α1-腎上腺素受體結(jié)構(gòu)改變影響其生物學(xué)活性及生理功能的研究進(jìn)展
2019-06-24 00:59:14楊澤平邵鈺婷郭曉汐徐天瑞
中國藥理學(xué)通報 2019年6期
馬 力,楊澤平,母 茜,邵鈺婷,楊 洋,安 輸,郭曉汐,郝 倩,徐天瑞,劉 瑩
(昆明理工大學(xué)生命科學(xué)與技術(shù)學(xué)院細(xì)胞信號傳導(dǎo)實驗室,云南 昆明 650500)
1994年8月,國際藥理聯(lián)合會受體命名與藥物分會根據(jù)腎上腺素受體(adrenergic receptor,AR)與其配體結(jié)合特性及功能等差異,將AR分為α亞型(包括α1和α2)、β亞型(包括β1、β2、β3)兩大類。α1-ARs在許多生理過程中發(fā)揮重要作用,包括平滑肌收縮、心肌變時變力、肝糖原代謝等等。大量藥理學(xué)研究表明,結(jié)構(gòu)同源但具不同特性的α1-ARs亞型,調(diào)控上述生理過程。不同組織細(xì)胞上的α1-ARs的藥理學(xué)特性并不均一,存在多種亞型,按照國際藥理聯(lián)合會受體命名與藥物分類委員會規(guī)定,將其分為α1A、α1B和α1D3種亞型。根據(jù)該受體與激動劑和拮抗劑親和力的不同,以及對烷化劑氯乙烷、可樂定敏感度的差異,首先鑒定出α1A-AR和α1B-AR。此后,應(yīng)用cDNA分子克隆技術(shù),克隆出人類3種α1-ARs(A、B、D)亞型。
近年來,應(yīng)用分子生物學(xué)技術(shù)、放射配體結(jié)合技術(shù)、拮抗劑功能分析等手段,對α1-ARs亞型的蛋白結(jié)構(gòu)、分類、分布等各方面的研究均取得很大進(jìn)展,但是,對α1-ARs結(jié)構(gòu)改變影響其生理功能的研究較少。本文總結(jié)了α1-ARs受體羧基端截短、二聚化、別構(gòu)效應(yīng)和點突變這4個方面的結(jié)構(gòu)改變,導(dǎo)致其內(nèi)吞、磷酸化、脫敏以及對激動劑的親和力等生理功能的改變,以期為α1-ARs受體的藥理、生理及病理生理學(xué)功能研究提供理論基礎(chǔ)。
1 α1-AR的結(jié)構(gòu)及分布
1.1 α1-AR的結(jié)構(gòu)α1-ARs屬于G蛋白偶聯(lián)受體(G protein-coupled receptors,GPCRs),為單一的多肽鏈,包含7個跨膜螺旋。α1-ARs的3種亞型的氨基末端均位于細(xì)胞外,但長度不盡相同,羧基端均位于細(xì)胞內(nèi),氨基酸序列的同源性較小,但該區(qū)域大約在第7個跨膜螺旋后的第16個氨基酸有1個保守的半胱氨酸殘基,可能與翻譯后的硫酯化過程有關(guān)。α1-ARs跨膜區(qū)域由3個細(xì)胞內(nèi)和3個細(xì)胞外回環(huán)結(jié)構(gòu)組成。