重組α-1,3-葡萄糖苷酶的畢赤酵母表達(dá)及發(fā)酵優(yōu)化
2019-06-18 09:04:46王曉燕
食品工程 2019年4期
關(guān)鍵詞:生長(zhǎng)
王曉燕 江 波 張 濤
(江南大學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,江蘇無(wú)錫 214122)
α-葡萄糖苷酶(EC 3.2.1.20),可從麥芽低聚糖、異麥芽糖、α-葡聚糖等底物的非還原末端釋放α-葡萄糖,或通過(guò)轉(zhuǎn)糖基作用將葡萄糖基從底物轉(zhuǎn)移至受體形成相應(yīng)非發(fā)酵性的低聚糖或糖脂、糖肽等。在自然界中分布廣泛,種類繁多,幾乎在所有的生命體中皆有發(fā)現(xiàn)。根據(jù)氨基酸分類系統(tǒng),α-葡萄糖苷酶分屬于5 個(gè)糖苷水解酶家族:GH4,GH13,GH31,GH63 和GH97,大多α-葡萄糖苷酶均屬于GH13 和GH31 家族。α-葡萄糖苷酶的應(yīng)用研究主要集中在制備低聚異麥芽糖等方面,近年來(lái),隨著對(duì)α-1,3 糖苷鍵具有高度選擇性的α-葡萄糖苷酶的發(fā)現(xiàn),有報(bào)道表明可利用α-葡萄糖苷酶催化麥芽低聚糖等制備黑曲霉低聚糖(Nigerooligosaccharides,NOS)。
黑曲霉低聚糖是一類分子中含有α-1,3 糖苷鍵的新型功能性低聚糖,主要包括黑曲霉糖(Nigerose)、黑曲霉糖基葡萄糖(Nigerosylglucose,NOG)和黑曲霉糖基麥芽低聚糖(Nigerosyl matooligosaccharides,NMS)等。黑曲霉糖天然存在于清酒、豆醬等發(fā)酵型產(chǎn)品中,含量較低,難以進(jìn)行大規(guī)模的提取工作,而對(duì)于聚合度更高的黑曲霉低聚糖,在自然界中幾乎不存在,因此黑曲霉低聚糖的生產(chǎn)方法主要有化學(xué)合成、多糖水解法和糖基轉(zhuǎn)移法3 種。據(jù)報(bào)道,黑曲霉低聚糖不僅具有一般功能性低聚糖所共有的熱量低、抗齲齒、促進(jìn)雙歧桿菌增殖等生理功能,還具有免疫激活、改善風(fēng)味、抑制色素褪色等多種作用,可廣泛應(yīng)用于醫(yī)學(xué)、食品等多個(gè)行業(yè)。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株及質(zhì)粒
細(xì)菌克隆宿主E.coli DH5α、真菌表達(dá)宿主P.pastoris GS115 均為本實(shí)驗(yàn)室保藏,含有源自Acremonium sp.S4G13 α-1,3-葡萄糖苷酶基因的重組質(zhì)粒pPIC9K 為委托上海捷瑞生物工程公司合成。
1.1.2 培養(yǎng)基
LB 培養(yǎng)基(g/L):酵母提取物10 g,胰蛋白胨5 g,NaCl 10 g;MD 培養(yǎng)基(g/L):YNB 13.4,生物素4×10-4,葡萄糖20g,瓊脂15 g;YPD 培養(yǎng)基(g/L):酵母提取物10 g,蛋白胨20 g,葡萄糖20 g;BMGY 培養(yǎng)基(g/L):YNB 13.4 mL,生物素4×10-4,甘油30 mL,0.1 mol/L 磷酸鉀緩沖液;BMMY培養(yǎng)基(g/L):YNB 13.4 mL,生物素4×10-4,甲醇0.5%,0.1 mol/L 磷酸鉀緩沖液。
1.1.3 主要試劑
