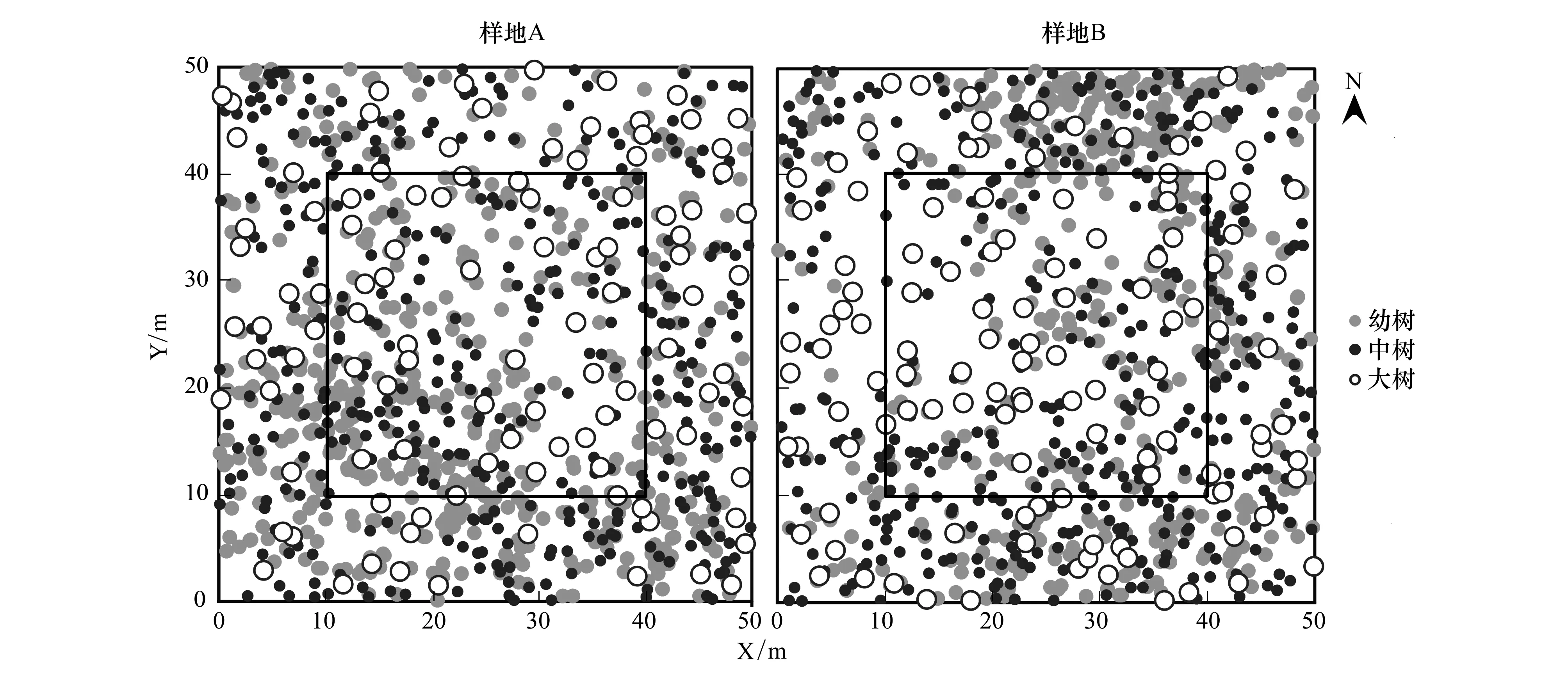
圖1 樣地林木分布Fig.1 Maps of trees distribution in sampling sites點代表樹木,點的大小代表樹木胸徑,實心灰色、實心黑色、空心圓分別代表幼樹、中樹和大樹;樣地內圈黑色實方框指代為消除邊緣效應,最終用于林分結構、競爭和更新分析的樣方大小
2.2 林分結構與對象木的選擇
將兩個樣地內對象木的胸徑從0 cm開始統計,以3 cm為間距,劃分為13個徑階:徑階I,DBH<3 cm;徑階II,3 cm≤DBH<6 cm;徑階III,6 cm≤DBH<9 cm;徑階IV,9 cm≤DBH<12 cm;徑階V,12 cm≤DBH<15 cm;徑階VI,15 cm≤DBH<18 cm;徑階VII,18 cm≤DBH<21 cm;徑階VIII,21 cm≤DBH<24 cm;徑階IX,24 cm≤DBH<27 cm;徑階X,27 cm≤DBH<30 cm;徑階XI,30 cm≤DBH<33 cm;徑階XII,33 cm≤DBH<36 cm;徑階XIII,36 cm≤DBH<39 cm,統計各個徑階的植株數,繪制徑階分布圖。將對象木的樹高從0 cm開始,按2 m為一個高階,劃分為8個高階:高階I,H<2 m;高階II,2 m≤H<4 m;高階III,4 m≤H<6 m;高階IV,6 m≤H<8 m;高階V,8 m≤H<10 m;高階VI,10 m≤H<12 m;高階VII,12 m≤H<14 m,統計各個高階的植株數、繪制樹高分布圖。為了分析樟子松不同生長階段的空間分布格局特征,結合實地調查樟子松的生活史特征,將樟子松劃分為3個生長階段:幼樹,DBH<9 cm;中樹,9 cm≤DBH<18 cm;大樹,DBH>18 cm。為了消除邊緣效應,將樣地劃分為25個10 m×10 m的樣方(圖1),選擇中間9個樣方內的所有林木作為對象木開展林分結構、競爭和更新分析。
2.3 競爭分析
競爭是個體或群體之間對生存空間與資源利用的關系,目前對個體競爭強度的研究較多,并提出了許多競爭模型,包括與距離相關的和與距離無關的競爭指數[29-30],但以Hegyi[31]的競爭模型模擬效果最好,應用最廣泛[32-33]。因此,本研究采用了簡便易算的與距離相關的Hegyi的競爭指數,其公式為:
(1)
(2)
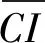
一般認為,當植株的樹冠或根系發生接觸或重疊時,才會發生競爭,所以對象木所受到的競爭壓力主要來自于其周圍一定距離內的競爭木,而當超出這一距離,其對對象木的競爭逐漸減弱或消失。為此,本文采用固定半徑法,選定A樣地35株和B樣地30株對象木,以2 m為起點,以1 m為樣圓半徑增量,分別計算2—20 m范圍內每個樣圓半徑對象木的平均競爭指數,做出樣圓半徑與平均競爭指數的散點圖,分析不同樣圓半徑與平均競爭指數的關系,隨著樣圓半徑的增加,當競爭強度變化不顯著時即確定為對象木的競爭范圍Lij[34]。