3種受體亞型中,構(gòu)成這些回環(huán)的氨基酸盡管有所不同,但長度基本相似。第1個和第2個細(xì)胞外回環(huán)結(jié)構(gòu)均含有1個半胱氨酸殘基。半胱氨酸殘基在所有GPCRs中均是高度保守的,由于它涉及二硫鍵的形成,因此,對于蛋白質(zhì)的折疊、磷酸化及在質(zhì)膜中的表達(dá)均發(fā)揮關(guān)鍵作用。
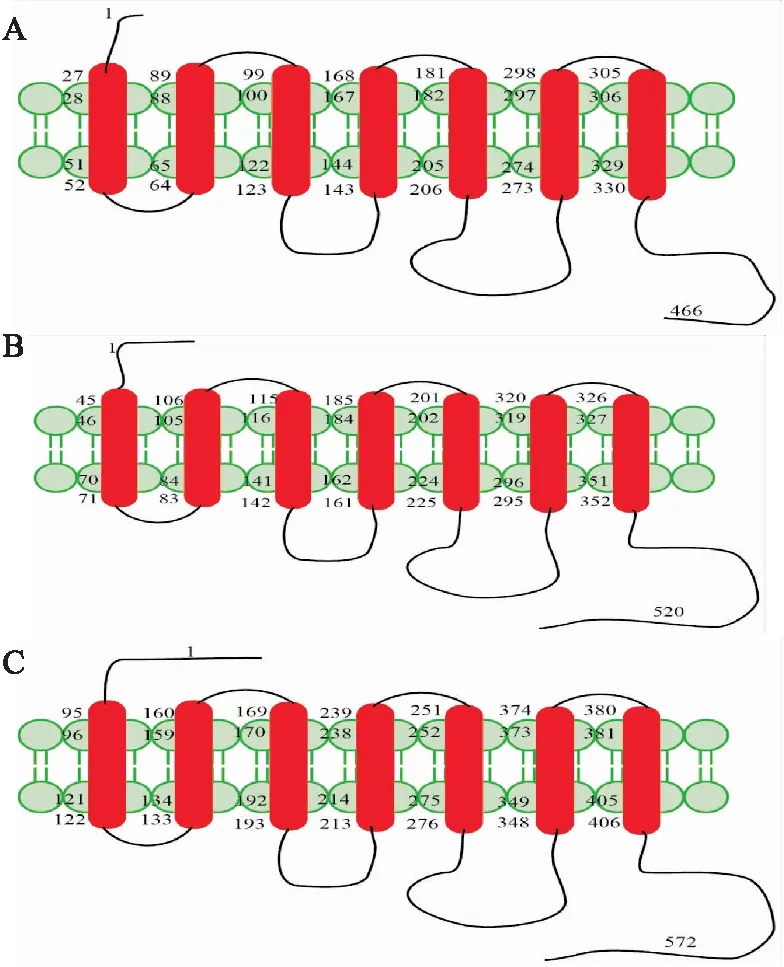
Fig 1 Structure diagram of α1-ARs
The α1-ARs consist of 7 transmembrane helices, 3 intracellular loops and 3 extracellular loops, as well as an extracellular amino terminus and an intracellular carboxy terminus. A: The amino terminus of α1A-AR consists of 27 amino acids and the carboxy terminus consists of 137 amino acids; B: The amino terminus of α1B-AR consists of 45 amino acids and the carboxy terminus consists of 169 amino acids; C: The amino terminus of α1D-AR consists of 95 amino acids and the carboxy terminus consists of 167 amino acids.
1.2 α1-AR的分布采用Western blot方法研究腦、心臟、腎臟、肝臟、輸精管、前列腺、腎上腺等組織中,α1-ARs各亞型蛋白的表達(dá)情況,發(fā)現(xiàn)α1A受體亞型絕大部分分布在腦、心臟、血管、肝、輸精管、腎上腺,少部分分布在腎臟、前列腺;α1B受體亞型在腦、心臟分布較高;而α1D受體亞型在腦分布較高。大腦皮層中的3種亞型受體隨著年齡的增長而增加,其中α1B亞型在心臟中的分布隨年齡的增加而增加,在肝臟中的分布隨年齡的增加而減少。在肝臟、腎臟、心臟的α1D和α1A亞型不受年齡的影響。
研究表明,α1-AR在不同亞細(xì)胞結(jié)構(gòu)上的表達(dá)存在差異。現(xiàn)已知α1A-AR主要表達(dá)在細(xì)胞內(nèi),但在成纖維細(xì)胞和血管平滑肌細(xì)胞表面也有一定程度的表達(dá);α1B-AR主要表達(dá)在細(xì)胞膜表面;而α1D-AR主要表達(dá)在細(xì)胞內(nèi)和細(xì)胞核周圍。