黑曲霉低聚糖、麥芽三糖標(biāo)準(zhǔn)品,購(gòu)自Sigama公司;蛋白胨、酵母粉、YNB、生物素、G418、卡那霉素,均購(gòu)自上海生工生物工程公司;麥芽糖、葡萄糖、磷酸氫二鈉、磷酸二氫鈉、甘油等,均為國(guó)產(chǎn)分析純?cè)噭?gòu)自國(guó)藥集團(tuán)化學(xué)試劑公司;DL 10000、15000 DNA Marker、BglⅡ限制性內(nèi)切酶、4S Green Plus 無(wú)毒核酸染料,購(gòu)自大連寶生物工程有限公司;質(zhì)粒小量制備試劑盒,購(gòu)自上海捷瑞生物工程有限公司。
1.2 主要設(shè)備及儀器
GI54DWS 高壓滅菌鍋,廈門致微儀器有限公司,THZ-100 恒溫培養(yǎng)搖床,上海一恒科技有限公司;Centrifuge 5804R 高速冷凍離心機(jī)、Authorized Thermal Cycler PCR 儀、電轉(zhuǎn)儀、電轉(zhuǎn)杯,德國(guó)Eppendorf 公司;Waters e2695 型高效液相色譜儀,美國(guó)Waters 公司;Asahipak NH2P-50 4E 氨基柱、Shodex 示差折光檢測(cè)器,日本Shodex 公司;冷凍真空干燥機(jī),Labconco 公司。
1.3 P.pastoris GS115 重組菌的構(gòu)建
1.3.1 重組質(zhì)粒pPIC9K-ALG 的構(gòu)建
通過(guò)NCBI 基因文庫(kù),查找到來(lái)源于Acremonium sp.S4G13 的編碼α-葡萄糖苷酶的基因,并將此基因片段委托上海捷瑞生物工程有限公司進(jìn)行合成,在該基因的5' 和3' 末端分別引入EcoR I 和Not I2 個(gè)限制性酶切位點(diǎn),將基因片段連接入酵母表達(dá)載體pPIC9K,合成重組質(zhì)粒pPIC9K-AGL。
1.3.2 重組質(zhì)粒的轉(zhuǎn)化
重組質(zhì)粒pPIC9K-agl 經(jīng)使用BglⅡ快切酶線性化后,加入P.pastoris GS115 感受態(tài)細(xì)胞中以1.5 kV電壓進(jìn)行電轉(zhuǎn)化。電轉(zhuǎn)液在30℃條件下經(jīng)1 h~2 h孵育后涂布于MD 平板,30 ℃倒置培養(yǎng)2 d~3 d直至長(zhǎng)出單菌落。挑選MD 平板上的單菌落依次接種至0.5 mg/mL、1.0 mg/mL、2.0 mg/mL、3.0 mg/mL的YPD-G418 平板上,30℃倒置培養(yǎng)2 d~3 d。同時(shí)轉(zhuǎn)化pPIC9K 空質(zhì)粒作為對(duì)照。
1.3.3 重組菌的培養(yǎng)及誘導(dǎo)產(chǎn)酶
1.3.3.1 種子培養(yǎng)
將MD 平板上的重組酵母單菌落接種到5 mL YPD 試管培養(yǎng)基中,30 ℃、200 r/min 條件下培養(yǎng)16 h。
1.3.3.2 擴(kuò)大培養(yǎng)
以體積濃度2%的接種量將上述菌液接入裝液量為50 mL BMGY 培養(yǎng)基的250 mL 錐形瓶中,30 ℃、200 r/min 條件下培養(yǎng)24 h。
1.3.3.3 誘導(dǎo)培養(yǎng)
離心收集上述菌體,在無(wú)菌條件下將其全部轉(zhuǎn)接入裝液量為50 mL BMMY 培養(yǎng)基的250 mL 錐形瓶中,30 ℃、200 r/min 條件下培養(yǎng)120 h,每24 h補(bǔ)加0.5%甲醇。
1.3.4 酶活測(cè)定方法及定義
將0.9 mL 的100 g/L 麥芽糖溶液與0.1 mL 的粗酶液在50 mmol/L、pH 7.0 磷酸緩沖液體系中充分混勻,55 ℃下反應(yīng)12 h,沸水浴10 min 終止反應(yīng)。反應(yīng)液經(jīng)樹脂除鹽及微孔過(guò)濾后經(jīng)HPLC 檢測(cè),用歸一化法(黑曲霉糖基葡萄糖占總糖的百分比)計(jì)算黑曲霉糖基葡萄糖的轉(zhuǎn)化率,以葡萄糖作為內(nèi)標(biāo)計(jì)算黑曲霉糖基葡萄糖的生成量,計(jì)算酶活。