利用樟子松對象木的競爭強度與其胸徑進行回歸分析,其冪函數回歸擬合關系式為:即CI=a×D-b,式中CI為競爭指數,D為對象木的胸徑,a,b為模型參數。當P<0.05時,達到顯著水平;當P<0.01時,達到極顯著水平。
2.4 點格局分析
點格局相關理論是由Ripley[35]首先提出,后經Diggle[36]等人發展,已被廣泛應用于種群的空間分布格局以及種內、種間空間關聯性分析中。Ripley′sK(r)函數是一個積累分布函數,在應用過程中,常因為積累效應的存在影響到結果的準確性[37-38]。Ripley′ sK(r)函數如下:
式中,A是研究區(樣地)的面積,uij為2個點i和j之間的距離;Ir(uij)為指示函數,當uij≤r時,Ir(uij)=1,
當uij>r時,Ir(uij)=0;wij為權重值,用于邊緣校正。
成對關聯函數(pair correlation function)簡稱為g(r)函數來源于Ripley′sK函數,但它可以較好地排除K(r)函數引起的累積效應,具有更廣的適用性[39-40]。本研究選擇g(r)函數分析不同物種的空間分布特征以及空間關聯性。g(r)函數與Ripley′K(r)函數之間的關系為:
g(r)=(2πr)-1dK(r)/dr
式中,dK(r)是函數K(r)的微分,dr是半徑r的微分。
種間或種內的空間關聯可以運用雙變量點格局分析,即:
g12(r)=(2πr)-1dK12(r)/dr
該公式計算以關聯物種或群體1的每個個體為圓心,半徑為r的圓環區域內關聯物種或群體2的數量[41- 43]。
在單變量g(r)分析時,g(r)值落在上下包跡線之間為隨機分布,在上包跡線以上為顯著的聚集分布,在下包跡線以下為顯著均勻分布。雙變量g12(r)可以用物種種間以及同一物種的不同生長階段之間的空間關聯性,按照植物的生長發育規律,大樹會對中樹、幼樹的空間分布特征產生影響[38]。所以在進行雙變量分析時,保持大樹的位置不變,中樹和幼樹隨機變化;中樹位置保持不變,幼樹隨機變化。分析時,g12(r)值落在上下包跡線之間,二者之間無相關性;在上包跡線以上為顯著的正相關;在下包跡線以下為顯著的負相關[44]。
2.5 種群更新分析
為了探討植株的競爭強度及其空間分布格局與種群之間的關系,將樣地內胸徑0—9 cm的幼樹作為更新苗,選擇樣地內胸徑18 cm以上對象木作為母樹,以單株母樹為中心,統計2、4、6、8 m樣圓半徑范圍內4個方向(E、S、W、N)的更新苗的存活數及死亡數。A樣地選取了37株對象木,B樣地選取了44株對象木。
2.6 統計分析