2 α1-AR結(jié)構(gòu)改變導(dǎo)致其生理功能的變化
2.1 羧基端結(jié)構(gòu)改變在結(jié)構(gòu)上,GPCRs是1條單一的多肽鏈,包含一段胞外氨基末端(N端)、7個由22-28個疏水性氨基酸組成的α螺旋形成的跨膜結(jié)構(gòu)域,以及1個胞內(nèi)的羧基末端(C端)[1]。連接疏水性氨基酸跨膜片段的親水性氨基酸片段組成3個細(xì)胞外環(huán)與3個細(xì)胞內(nèi)環(huán)。在GPCRs結(jié)構(gòu)中,胞外N端或跨膜區(qū)形成配體結(jié)合域,位于胞內(nèi)側(cè)的C末端包含絲氨酸和蘇氨酸的磷酸化位點,而連接跨膜區(qū)段的胞內(nèi)環(huán)和C末端形成G蛋白結(jié)合域[2]。配基與受體結(jié)合后,將信號傳遞給與受體偶聯(lián)的G蛋白,導(dǎo)致下游信號的產(chǎn)生,受體與配基結(jié)合后發(fā)生的構(gòu)象變化決定了下游信號的特異性[3]。而C-端在G蛋白活化幅度方面起關(guān)鍵作用,同時C-端的近端也參與受體與G蛋白的偶聯(lián)。C-端截短的α1B-AR突變體則表現(xiàn)出激動劑依賴的磷酸化和受體內(nèi)陷作用的消失[4-5](Tab 1)。
先前已經(jīng)證明,C-端是α1-ARs的重要調(diào)節(jié)域。它的完整性是α1-ARs脫敏和內(nèi)吞作用所必需的[6]。因此,為了研究C-端對α1A-AR、α1B-AR脫敏和內(nèi)吞的影響,Uberti等[7]通過用一種受體亞型的C末端替換另一種受體亞型的C末端,來設(shè)計α1A/α1B嵌合體,并且監(jiān)測它們的內(nèi)吞作用,證明α1B-AR的C端完整性是α1B-AR內(nèi)吞作用所必需的(Tab 1)。
2.2 二聚化α1-ARs的3種亞型均可形成同源二聚體,但是異源二聚體只發(fā)生在α1B-AR與α1A/α1D-AR之間,α1A-AR與α1D-AR的二聚體并未發(fā)現(xiàn),表明異二聚體的形成僅發(fā)生在亞型的特定組合之間。Uberti等[7]通過α1-ARs與標(biāo)記有Flag-或HA-不同表位標(biāo)簽的受體免疫共沉淀,為HEK293細(xì)胞中所有3種亞型的同源二聚化提供了直接證據(jù)(Tab 2)。Uberti等[7]研究還表明,α1-AR的同源和異源二聚體復(fù)合物均在細(xì)胞膜表面表達(dá),但是在這些復(fù)合物中難以確定哪些受體實際存在以及具體分布比例。