HPLC 檢測(cè)條件:Waters e2695 型高效液相色譜儀,色譜柱AsahipakNH2P-504E(4.6mm×250mm),示差折光檢測(cè)器,流動(dòng)相為乙腈∶水=75 ∶25(v/v),流速1 mL/min,柱溫30 ℃。
酶活定義:在上述反應(yīng)條件下,每小時(shí)生成1 μmol 的黑曲霉糖基葡萄糖所需酶量為一個(gè)酶活單位(1 U)。
1.4 重組P.pastoris GS115 產(chǎn)酶條件的優(yōu)化
1.4.1 接種量對(duì)重組P.pastoris GS115 生長(zhǎng)階段的影響
重組菌活化后,分別以1%、3%、6%、10%、15%的接種量接入50 mL/250 mL BMGY 培養(yǎng)基中,30 ℃、200 r/min 培養(yǎng),每隔3 h 取樣,測(cè)定OD600監(jiān)測(cè)重組菌的生長(zhǎng)狀況。
1.4.2 碳源對(duì)重組P.pastoris GS115 生長(zhǎng)階段的影響
重組菌活化后,以10%的接種量分別接種至甘油比例為1%、2%、3%、4%、5%的50 mL/250 mL BMGY 培養(yǎng)基中,30 ℃、200 r/min 培養(yǎng),每隔3 h取樣,測(cè)定OD600 監(jiān)測(cè)重組菌的生長(zhǎng)狀況。
1.4.3 初始pH 對(duì)重組P.pastoris GS115 生長(zhǎng)階段的影響
重組菌活化后,以10%的接種量分別接種到pH 值為5.0、6.0、7.0、8.0 的50 mL/250 mL BMGY培養(yǎng)基中,30 ℃、200 r/min 培養(yǎng),每隔3 h 取樣,測(cè)定OD600 監(jiān)測(cè)重組菌的生長(zhǎng)狀況。
1.4.4 初始甲醇體積濃度對(duì)重組P.pastoris GS115 誘導(dǎo)階段的影響
無(wú)菌條件下,將BMGY 培養(yǎng)基中的全部菌體轉(zhuǎn)接入50 mL/250 mL BMMY 培養(yǎng)基中,初始甲醇體積濃度分別為0.5%、1%、2%、3%、4%、5%,30 ℃、200 r/min 培養(yǎng),24 h 后取樣,測(cè)定酶活和菌體干重。
1.4.5 補(bǔ)加甲醇量對(duì)重組P.pastoris GS115 誘導(dǎo)階段的影響
無(wú)菌條件下,將BMGY 培養(yǎng)基中的全部菌體轉(zhuǎn)接入甲醇初始體積濃度為1%的50 mL/250 mL BMMY 培養(yǎng)基中,每隔24 h 分別補(bǔ)加比例為0.25%、0.5%、1.0%、1.5%、2.0%、2.5%的100%甲醇,30 ℃、200 r/min 培養(yǎng),每隔24 h 取樣,測(cè)定酶活和菌體干重。
1.4.6 裝液量對(duì)重組P.pastoris GS115 誘導(dǎo)階段的影響
無(wú)菌條件下,將BMGY 培養(yǎng)基中的全部菌體轉(zhuǎn)接入裝液量分別10%、20%、30%、40%(250 mL錐形瓶計(jì))的BMMY 培養(yǎng)基中,30 ℃、200 r/min培養(yǎng),每隔24 h 取樣,測(cè)定酶活和菌體干重。
1.4.7 誘導(dǎo)溫度對(duì)重組P.pastoris GS115 誘導(dǎo)階段的影響
無(wú)菌條件下,將BMGY 培養(yǎng)基中的全部菌體轉(zhuǎn)接入裝液量為20%的BMMY 培養(yǎng)基中,分別在25 ℃、28 ℃、30 ℃條件下誘導(dǎo),每隔24 h 取樣,測(cè)定酶活和菌體干重。