點格局分析通過Programita 2014軟件完成,為降低邊緣效應對分析的影響,在本研究中空間尺度選擇樣地邊長的一半(0—25 m),步長為1 m,采用Monte-Carlo檢驗擬合199次,產生置信水平為99%的上下包跡線以檢驗分析結果的顯著性。其他統計分析都在Excel 2013和SPSS 19.0軟件完成。
3 結果與分析
3.1 對象木徑階與樹高結構
A樣地共調查了368株對象木,最小胸徑0.8 cm,最大胸徑38.7 cm,平均胸徑為9.1 cm;B樣地共調查了285株對象木,最小胸徑為1.2 cm,最大胸徑36.8 cm,平均胸徑11.0 cm。如圖2所示,中小徑階沙地樟子松林木所占比例較高,A樣地徑階I—VII(0—21 cm)的株數為355株,占總株數96.47%,B樣地的徑階I—VII為267株,占總株數94.35%;A樣地、B樣地徑階均呈正態分布(Kolmogorov-Smirnov 正態性檢驗,P值分別為0.696和0.619)(圖2),兩個樣地的樟子松種群均屬于增長型種群結構。且兩個個樣地的高階分布型非常相似(圖2),其平均樹高8.0 m,其中高階VI 10—12 m的林木所占比例最大。

圖2 對象木胸徑與樹高分布特征Fig.2 The distribution features of object trees′ height and DBHI:DBH<3 cm;II:3 cm≤DBH<6 cm;III:6 cm≤DBH<9 cm;IV:9 cm≤DBH<12 cm;V:12 cm≤DBH<15 cm;VI:15 cm≤DBH<18 cm;VII:18 cm≤DBH<21 cm;VIII:21 cm≤DBH<24 cm;IX:24 cm≤DBH<27 cm;X:27 cm≤DBH<30 cm;XI:30 cm≤DBH<33 cm;XII:33 cm≤DBH<36 cm;XIII:36 cm≤DBH<39 cm
3.2 競爭強度
3.2.1競爭范圍的選定
如圖3所示,隨著競爭林木之間距離的增加,林木的平均競爭指數逐漸減小。對競爭指數變化量與林木距離關系研究發現,隨著林木之間距離的增加,競爭指數的變化量逐漸減小,在8 m半徑處有一個明顯的拐點,在8 m半徑以內,競爭指數變化量減小非常明顯(如樣地A 2 m處平均為1.234,4 m處平均為0.644,6 m處平均為0.435;樣地B 2 m處平均為1.197,4 m處平均為0.698,6 m處平均為0.471),而在8—14 m半徑范圍內,半徑每增加2 m,競爭指數變化量約減少0.01的幅度,而14 m之外競爭指數變化量基本保持穩定,競爭木對對象木的影響非常微弱。這說明,競爭指數的變化明顯與林木之間的距離有關,即競爭木距離對象木越近,對象木受到的影響越大。基于上述分析,本研究認為,對于天然沙地樟子松地理分布最南緣區的中幼林來說,8 m是研究確定其林木競爭的最適宜競爭半徑范圍,其競爭強度能夠較好地反映沙地樟子松種內競爭的真實情況。