Gao等[8]發(fā)現(xiàn),α1A/α1B-AR能與趨化因子(C-X-C基序)受體4[chemokine (C-X-C motif) receptor 4,CXCR4]形成異源二聚體,且該異二聚體復(fù)合物在人血管平滑肌細(xì)胞(human vascular smooth muscle cell,hVSMC)的細(xì)胞膜上組成型表達(dá),二聚體的形成能增強α1-ARs對激動劑去氧腎上腺素(phenylephrine,PE)的敏感性,這表明二聚體可以為調(diào)節(jié)血管功能和血壓的替代療法提供作用靶點(Tab 2)。Albee等[9]發(fā)現(xiàn),α1A/B/D-AR能與非典型趨化因子受體3(atypical chemokine receptor 3,ACKR3)形成異二聚體,該復(fù)合物在hVSMC的細(xì)胞膜表面上組成型表達(dá),二聚體的形成能夠負(fù)調(diào)節(jié)α1-ARs。靶向α1-ARs和ACKR3異源二聚體的藥物研發(fā),能夠為血管類疾病的治療提供新的靶點和方向(Tab 2)。
受體的二聚化對于受體活化及信號轉(zhuǎn)導(dǎo)是必需的。α1B-AR與復(fù)雜的G蛋白、信號蛋白或支架蛋白相偶聯(lián),并能與AR形成同源二聚體或異二聚體[10-12]。受體的二聚化對細(xì)胞的特性和功能的影響如下:(1)二聚體的形成對受體的細(xì)胞表面表達(dá)和功能發(fā)揮是必需的。α1D-AR和α1B-AR形成異源二聚體是α1D-AR在細(xì)胞膜表面正常表達(dá)所必需的。α1B-AR可能通過掩蓋α1D-AR的N端內(nèi)質(zhì)網(wǎng)滯留結(jié)構(gòu)域,促進(jìn)α1D-AR在細(xì)胞膜上的表達(dá)[7]。(2)二聚體對配體的親和力表現(xiàn)出一定的偏向性,可增強或降低對配體的結(jié)合力。α1B-AR和α1D-AR二聚體能明顯增加α1B-AR與腎上腺素配體的結(jié)合能力[4]。表明GPCRs二聚化與配體結(jié)合有正協(xié)同作用。
2.3 別構(gòu)效應(yīng)別構(gòu)效應(yīng)又稱為變構(gòu)效應(yīng),是寡聚蛋白與配基結(jié)合改變蛋白質(zhì)的構(gòu)象,進(jìn)而導(dǎo)致蛋白質(zhì)生物活性改變的現(xiàn)象。由于某些不直接影響蛋白質(zhì)活性的物質(zhì),結(jié)合于蛋白質(zhì)活性部位以外的其他部位(即別構(gòu)部位),引起蛋白質(zhì)分子的構(gòu)象變化,進(jìn)而導(dǎo)致蛋白質(zhì)活性改變的現(xiàn)象。變構(gòu)調(diào)節(jié)劑具有克服別構(gòu)效應(yīng)的潛力,但是對于α1-ARs家族的變構(gòu)調(diào)節(jié)劑的研究很少,關(guān)于任何變構(gòu)位點的結(jié)構(gòu)信息甚至更少。Campbell等[13]的研究表明,變構(gòu)相互作用與結(jié)合位點相關(guān)。即最佳大小的二價配體將促進(jìn)變構(gòu)位點的結(jié)合,從而增強配體的變構(gòu)效應(yīng),否則無效力或效力差。