1.4.8 山梨醇添加量對(duì)重組P.pastoris GS115 誘導(dǎo)階段的影響
無(wú)菌條件下,將BMGY 培養(yǎng)基中的全部菌體轉(zhuǎn)接入山梨醇添加量分別為3 g/L、6 g/L、9 g/L、12 g/L 的BMMY 培養(yǎng)基中,25 ℃條件下誘導(dǎo),培養(yǎng)144 h 后取樣,測(cè)定酶活和菌體干重。
2 結(jié)果與分析
2.1 高拷貝轉(zhuǎn)化菌P.pastoris GS115 的篩選
首先通過(guò)不含組氨酸的基本培養(yǎng)基MD 對(duì)轉(zhuǎn)化子進(jìn)行初篩,未轉(zhuǎn)化的宿主P.pastoris GS115 在組氨酸脫氫酶位點(diǎn)(His4)有突變,不能在此平板上生長(zhǎng),只有整合成功的重組P.pastoris GS115 才能在MD 平板上生長(zhǎng)。
為提高篩選效率,得到高拷貝轉(zhuǎn)化子,通過(guò)YPD-G418 平板進(jìn)行復(fù)篩。收集MD 平板上的陽(yáng)性轉(zhuǎn)化子,依次經(jīng)0.25 mg/mL、0.5 mg/mL、1.0 mg/mL、2.0 mg/mL、3.0 mg/mL 的G418 平板進(jìn)行梯度篩選。
經(jīng)過(guò)多次轉(zhuǎn)化試驗(yàn)后,分別從2.0 mg/mL、3.0 mg/mL 的G418 抗性平板上挑取20 個(gè)轉(zhuǎn)化子,依次編 碼 為PKAGL-2-1~PKAGL-2-20,PKAGL-3-1~PKAGL-3-20。活化后進(jìn)行搖床誘導(dǎo)培養(yǎng),96 h 后取樣進(jìn)行酶反應(yīng):以100 g/L 麥芽糖(50 mmol/L,pH 7.0 的磷酸緩沖液配制)為底物在45 ℃反應(yīng)6 h,沸水浴10 min 滅酶。經(jīng)HPLC 進(jìn)樣檢測(cè)后,因產(chǎn)物黑曲霉糖轉(zhuǎn)化率較低,難以進(jìn)行準(zhǔn)確定量,而在前期反應(yīng)階段中,黑曲霉糖的轉(zhuǎn)化率與葡萄糖的轉(zhuǎn)化率呈正相關(guān),因此可簡(jiǎn)單的以體系中檢測(cè)到的葡萄糖轉(zhuǎn)化率作為篩選高拷貝轉(zhuǎn)化子的相對(duì)指標(biāo),結(jié)果如下頁(yè)表1 所示,選擇菌株P(guān)KAGL3-14繼續(xù)進(jìn)行后續(xù)試驗(yàn)。
2.2 重組P.pastoris GS115 產(chǎn)酶條件的研究
畢赤酵母的發(fā)酵采取分階段培養(yǎng),第一階段以甘油為碳源,主要目的是促進(jìn)菌體生長(zhǎng),實(shí)現(xiàn)菌體大量增殖,為后期發(fā)酵階段做準(zhǔn)備,根據(jù)Invitrogen的畢赤酵母操作手冊(cè),這一階段培養(yǎng)溫度一般設(shè)為30 ℃,無(wú)需優(yōu)化,但其他影響因素例如接種量、碳源濃度、初始pH 等對(duì)菌體生長(zhǎng)的影響需要做進(jìn)一步的研究;第二階段是甲醇誘導(dǎo)表達(dá)階段,此過(guò)程中,初始甲醇體積濃度、甲醇補(bǔ)加量、誘導(dǎo)溫度、轉(zhuǎn)接量、多碳源補(bǔ)充等因素均會(huì)對(duì)畢赤酵母的生長(zhǎng)和發(fā)酵產(chǎn)生不同程度的影響,需要逐一進(jìn)行研究。
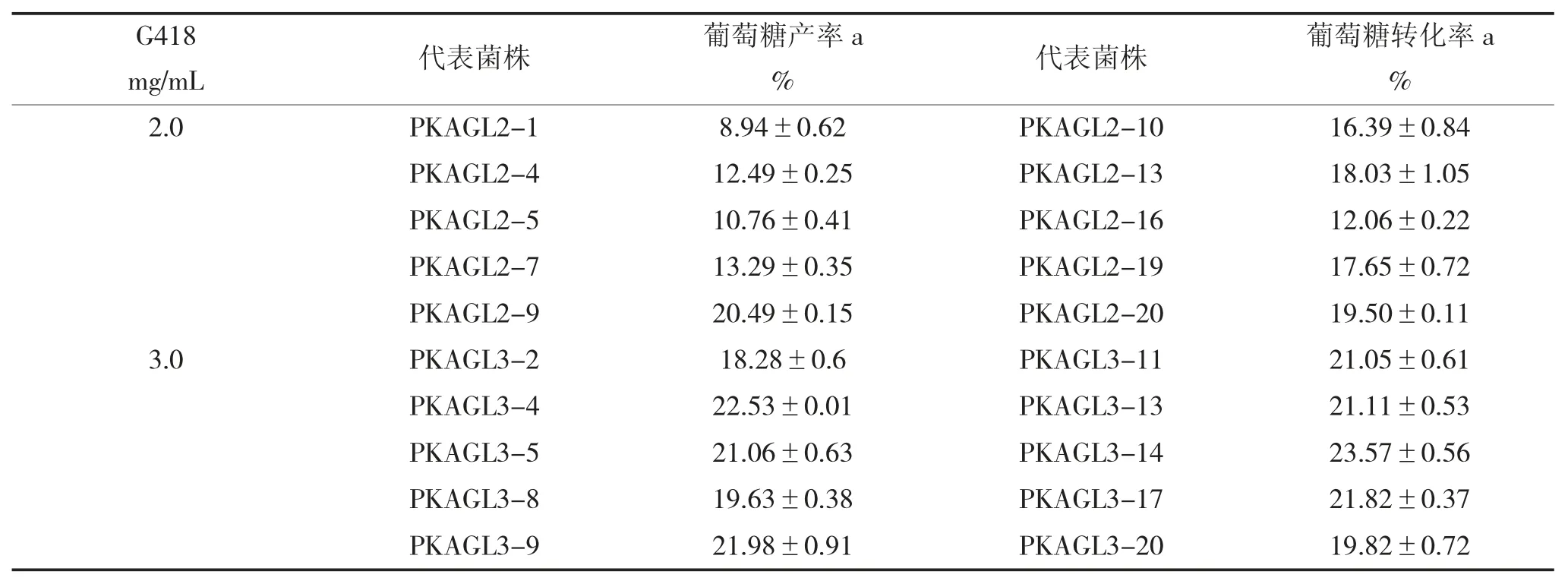
表1 AGL 基因高拷貝重組畢赤酵母的篩選