圖3 樣圓半徑與競爭指數變化的關系Fig.3 Relationship between radius of the round quadrats and the variation of competition intensity
3.2.2樟子松種內競爭強度
隨林木徑階的增大,其競爭指數逐漸減小。由表2可見,A樣地中,I徑階的競爭指數最大,為120.31,顯著高于其他徑階(P<0.05),IX徑階的競爭指數最低,為5.20,顯著低于其他徑階(P<0.05),而IV、V、VI、VII徑階之間無顯著差異(P>0.05)。B樣地中I徑階的競爭指數最大,為113.65,顯著高于其他徑階(P<0.05),IX徑階的競爭指數最低,為5.20,顯著低于其他徑階(P<0.05),而IV、V徑階之間以及VII、VIII、IX徑階之間的無顯著差異(P>0.05)。以上結果表明徑階小的樟子松受到種內競爭強度較大,徑階較大的受到的種內競爭強度較小。

表2 沙地樟子松林分種內競爭強度
不同字母表示不同處理間在5%水平差異顯著(P<0.05);徑階IX、X、XI、XII、XIII的株數太少,缺乏統計意義,因此該處未統計分析
3.2.3競爭強度與胸徑的關系
A樣地的關系為:CI=235.16D-1.10(R2=0.91,P<0.001)(圖4);B樣地為CI=254.41D-1.15(R2=0.90,P<0.001)(圖4);所有林木中對象木胸徑與競爭指數的關系為CI=242.24D-1.12(R2=0.91,P<0.001),兩個樣地均達到極顯著水平,表明競爭強度與對象木胸徑的關系適合用該模型模擬和預測。由圖4可知,隨著對象木胸徑的增大,個體數逐漸減少,且對象木胸徑越大,其所受到的競爭壓力越小,而對其周邊的個體卻可造成極大的競爭壓力。當對象木胸徑達到20 cm 以上,競爭強度趨于穩定,維持在較低水平。

圖4 對象木競爭指數與其胸徑的散點圖Fig.4 Scatter of competition index of object trees and its DBH空心圓代表樟子松,虛線代表冪函數擬合曲線
3.3 樟子松空間格局分析
3.3.1不同生長階段林木的空間格局
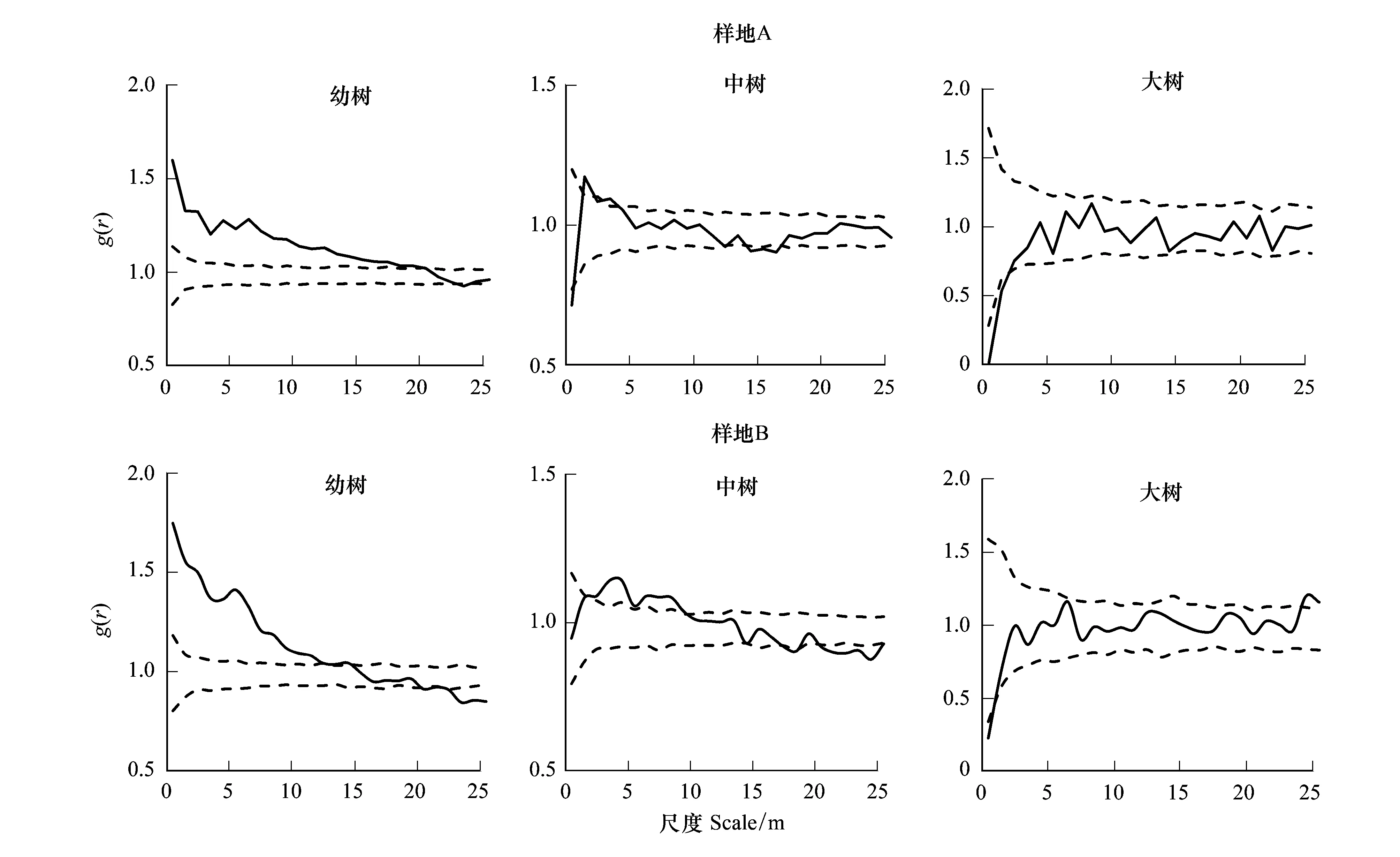
圖5 不同生長階段樹木的空間格局Fig.5 Spatial patterns of Pinus sylvestris var. mongolica among different life stages in sampling sites
