籽粒莧優質貯藏蛋白AhAMA1基因功能分析
2019-06-01 02:23:12趙熙寧宋亞楠季春麗崔紅利李潤植
山西農業科學 2019年5期
趙熙寧,張 飛,岳 敏,安 茜,宋亞楠,季春麗,崔紅利,李潤植
(山西農業大學分子農業與生物能源研究所,山西 太谷 030801)
籽粒莧(Amaranthus hypochondriacus L.)是莧科(Amaranthaceae)莧屬(Amaranthus)1年生植物,可作為糧、飼、藥、菜等多種用途的救荒作物[1-6]。籽粒莧富含油脂、蛋白質、碳水化合物和礦物質等,其中,種子油脂含量在10%以上,高于玉米、小麥和水稻等一般禾谷類作物,且油脂中角鯊烯等不飽和脂肪酸含量高達70%~80%[2],是一種健康型食用油脂資源;籽粒蛋白質含量大于30%,接近牛奶[3];種子貯藏蛋白品質好,沒有引起人體過敏反應的蛋白,8 種人體必需氨基酸含量均高于其他谷類作物籽粒[4]。因此,籽粒莧營養豐富,是一種營養均衡、適宜于所有人群的健康食源。此外,籽粒莧亦是一種優良的高產飼料作物,其莖葉柔嫩,適口性好,纖維素含量低,粗蛋白質含量為17.7%~27.1%,用作青貯飼料,營養價值較高。其青貯飼料和籽實可代替部分精飼料或餌料,用于畜禽及水產業[5-6]。
源于籽粒莧成熟種子的AMA1 蛋白是一種優質貯藏蛋白,其中,人體必需氨基酸含量高且均衡。該蛋白不會導致人體過敏反應,所含8 種人體必需氨基酸含量都高于FAO/WHO 建議的優質蛋白質標準。已有研究將編碼該蛋白的基因AhAMA1 應用于水稻籽粒遺傳改良,以提高稻米營養品質[7]。
本研究從籽粒莧(Amaranthus hypochondriacus L.)(cv.SX-04)發育種子中分離到 AMA1 基因編碼序列(AhAMA1),并檢測其時空表達譜;應用生物信息學工具系統分析AhAMA1 蛋白理化特性;進一步構建AhAMA1 基因組成型植物表達載體,并通過農桿菌介導滲透侵染法在本氏煙草(Nicotiana benthamiana)葉組織瞬時表達AhAMA1,檢測煙葉組織蛋白質、油脂、淀粉含量等生理生化性狀,旨在評估AhAMA1 基因是否可用于改良生物量大的植物營養器官蛋白含量和營養品質。
1 材料和方法
1.1 試驗材料
本研究所用的籽粒莧(Amaranthus hypochondriacus L.)種質(編號為 SX-04)與本氏煙草(Nicotiana benthamiana)種質材料均由山西農業大學分子農業與生物能源研究所提供。
1.2 試驗方法
1.2.1 籽粒莧不同組織總RNA 提取及AhAMA1基因表達量分析 以播種后40 d 籽粒莧盆栽苗的根、莖、葉以及不同發育時期種子(開花后7,12,17,22 d)為材料。按照Trizol 法提取各組織總RNA。-80 ℃保存備用。在http://biodb.swu.edu.cn/qprimerdb查找,獲得本氏煙草內參基因NtActin 引物NtActin-F,NtActin-R;獲得籽粒莧內參基因AHYPO 引物AHYPO-F,AHYPO-R。依據 GenBank 中籽粒莧(Amaranthus hypochondriacus)AhAMA1 基因轉錄本序列(Z11577.1)ORF(915 nt)設計 PCR 擴增引物:全長克隆引物AhAMA1-F1(加粗序列為XbaⅠ)、AhAMA1-R1(加粗序列為SmaⅠ);全長PCR 引物AhAMA1-F2,AhAMA1-R2;定量 PCR 引物 AhAMA1-F3,AhAMA1-R3(表1)。
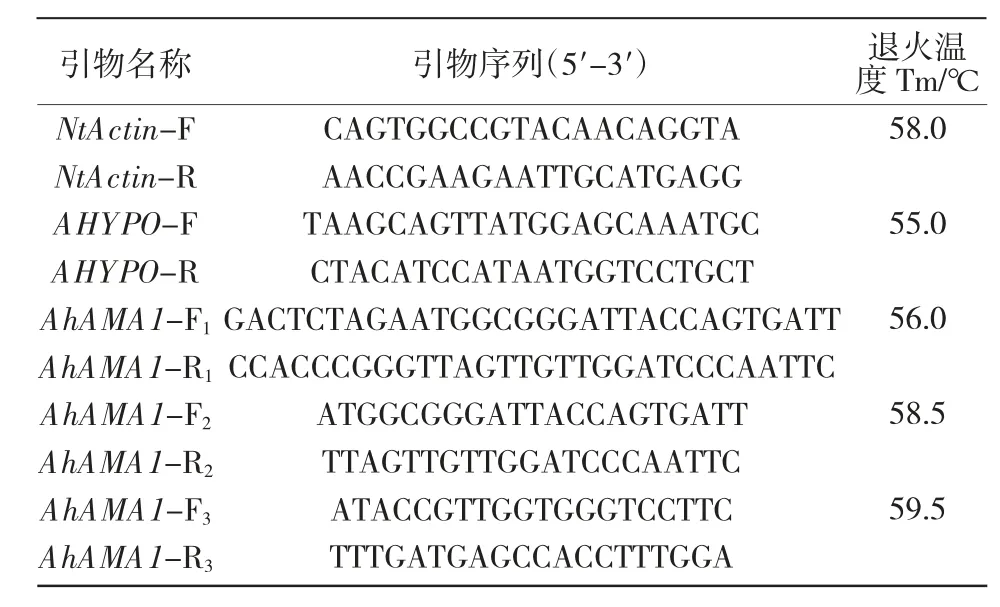
表1 PCR引物