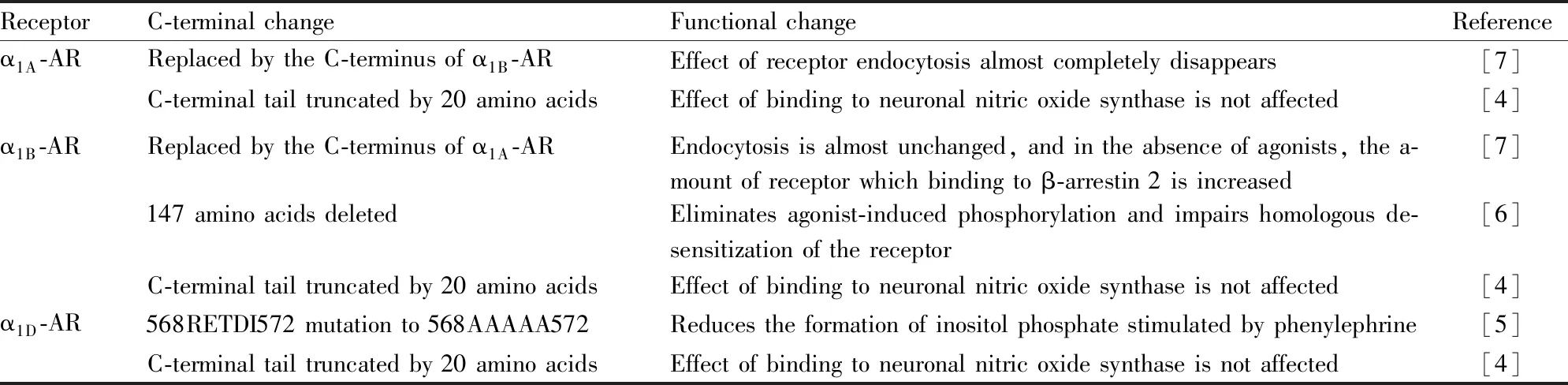
Tab 1 Changes in function induced by C-terminus of α1-AR
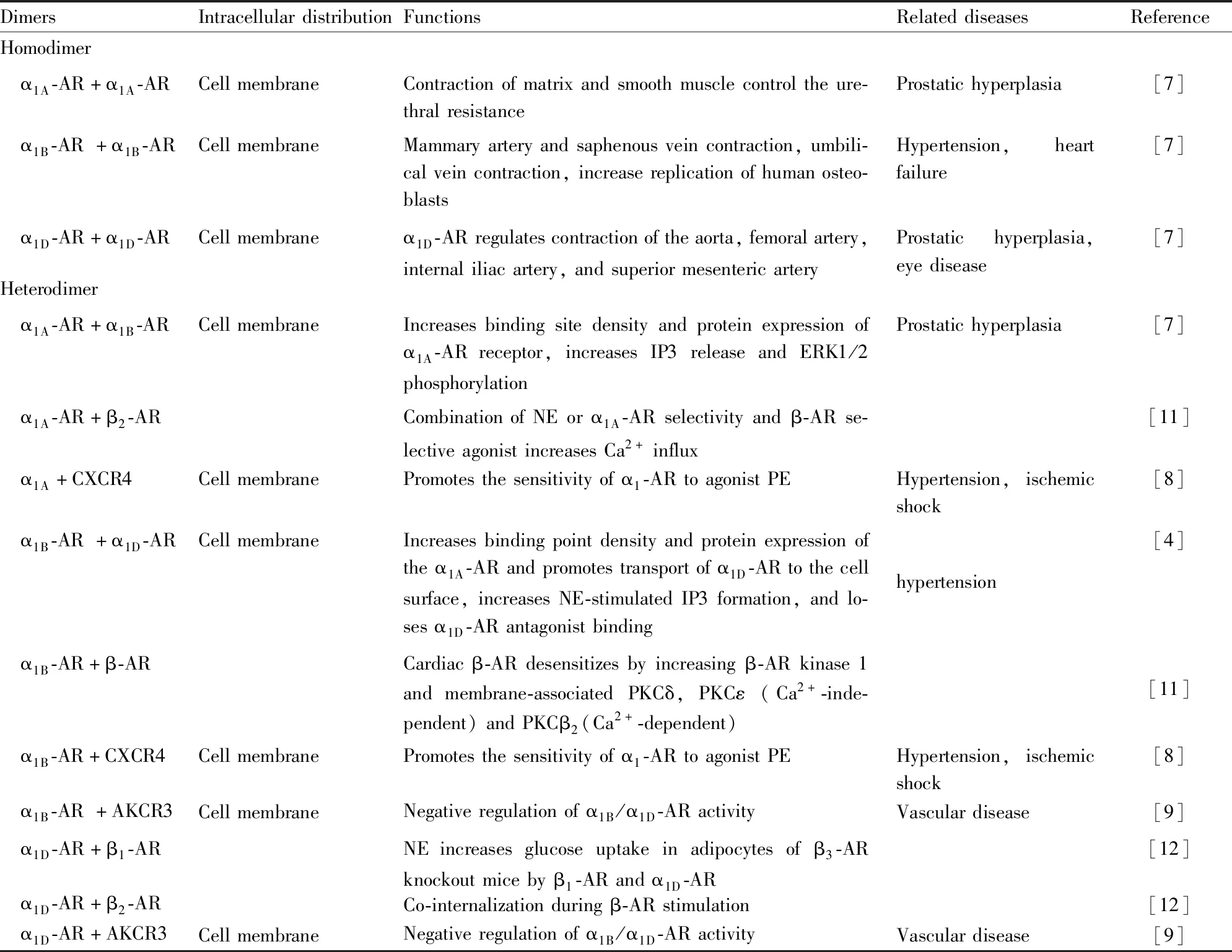
Tab 2 Effect of dimerization on physiological function of α1-AR
Ciolek等[14]基于細(xì)胞的功能實驗證明,Zn2+和Cu2+可以通過降低激動劑功效,拮抗AR活化,此時AR活化曲線幾乎無變化。該研究證明了金屬離子與α1-ARs之間的相互作用,說明Zn2+和Cu2+可能具有調(diào)控前列腺功能的生物學(xué)作用。Zn2+和Cu2+的變構(gòu)作用主要歸因于哌唑嗪(α1-AR阻滯劑)動力學(xué)結(jié)合率的降低。盡管AR中的變構(gòu)位點代表了新型亞型選擇性藥物的替代靶點,但它們的研究很少。繼阿米洛利后,Zn2+和Cu2+是第2類分子,已發(fā)現(xiàn)它們作為哌唑嗪與α1-ARs結(jié)合的負(fù)變構(gòu)調(diào)節(jié)劑而發(fā)揮作用。
2.4 點突變α1-ARs結(jié)構(gòu)上關(guān)鍵位點的氨基酸對其生理功能至關(guān)重要,關(guān)鍵位點的氨基酸突變體在受體磷酸化、脫敏、受體異構(gòu)化及受體與激動劑結(jié)合的親和力等生理功能方面均會發(fā)生改變。
研究表明,Ser394和Ser400在佛波醇酯誘導(dǎo)蛋白激酶C(protein kinase C,PKC)活化后被磷酸化,而Ser404、Ser408和Ser410可以被α1B-AR激動劑腎上腺素激活。G蛋白偶聯(lián)受體激酶2(G protein-coupled receptor kinase 2,GRK2)介導(dǎo)的Ser404、Ser408和Ser410的磷酸化可能引發(fā)α1B-AR的脫敏。以上結(jié)果表明,缺乏這3種絲氨酸的受體,α1-AR通過GRK2介導(dǎo)受體脫敏的能力受損[15]。在α1B-AR的第3個細(xì)胞內(nèi)環(huán)的保守取代(A293E),導(dǎo)致受體組成型激活。無激動劑刺激的情況下,與表達(dá)野生型α1B-AR的細(xì)胞相比,表達(dá)突變受體的細(xì)胞表現(xiàn)出更高基礎(chǔ)水平的肌醇磷酸。為了進(jìn)一步評價A293E的作用,將A293分別突變成其他19種氨基酸[16]。結(jié)果表明,α1B-AR中A293的所有可能的氨基酸點突變體誘導(dǎo)了不同水平的受體組成型激活(Tab 3)。