如圖5所示,對兩個樣地的樟子松種群進行g(r)單變量分析。A樣地中,幼樹在0—20 m尺度上為顯著聚集分布,隨著尺度的增加,基本上為隨機分布;中樹在0—1 m和13.5—16.5 m尺度上為均勻分布,在1—4 m尺度上為聚集分布,在4—14 m和16.5—25 m尺度上為隨機分布;大樹在0—1.5 m尺度上為顯著均勻分布,其他尺度上為隨機分布。
B樣地中,幼樹在0—14.5 m尺度上為顯著聚集分布,隨著尺度的增加轉變為隨機分布和均勻分布;中樹在2.5—9.5 m尺度上為顯著聚集分布,在20—25 m尺度上為均勻分布,其他尺度上為隨機分布;大樹在0—1.5 m尺度上為顯著均勻分布,其他尺度上為隨機分布。
3.3.2不同生長階段林木的空間關聯分析
如圖6,對樟子松種群的3個生長階段的空間關聯性進行雙變量g12(r)分析,結果表明,A樣地中幼樹與中樹在2.5—7.5 m尺度上呈顯著正相關,B樣地在0—3.5 m尺度上呈顯著正相關,其他尺度上兩個樣地均無相關性;兩個樣地的幼樹和大樹呈相互獨立關系,無空間相關性;A樣地的中樹和大樹在0—1.5 m尺度上呈顯著負相關,而B樣地在0—2.5 m尺度上呈顯著負相關,其他尺度上無空間相關性。

圖6 不同生長階段樹木之間的空間關聯性Fig.6 Spatial association of Pinus sylvestris var. mongolica among different life stages in sampling sites
3.4 幼樹更新
A樣地內更新幼樹占樣地林木總數量的50.51%,B樣地的占43.07%;分別統計對象木的東、南、西、北方向上0—2、2—4、4—6 m和6—8 m樣圓半徑范圍內的更新苗數量,結果如圖7所示,在對象木外圍0—2 m半徑范圍內,更新幼樹的密度最大,在此范圍內A樣地的平均更新幼樹密度為0.09株/m2, B樣地的平均更新幼樹密度為0.08株/m2;而在A和B樣地分別在6—8 m和4—6 m范圍內的更新幼樹密度最低,A平均更新幼樹密度分別為0.03株/m2和0.01株/m2。對不同的方向更新幼樹統計發現,對象木冠層南側的密度最大,平均達到0.43株/m2,冠層西側的密度次之,平均為0.42株/m2,冠層北側更新幼樹密度最低,為0.39株/m2。然而在A樣地6—8 m范圍內,E方向與S和W方向差異顯著(P<0.05),其他范圍內4個方向都無顯著從差異(圖7)。
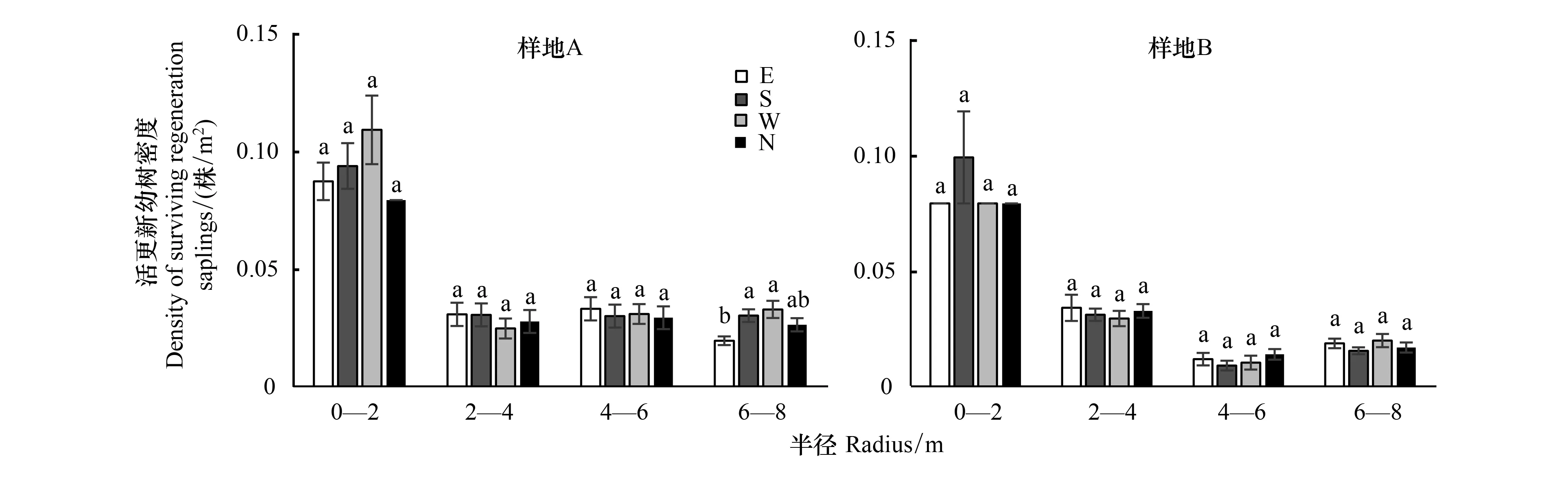
圖7 對象木不同方位和半徑的更新幼樹密度Fig.7 The saplings′ density of different radii in 4 orientations of object trees不同字母表示不同處理間在5%水平差異顯著(P<0.05);E:正東方向,East direction;S:正南方向,South direction;W:正西方向,West direction;N:正北方向,North direction