培養(yǎng)前期篩選的高拷貝轉(zhuǎn)化子PKAGL3-14,定時(shí)取樣測(cè)定酶活和菌體生長(zhǎng)量,繪制該重組菌生長(zhǎng)階段的生長(zhǎng)曲線和誘導(dǎo)階段的發(fā)酵曲線,見圖1。由圖1 可知,該重組菌的發(fā)酵周期較長(zhǎng),在轉(zhuǎn)接到發(fā)酵培養(yǎng)基的前2 天,菌體處于適應(yīng)階段,產(chǎn)酶量為0,之后隨著發(fā)酵時(shí)間延長(zhǎng),酶活逐漸增加,在第7 天達(dá)到最高值1.22 U/mL。
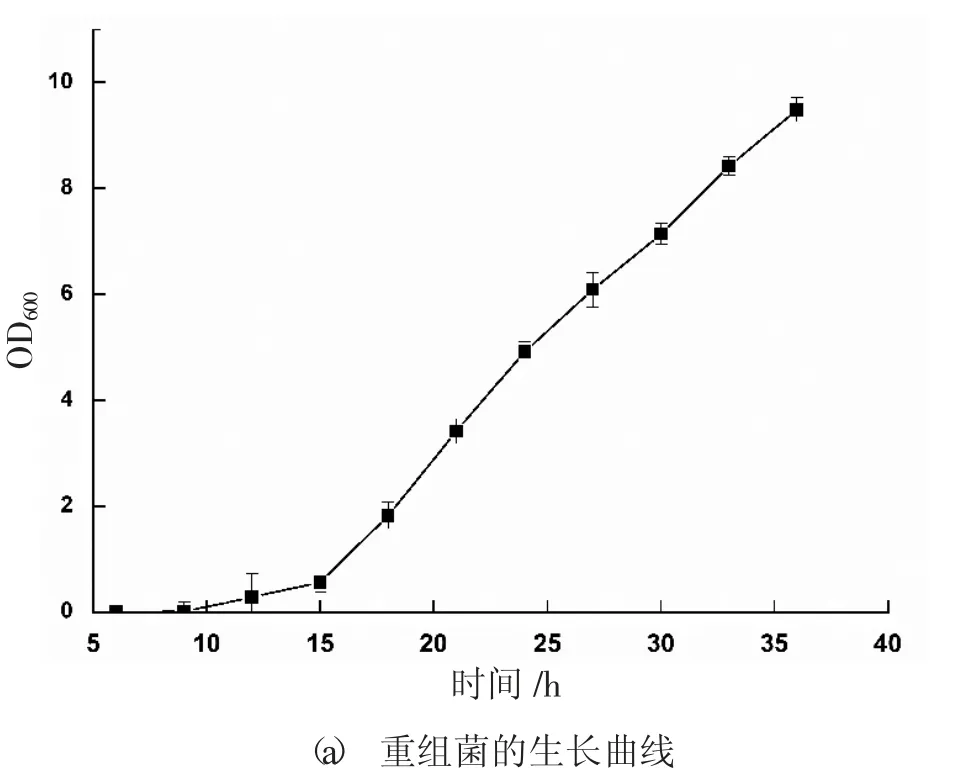
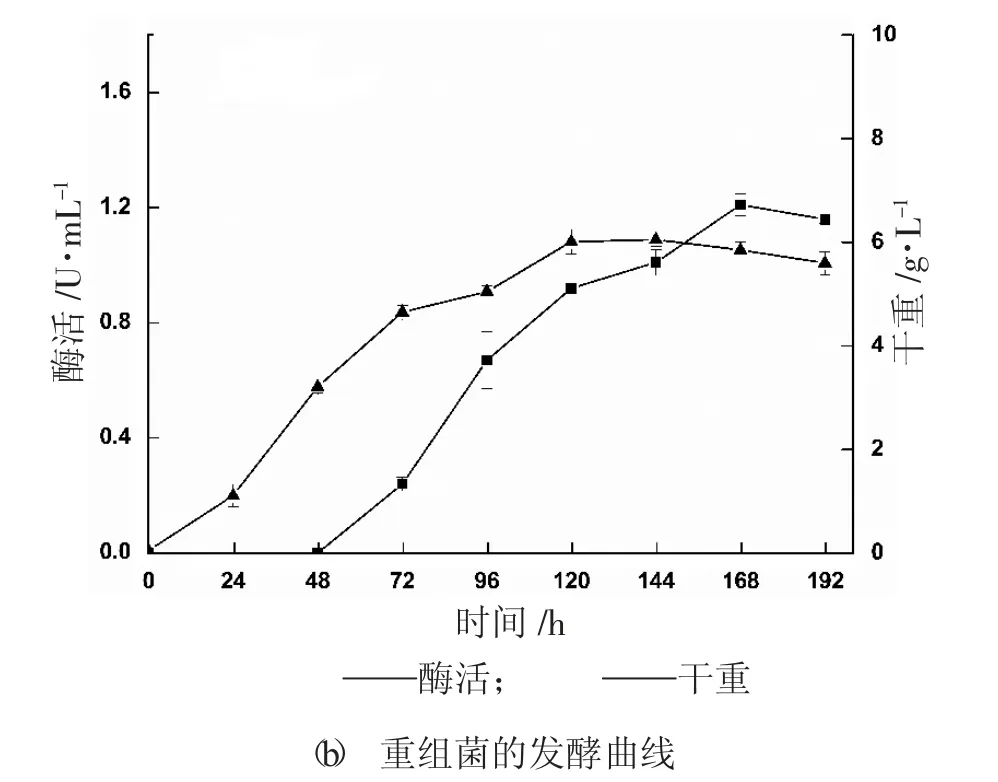
圖1 重組菌生長(zhǎng)及發(fā)酵曲線
2.2.1 接種量對(duì)重組P.pastoris GS115 生長(zhǎng)的影響
對(duì)于一個(gè)特定的生長(zhǎng)周期,接種量過(guò)小會(huì)延緩菌株到達(dá)對(duì)數(shù)期的時(shí)間,從而延長(zhǎng)整個(gè)發(fā)酵周期,過(guò)大的接種量則會(huì)在短時(shí)間內(nèi)消耗掉培養(yǎng)基中營(yíng)養(yǎng)成分。不同接種量對(duì)重組畢赤酵母生長(zhǎng)的影響結(jié)果見下頁(yè)圖2。
由圖2 可知,培養(yǎng)前2 h,菌體生長(zhǎng)緩慢,處于延滯期;12 h~30 h 期間,生長(zhǎng)速度均持續(xù)加快,處于指數(shù)增長(zhǎng)階段,該階段的菌體適宜于轉(zhuǎn)接到發(fā)酵培養(yǎng)基進(jìn)行誘導(dǎo)培養(yǎng);30 h 過(guò)后,1%、3%、6%接種量菌體生長(zhǎng)速度仍處于對(duì)數(shù)增長(zhǎng)期,10%、15%接種量的菌體生長(zhǎng)增速減緩,即將過(guò)渡到穩(wěn)定期。因此,選取10%為最適接種量。
2.2.2 甘油含量對(duì)重組P.pastoris GS115 生長(zhǎng)的影響
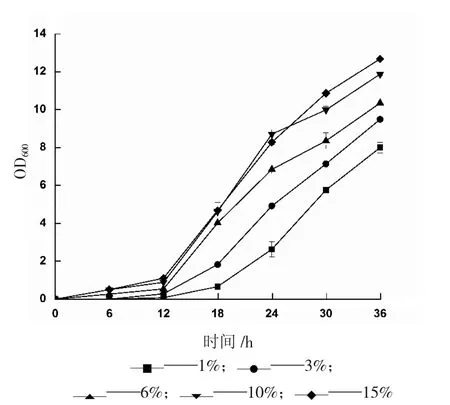
圖2 不同接種量對(duì)重組畢赤酵母生長(zhǎng)的影響
碳是微生物正常生命活動(dòng)所必需的元素,能為微生物生長(zhǎng)提供碳元素的營(yíng)養(yǎng)物質(zhì)均稱為碳源,一方面,碳源可以為微生物的碳架構(gòu)建以及細(xì)胞生長(zhǎng)提供碳成分,另一方面,也可以為微生物的生命活動(dòng)提供能量。
甘油作為酵母生長(zhǎng)階段的唯一碳源,是影響菌體生長(zhǎng)的關(guān)鍵因素。將重組菌接種至甘油含量不同的BMMY 培養(yǎng)基中,每隔3 h 取樣檢測(cè)該重組菌的生長(zhǎng)狀況,測(cè)定結(jié)果見圖3。
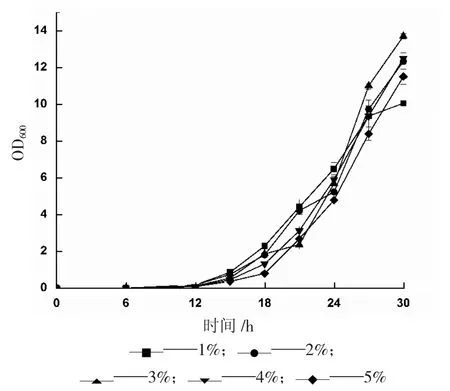
圖3 不同甘油濃度對(duì)重組畢赤酵母生長(zhǎng)的影響
由圖3 可知,在開始階段,甘油消耗較少,不同含量均能滿足菌體生長(zhǎng)的需要,因此對(duì)菌體量影響較小,之后隨著時(shí)間延長(zhǎng),差異逐漸明顯。甘油含量1%的培養(yǎng)基中菌體生長(zhǎng)速度最慢,也最早進(jìn)入平緩期,說(shuō)明少量的甘油被快速消耗,后期已不能滿足菌體生長(zhǎng)需要;含量3%的培養(yǎng)基中菌體生長(zhǎng)速度最快,甘油含量2%、4%、5%的培養(yǎng)基中菌體生長(zhǎng)趨勢(shì)差別不大,說(shuō)明過(guò)高的碳源含量一方面會(huì)對(duì)菌體造成負(fù)擔(dān),延緩它的生長(zhǎng)趨勢(shì),另一方面也會(huì)增加經(jīng)濟(jì)成本,因此,選取3%為最適甘油含量。