此外,α1B-AR的D142具有疏水、親水雙特性,可以通過該殘基的質(zhì)子化、去質(zhì)子化來調(diào)節(jié),是α1B-AR的無活性(R)和活性(R*)狀態(tài)之間轉(zhuǎn)變的重要調(diào)節(jié)劑(Tab 3)。此外,結(jié)合先前描述的α1B-AR突變體A293E對F310突變體進(jìn)行研究,結(jié)果表明,F(xiàn)310與激動劑兒茶酚胺的相互作用,對于從基礎(chǔ)狀態(tài)R到完全活化狀態(tài)R*的異構(gòu)化是必不可少的[17](Tab 3)。
研究發(fā)現(xiàn),R143的主要作用是介導(dǎo)受體激活,允許第2個細(xì)胞內(nèi)環(huán)(2-loop)中的幾個氨基酸,以及第3個細(xì)胞內(nèi)環(huán)(3-loop)的N末端和C末端獲得正確的構(gòu)型,以形成具有對接互補性的位點。表明靶向R143的突變能夠完全誘導(dǎo)α1B-AR失活[18]。單突變體A204V對兒茶酚胺腎上腺素和去甲腎上腺素(norepinephrine,NE)顯示出增強的結(jié)合親和力(3~10倍)。該突變體對羥甲唑啉以及其他咪唑啉激動劑的親和力也增加10倍,這是該受體的兩個其他組成型激活突變所不具備的作用。表明親和力的增強是A204V突變,導(dǎo)致配體結(jié)合口袋的藥理學(xué)特異性改變的結(jié)果。此外,Y203突變成丙氨酸導(dǎo)致受體活化減少,以及對激動劑結(jié)合的親和力適度降低也已被證實[19](Tab 3)。

Tab 3 Changes in physiological functions induced by α1-AR mutations
3 總結(jié)和展望
本文主要總結(jié)了α1-ARs結(jié)構(gòu)改變導(dǎo)致其生理功能的變化,以及該受體在生理和病理過程中的調(diào)節(jié)作用。研究α1-ARs激活參與的生物學(xué)過程,有助于了解人類疾病,尤其是心腦血管疾病的發(fā)生與發(fā)展過程。了解心腦血管疾病的分子基礎(chǔ)后,對后續(xù)心腦血管疾病的早期檢測、診斷、預(yù)防以及新的治療策略提供可能。相信在不久的將來,人們通過對α1受體活化的深入研究,可以更加明確其在心腦血管疾病發(fā)生、發(fā)展中的作用機制,為指導(dǎo)心腦血管疾病基因治療提供新靶點。雖然α1-ARs活化在人體生理和病理過程中的重要作用已被逐漸證實,但具體的機制,如與α1-ARs活化密切相關(guān)的心肌肥厚的發(fā)病機制等尚不明確。因此,闡明α1-ARs的關(guān)鍵調(diào)控因素,對臨床研究心肌肥厚的發(fā)生及開發(fā)逆轉(zhuǎn)心肌肥厚的藥物均具有十分重要的理論意義和實用價值。