對A和B樣地內更新幼樹密度與競爭指數進行相關分析,結果如表3,競爭指數與所有更新幼樹密度呈極顯著正相關(P<0.01),與存活更新幼樹密度基本呈正相關。表明隨著競爭指數增加,不同樣圓半徑范圍內的更新幼樹的密度也隨之增加。

表3 更新幼樹密度與競爭指數的相關性
**P<0.01,*P<0.05
4 討論
4.1 種內競爭
在林分內部,林木之間的競爭是經常存在的,由于林分內林木個體的空間生態位大多處于重疊狀況,其個體之間必然為其生長而競爭利用有限的資源如光照、土壤水分和養分等,但林木個體之間的競爭是有一定范圍的,主要發生在相鄰或相近的個體之間[34]。為了確定林木競爭的范圍,目前的方法主要有固定半徑法[45]、樹冠重疊法[46]、角規計數抽樣方法[34]和圓錐調查[47]等方法,但應用較多的是簡便易算的固定樣圓半徑法。基于此方法,張學龍等[48]發現,祁連山青海云杉種群的種內競爭主要發生在6 m范圍內,黃山松的競爭范圍主要集中在7.5 m半徑以內[49],而在渾善達克沙地天然沙地榆種群,其競爭主要發生在8 m半徑范圍以內[50]。
對于天然沙地樟子松地理分布南緣的純林來說,由于林分密度較大,個體之間的競爭是非常明顯的,本研究通過固定樣圓法研究發現,當樣圓半徑介于2—8 m時,其競爭指數持續下降,當超過8 m時,其競爭指數保持在一個較穩定的幅度內,因此,本研究認為8 m是沙地樟子松競爭估算最適宜的范圍,這與毛磊等[51]的研究有所差異,他們認為10 m是最適宜的競爭估算樣圓半徑。這可能與兩個研究地點樟子松樹木大小不同有關。在紅花爾基自然保護區,其天然樟子松較大,林木冠幅半徑多介于3—5 m,胸徑介于18.5—22.9 cm[51],而本研究中的樟子松樹冠半徑多介于2—2.5 m,平均胸徑9.7—10.5 cm。
林木的競爭受到多種因素的影響,其中包括內在因素(如個體大小、所處生長發育階段、個體生活力)和外界因素(如所處地形、氣候、相鄰植株個體大小等)[52]。Hegyi單木競爭指數是測度林木個體間競爭強度較常用的指標[19,32,53-54],它能間接反映出林木個體對光照、水分和營養等資源的現實分配。樹木的大小對其競爭強弱有著較大的影響,本研究擬合了胸徑與其競爭指數的關系,發現競爭強度與對象木胸徑服從冪函數關系, 這與其他一些研究的結果基本一致[19,55]。此外,本研究也發現,當樹木胸徑低于20 cm時,競爭指數變化顯著,而當胸徑大于20 cm時,其競爭強度變化很小,基本趨于穩定。
4.2 空間格局