2.2.3 初始pH 值對(duì)重組P.pastoris GS115 生長(zhǎng)的影響
微生物所處環(huán)境的酸堿度對(duì)其生長(zhǎng)和繁殖有很大的影響,一方面,pH 可以通過(guò)激活或抑制某些特定酶的活性從而改變菌體的代謝途徑,進(jìn)而影響到菌體的生長(zhǎng);另一方面,pH 值可以通過(guò)改變細(xì)胞膜表面所帶電荷的狀態(tài)從而改變細(xì)胞膜的通透性,進(jìn)而影響到菌體對(duì)營(yíng)養(yǎng)物質(zhì)的吸收利用以及代謝產(chǎn)物的排放等;此外,pH 還可以通過(guò)改變?nèi)芤褐心承┪镔|(zhì)的離子化程度進(jìn)而影響到菌體對(duì)該物質(zhì)的吸收。
重組畢赤酵母菌在初始pH 值不同的BMMY 培養(yǎng)基中生長(zhǎng)狀況如圖4 所示。
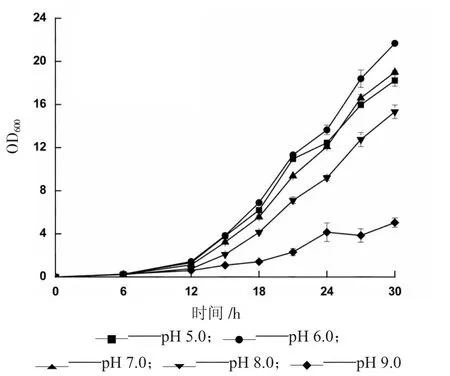
圖4 初始pH 值對(duì)重組畢赤酵母生長(zhǎng)的影響
由圖4 可知,當(dāng)初始pH 值為6.0 時(shí),菌體生長(zhǎng)狀態(tài)最好,pH 5.0 和7.0 條件下對(duì)菌體生長(zhǎng)狀況影響較小,菌體生長(zhǎng)量略有下降,但偏堿性環(huán)境(pH 8.0 和9.0)不適于菌體生長(zhǎng),生長(zhǎng)量大幅度下降,說(shuō)明菌體更偏好在弱酸性環(huán)境中生長(zhǎng),因此,選擇pH 6.0 為重組畢赤酵母生長(zhǎng)階段的初始pH值。
通過(guò)對(duì)畢赤酵母生長(zhǎng)階段影響因素的研究,確定了最佳的生長(zhǎng)階段培養(yǎng)條件為:接種量10%,碳源(甘油)含量3%,初始pH 值6.0。為進(jìn)一步確定合適轉(zhuǎn)接到誘導(dǎo)培養(yǎng)基BMMY 的種齡,在上述最優(yōu)培養(yǎng)條件下,測(cè)定了菌體在BMGY 培養(yǎng)基中的生長(zhǎng)曲線,如圖5 所示。
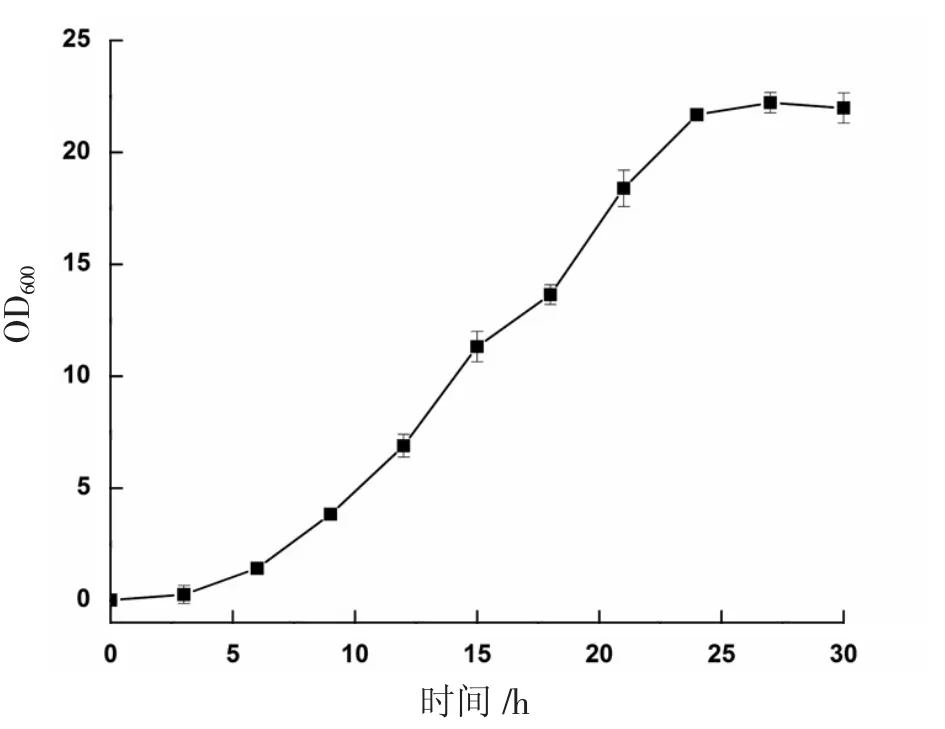
圖5 重組畢赤酵母的生長(zhǎng)曲線
由圖5 可知,在發(fā)酵前6 h,菌體生長(zhǎng)較慢,沒(méi)有明顯增長(zhǎng),為延滯期;6~24 h 期間,菌體生長(zhǎng)迅速,呈指數(shù)增長(zhǎng),為對(duì)數(shù)期;24 h 以后,菌體量基本維持恒定,菌體生長(zhǎng)進(jìn)入了穩(wěn)定期。因此,選取培養(yǎng)24 h 的菌體轉(zhuǎn)接入后續(xù)BMMY 培養(yǎng)基進(jìn)行發(fā)酵培養(yǎng)。
2.2.4 初始甲醇體積濃度對(duì)重組P.pastoris GS115 產(chǎn)酶的影響
在畢赤酵母的發(fā)酵過(guò)程中,甲醇既是碳源又是誘導(dǎo)劑,當(dāng)菌體由生長(zhǎng)階段過(guò)渡到發(fā)酵階段時(shí),需要對(duì)含有甲醇的生長(zhǎng)環(huán)境進(jìn)行適應(yīng),因此,確定合適的甲醇初始體積濃度至關(guān)重要,過(guò)低會(huì)使菌體處于饑餓狀態(tài),延緩菌體的生長(zhǎng),不能達(dá)到足夠的誘導(dǎo)效果,過(guò)高會(huì)對(duì)菌體產(chǎn)生毒害作用,抑制醇氧化酶的活性從而影響目的蛋白的表達(dá)甚至使菌體死亡。
初始甲酵體積濃度對(duì)α-葡萄糖苷酶的影響見圖6。
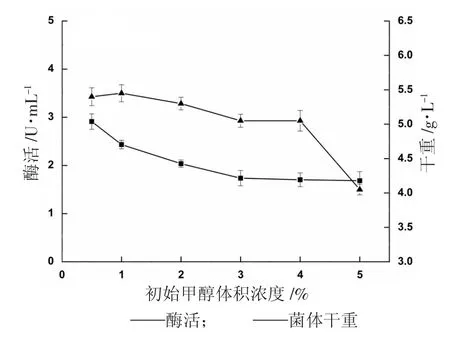
圖6 初始甲醇濃度對(duì)產(chǎn)α-葡萄糖苷酶的影響