許多研究已經發現自然生態系統內的幼年個體大多呈聚集分布的特征,然而,隨著個體逐年長大,生態系統內的個體可能會逐漸轉向隨機或均勻分布的狀態[7]。在本研究的中幼林中,幼樹階段的樟子松種群大約在小于15—20 m的尺度上呈聚集分布狀態,在此階段,由于個體較小,其對資源與空間的需求較小,通過聚集分布有利于共同抵抗不良環境的影響,以抵抗外界干擾來維持種群自身穩定性,從而促進種群的繁衍[56-57];而隨著樹木胸徑的增加,個體對資源和空間的需求越來越大,在有限的資源環境約束下,生活力弱的個體由于激烈的競爭而無法獲取水分與營養等資源,種群開始出現自疏現象,在這一作用下,個體之間的相互作用變弱,種群的分布趨向于均勻或隨機分布[58]。而在1—4 m尺度上中樹呈聚集分布特征,這可能是由于生境異質性效應(土壤養分、洼地等)導致的[59]。
許多生態過程會導致物種的聚集分布特征,這是空間格局尺度依賴性的重要理論基礎[60]。許多研究表明,種子傳播限制和生境異質性(如土壤養分與水分、地形差異等)是影響物種聚集分布的重要因子[61-63]。本研究中,種子繁殖是樟子松種群的主要繁殖方式[64-65],種子成熟后會在重力和風等因素的共同作用下落到地面,以母樹為中心形成聚集分布狀態,而隨著與母樹距離的增加樟子松的種子密度逐漸降低[64-65],或者在風力、地面徑流、地面植物的阻截等外力作用下形成聚集狀態。因此,種子傳播限制是影響樟子松幼樹聚集分布格局的重要因子。此外,生境異質性也是樟子松幼樹聚集分布的重要影響因素之一,在沙丘的作用下,土壤、地形、光照等資源呈現出空間上的斑塊狀分布,資源良好的生境為種子的萌發和定植創造了有力條件,從而導致物種的聚集分布[66]。
在中或較大尺度上,幼樹、中樹和大樹之間彼此都表現相互獨立的關系,但幼樹和中樹在較小尺度上呈正相關,這可能是由于中樹為其種子的萌發和幼樹的生長提供適宜的微環境,形成了一種相互有利的空間關系[67-68],即植物個體間的“庇護與被庇護”效應[69]。由于具備更多的可用水量、更高的土壤養分以及良好的遮陰環境,“庇護個體”以其固定的樹冠可以持續地為種子的萌發、幼苗的存活提供良好的環境[70]。幼樹與大樹在所有尺度上無相關性,這或許是由于大樹對幼樹造成極大的競爭壓力,使得幼樹無法獲取生存所需資源,最后導致大樹周圍的幼樹存活率降低[71],從而主要分布于競爭壓力適宜的中樹周圍;中樹和大樹在小尺度上呈負相關,這可能與兩者的營養生態位發生重疊有關,中樹在獲取資源及生存空間時會受到大樹的抑制,中樹的平均冠幅達1.8 m,而大樹的平均冠幅約3.0 m,且兩者的冠層在小尺度上發生較大的重疊效應,這支持了本研究的結果,而在較大尺度上,兩者之間的競爭效應減弱,從而表現出相互獨立的關系。
4.3 種群更新