圖6 反映了在甲醇初始濃度不同的BMMY 培養(yǎng)基中培養(yǎng)重組菌24 h 后的產(chǎn)酶情況,初始甲醇體積濃度1%的條件下,發(fā)酵24 h 酶活為2.912 U/mL,之后隨著甲醇體積濃度的升高而降低,當(dāng)甲醇體積濃度為5%時(shí),酶活已降低了42%,但菌體量之間差異不超過(guò)10%,說(shuō)明在誘導(dǎo)開始階段,菌體還處于適應(yīng)期,1%的甲醇已能滿足生長(zhǎng)和發(fā)酵產(chǎn)酶的需要,過(guò)高反而會(huì)影響目的蛋白的表達(dá),因此,甲醇初始體積濃度選定為1%。
2.2.5 補(bǔ)加甲醇量對(duì)重組P.pastorisGS115 產(chǎn)酶的影響
隨著發(fā)酵的持續(xù)進(jìn)行,初始添加的甲醇被不斷消耗,因此需要每隔24 h 補(bǔ)加一定量的甲醇,當(dāng)補(bǔ)加量在0.25%~2.5%范圍內(nèi)時(shí),結(jié)果如圖7 所示。當(dāng)補(bǔ)加甲醇量為0.5%時(shí),發(fā)酵96 h 酶活達(dá)到最高4.301 U/mL,補(bǔ)加量低于0.5%時(shí),甲醇無(wú)法同時(shí)完全滿足菌體生長(zhǎng)和誘導(dǎo)產(chǎn)酶的需求量,因此,雖然變化趨勢(shì)相同,但所能達(dá)到的最高酶活有所差異,當(dāng)甲醇體積濃度高于0.5%時(shí),會(huì)在更短的時(shí)間內(nèi)達(dá)到產(chǎn)酶的最大值,但由于過(guò)剩甲醇對(duì)于菌體的損傷作用,反而影響了目的蛋白的表達(dá)。
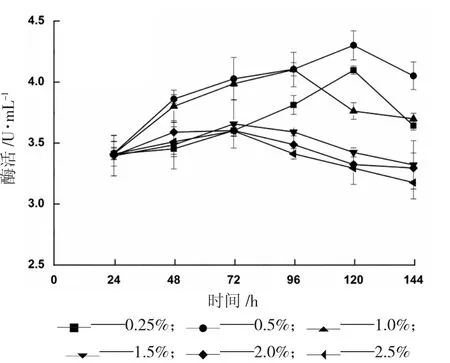
圖7 不同濃度甲醇補(bǔ)加量對(duì)產(chǎn)α-葡萄糖苷酶的影響
2.2.6 裝液量對(duì)重組P.pastoris GS115 產(chǎn)酶的影響
在搖瓶培養(yǎng)中,不同體積的裝液量會(huì)導(dǎo)致溶氧水平的差異,裝液量越少,傳氧系數(shù)越大。重組畢赤酵母的發(fā)酵過(guò)程需要足夠的氧氣激活體內(nèi)的過(guò)氧化氫酶體,進(jìn)而誘導(dǎo)外源重組蛋白的表達(dá),本試驗(yàn)研究了不同裝液量條件下對(duì)菌體產(chǎn)酶的影響,結(jié)果如圖8 所示,氧氣含量對(duì)重組菌發(fā)酵酶活有顯著的影響,裝液量逐漸加大時(shí)培養(yǎng)基中氧氣含量逐漸減少,菌體產(chǎn)酶效率降低,裝液量小于20%時(shí),菌體產(chǎn)酶較少,可能是因?yàn)榫w將大部分能量用于自身繁殖所致。裝液量大于20%時(shí),氧氣含量不足,菌體生長(zhǎng)與繁殖受到影響,產(chǎn)酶效率也很低;當(dāng)裝液量為20%時(shí),發(fā)酵酶活達(dá)到最高,為4.442 U/mL。因此,選取20%的裝液量為最適裝液量。
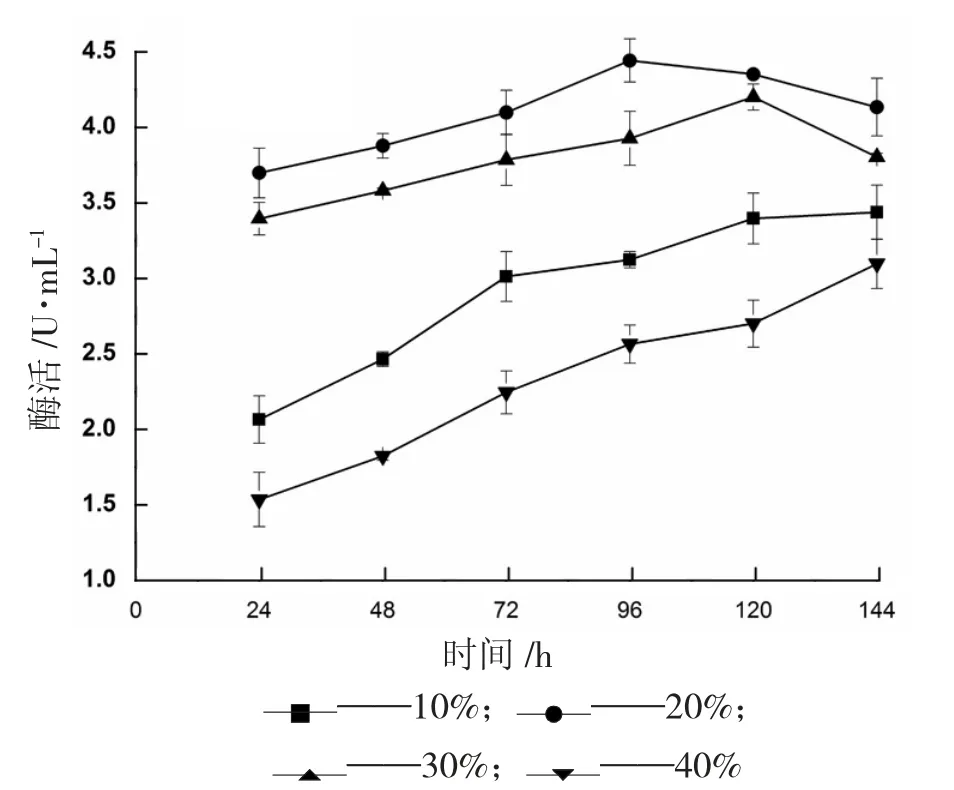
圖8 裝液量對(duì)產(chǎn)α-葡萄糖苷酶的影響
2.2.7 誘導(dǎo)溫度對(duì)重組P.pastoris GS115 產(chǎn)酶的影響
相較于最適生長(zhǎng)溫度30 ℃,已有多項(xiàng)研究表明,低溫有利于酵母重組外源蛋白的分泌表達(dá),低溫對(duì)其產(chǎn)酶的影響主要分為幾個(gè)方面:減輕甲醇對(duì)細(xì)胞的損傷作用,增加細(xì)胞存活率;延長(zhǎng)目的蛋白在內(nèi)質(zhì)網(wǎng)中的滯留時(shí)間,使得到更好的加工從而增加目的蛋白的正確折疊率;降低胞內(nèi)蛋白的活性,減少對(duì)目的蛋白的降解;促進(jìn)畢赤酵母自身的物質(zhì)和能量代謝,提高目的蛋白的表達(dá)速率;提高了胞內(nèi)AOX 基因的活性,進(jìn)而增加了甲醇的代謝速率,提升了目的蛋白的表達(dá)效率。