種群更新是一個極為重要的生態學過程,對群落演替與動態變化具有重要的影響,更新過程涉及到種子擴散、土壤種子庫、種子萌發與生長、幼樹競爭等方面[72-74]。本研究發現樟子松幼樹更新多分布于樹冠的南側和東側,這與閆德仁等[73-74]的研究結果較為一致,他們在進行土壤種子庫研究時發現,樹冠南側土壤種子庫的種子數量最多,且隨距樹干距離(2—6 m)增加而增加,但距樹干8 m后降低。另外,本研究也發現更新幼樹密度最大的區域多位于樹木外圍的0—2 m半徑范圍內,即樹冠下,這與閆德仁等[73-74]的研究稍有不同,他們認為樟子松更新苗主要集中在母樹樹冠的邊緣和距離母樹10 m范圍內,這可能與閆德仁等[73-74]研究的樹木冠幅較大有關。
種群更新受許多生物因素如母樹大小、密度[73]和環境因素如氣候[73-74]、火干擾[19,24- 25]等的影響。本研究分析了母樹競爭與幼樹更新的關系,發現隨著競爭指數增加,不同樣圓半徑范圍內的更新幼樹的密度也隨之增加。即競爭指數越高,胸徑越小,個體的競爭能力就越弱,其受周圍個體的競爭壓力就越強,而對周圍更新幼樹產生的競爭壓力就會越小,因此更新幼樹趨向于分布在競爭指數高的個體周圍。
本研究發現更新幼樹呈聚集分布的空間格局,在小尺度上,更新幼樹與中樹呈正相關,而在較大尺度上,與中、大樹之間均呈相互獨立的關系,這與尤海舟等[71]的結果相似。即兩個徑階之間差異越小,其個體之間的正相關就越強,個體之間更趨向于協調[71]。而母樹競爭強弱又會影響著幼樹的更新與格局,即隨著競爭指數的增加,更新幼樹的密度會逐漸增加。
在本研究區的沙地樟子松純林中,林分受外界干擾較少,天然更新良好,種群處于增長狀態,但由于林分密度較大,個體受競爭的影響非常大,推測未來競爭可能導致生活力弱的個體死亡率增加,自疏作用增強。在競爭壓力下,林分的空間格局可能發生改變,導致形成許多林隙,從而使樟子松的幼樹多以集群狀態分布在林隙邊緣競爭較弱的個體周圍,從而加速本區沙地樟子松種群的擴張。
5 結論
本研究采用Hegyi競爭指數與點格局等分析方法,分析了呼倫貝爾沙地東南端沙地樟子松天然林沙地樟子松的徑階和樹高結構、種內競爭、不同生長階段空間格局及空間關聯性、以及幼樹更新特征,主要結論如下:1) 在地理分布南部邊緣區的沙地樟子松林處于中幼齡階段,屬增長型種群;2) 沙地樟子松對象木胸徑與競爭指數服從冪函數關系,隨著林木胸徑的增加,其競爭指數逐漸減小(即競爭強度越強);3) 林木幼樹在小尺度上呈聚集分布特征,而隨著胸徑的增加,在自疏等作用影響下,個體之間相互作用變弱,促使中樹與大樹在中大尺度呈隨機分布;幼樹與中樹在“庇護與被庇護”效應的作用下導致其在小尺度呈正相關,而中樹與大樹由于營養生態位的重疊,導致兩者在小尺度呈負相關;4) 競爭指數與更新幼樹和存活更新幼樹的密度均呈顯著正相關(P<0.05),即更新幼樹趨向于分布在競爭較弱的林木個體周圍。未來在沙地樟子松的經營管理中,需要充分考慮林木競爭、空間格局以及種群更新的關系,統籌規劃,采取合理的培育措施,以促進沙地樟子松種群的天然更新。