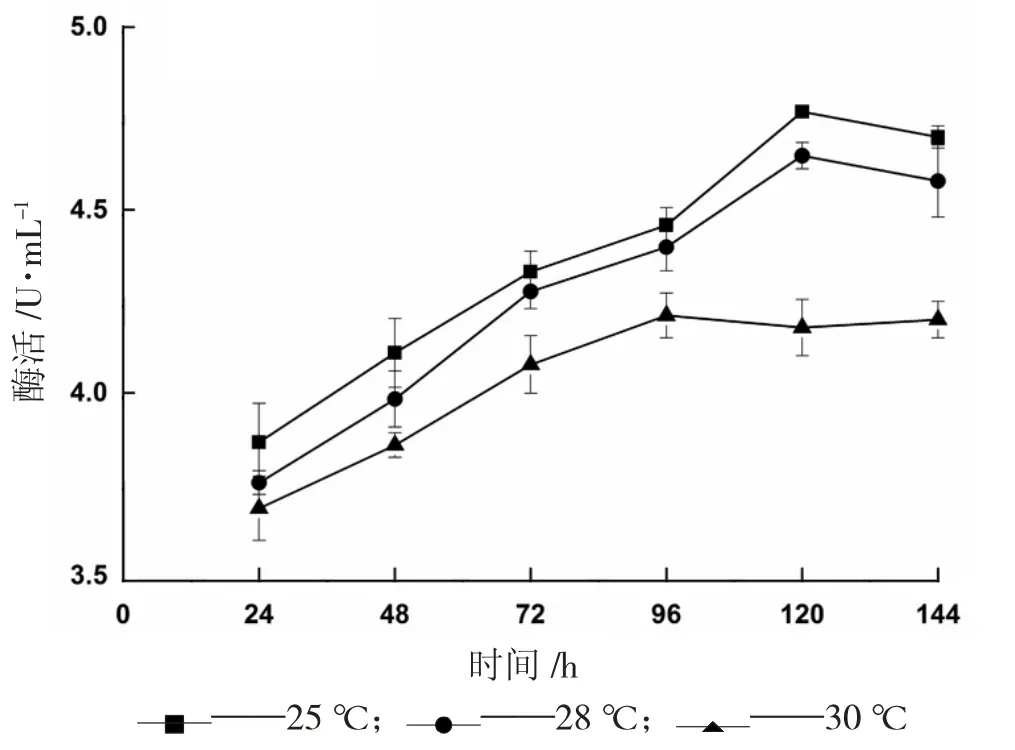
圖9 溫度對(duì)產(chǎn)α-葡萄糖苷酶的影響
為提升目的蛋白的表達(dá)效率,本試驗(yàn)設(shè)置了25 ℃、28 ℃、30 ℃3 個(gè)梯度進(jìn)行誘導(dǎo)表達(dá),結(jié)果如圖9 所示。溫度的降低雖然延長(zhǎng)了發(fā)酵過(guò)程中到達(dá)最高酶活的時(shí)間,但同時(shí)也提升了外源蛋白的表達(dá)效率,在25 ℃條件下誘導(dǎo)120 h,酶活達(dá)到最高值4.782 U/mL。此外,28 ℃條件下的產(chǎn)酶效果與25 ℃相比沒(méi)有明顯區(qū)別,因此若應(yīng)用于工業(yè)生產(chǎn),為降低成本,只需適當(dāng)降低溫度即可。
2.2.8 山梨醇添加量對(duì)重組P.pastoris GS115 產(chǎn)酶的影響
在畢赤酵母誘導(dǎo)階段,甲醇同時(shí)作為碳源和誘導(dǎo)劑,使得誘導(dǎo)階段酵母菌生長(zhǎng)速率較低,因此可以考慮采用復(fù)合碳源的方式提高菌體生長(zhǎng)速度,加快產(chǎn)酶速率。據(jù)報(bào)道,對(duì)于Muts 型重組子,山梨醇不會(huì)對(duì)AOX 基因造成抑制,不會(huì)抑制醇氧化酶的表達(dá),可用作甲醇誘導(dǎo)期的共同碳源,促進(jìn)畢赤酵母的生長(zhǎng),提高細(xì)胞活力,增強(qiáng)甲醇誘導(dǎo)的效果,減輕甲醇以及代謝中有毒副產(chǎn)物對(duì)菌體的毒性作用,最終達(dá)到促進(jìn)外源蛋白表達(dá)的目的。
山梨醇添加量對(duì)產(chǎn)α-葡萄糖苷酶的影響見圖10。
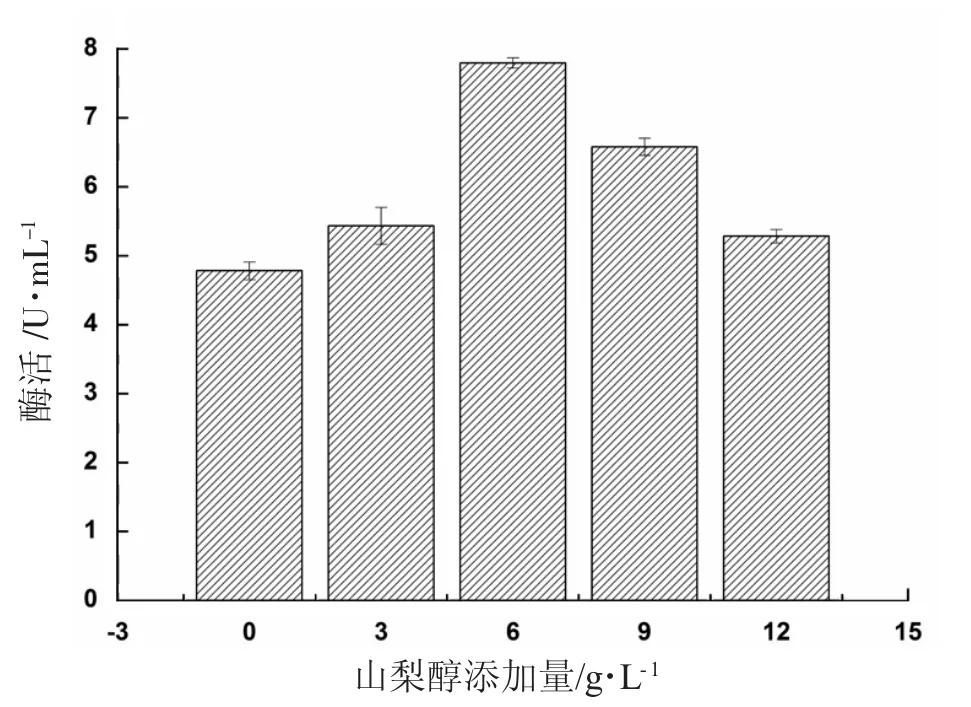
圖10 山梨醇添加量對(duì)產(chǎn)α-葡萄糖苷酶的影響
由圖10 可知,向誘導(dǎo)培養(yǎng)基中添加不同量的山梨醇,均可以提升發(fā)酵液中重組α-葡萄糖苷酶的表達(dá)量,當(dāng)添加量為6 g/L 時(shí),重組α-葡萄糖苷酶酶活達(dá)到最高值7.80 U/mL,但當(dāng)進(jìn)一步加大山梨醇的添加量時(shí),過(guò)多的山梨醇使得發(fā)酵液的滲透壓上升,阻礙了菌體的正常生長(zhǎng)與代謝產(chǎn)酶,使得酶活下降。因此,山梨醇的最適添加量為6 g/L。
3 結(jié)論
本研究實(shí)現(xiàn)了重組α-1,3-葡萄糖苷酶的畢赤酵母胞外表達(dá),與其他表達(dá)系統(tǒng)相比,畢赤酵母表達(dá)系統(tǒng)具有自身表達(dá)蛋白簡(jiǎn)單、操作簡(jiǎn)便以及遺傳穩(wěn)定等特性。此外,通過(guò)單因素試驗(yàn)對(duì)培養(yǎng)過(guò)程中接種量、碳源濃度、初始pH、培養(yǎng)溫度以及誘導(dǎo)過(guò)程中初始甲醇添加量、甲醇補(bǔ)加量、誘導(dǎo)溫度、裝液量等條件進(jìn)行了優(yōu)化,上清液中α-1,3-葡萄糖苷酶酶活由1.22 U/mL 提升至7.8 U/mL,提升了5.4 倍。目前關(guān)于α-1,3-葡萄糖苷酶的工程菌表達(dá)的報(bào)道還比較少,本試驗(yàn)為相關(guān)方面的研究提供了一定的數(shù)據(jù)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