1.2.2 AhAMA1 基因ORF 克隆 按照ABM 試劑盒使用方法,以經過濃度、純度測試的發育種子總RNA 為模板,反轉錄得到cDNA,-20 ℃保存備用。高保真PCR 反應體系參照說明書:Sterilized ddH2O(20 μL),2×San Taq PCR Mix(25 μL),DNA(1 μL),上游引物(10 μmol/L,2 μL),下游引物(10 μmol/L,2 μL)。反應條件為:95 ℃預變性 5 min;S1:95 ℃變性 30 s,S2:退火 Tm(表1)30 s,S3:72 ℃延伸 40 s,循環數(內參、定量×25)(克隆、全長×35)次;72 ℃,10 min。按照瓊脂糖凝膠DNA 回收試劑盒(Biomiga公司)操作說明書,將RT-PCR 擴增片段純化回收,然后連接到PBI121 表達載體上。XbaⅠ,SmaⅠ雙酶切驗證連接成功后,使用熱激法將重組質粒轉入到大腸桿菌(Escherichia coli)感受態DH5a 細胞中,涂布于LB 瓊脂培養基(含卡那霉素),37 ℃倒置培養。挑單克隆于LB 液體培養基擴大培養,PCR 檢測后將有目的條帶的菌落送至生工公司對重組質粒進行DNA 測序。
1.2.3 AhAMA1 蛋白理化特性的生物信息學分析通過NCBI 數據庫Blast 發現,籽粒莧AhAMA1(AAM09540.1)與籽粒莧凝集素(AAD33922.1)和尾穗莧(Amaranthus caudatus L.)凝集素(AAQ03084.1)氨基酸序列相似性較高,進一步通過DNAMAN對這3 個蛋白進行同源性分析。使用ProtParam(https://web.expasy.org/protparam/)軟件預測AhAMA1的氨基酸理論等電點、不穩定系數和總平均疏水性等理化性質;通過NCBI 預測分析AhAMA1 保守結構域;通過SignalP 分析AhAMA1 信號肽;通過NetPhos(http://www.cbs.dtu.dk/services/NetPhos/)分析AhAMA1 潛在磷酸化位點;通過SOPMA 軟件折疊預測分析AhAMA1 蛋白二級結構;應用SWISSMODEL 對AhAMA1 蛋白進行三維結構預測。
1.2.4 農桿菌介導煙草瞬時表達驗證AhAMA1 基因功能 將重組質粒AhAMA1-GFP-PBI121 以及PBI121 空載體通過熱激法轉入農桿菌(Agrobacterium)GV3101 中,在含有利福平和卡那霉素的LB瓊脂培養基上28 ℃恒溫倒置培養。挑取單克隆擴大培養后進行菌檢,將有目的條帶的農桿菌和含空載的農桿菌重懸后,采用滲透法侵染本氏煙草葉片。所侵染煙草葉片 3 d 后提取RNA,檢測A-hAMA1 基因是否表達;侵染5 d 后的煙葉樣品和對照煙葉樣品,經冷凍干燥后,用于測定蛋白質、油脂、淀粉、必需氨基酸含量等生理生化性狀。
2 結果與分析
2.1 籽粒莧不同組織器官AhAMA1基因表達分析


使用Trizol 法分別提取籽粒莧根、莖、葉以及不同發育時期種子總RNA。1%瓊脂糖凝膠電泳檢測結果顯示(圖1),各 RNA 樣品的 28S,18S,5S 條帶完整,質量符合試驗要求;半定量PCR 檢測各組織器官AhAMA1 基因表達表明(圖2),各組織中煙草內參基因AHYPO(a)表達量一致(條帶亮度相同),AhAMA1 基因(b)表達量有所差異(條帶亮度不同),揭示AhAMA1 基因在所測各組織中均有表達,其中在發育種子中高量表達。
2.2 AhAMA1基因ORF克隆
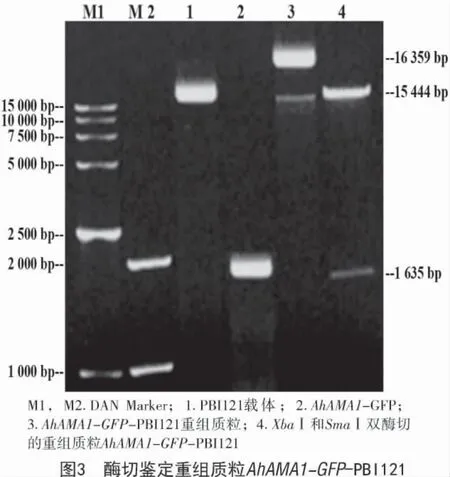
以籽粒莧(cv.SX-04)發育種子RNA 反轉錄獲得的cDNA 為模板,用分別帶有XbaⅠ和SmaⅠ酶切位點的目的基因引物進行高保真PCR,擴增AhAMA1 基因ORF,擴增的靶基因片段AhAMA1(915 bp)與 GFP(720 bp)基因連接后經 XbaⅠ和SmaⅠ雙酶切后電泳檢測結果如圖3所示,PCR 擴增的條帶與預期目的基因條帶(1 635 bp)相符。經測序驗證,該PCR 擴增產物序列與AhAMA1 基因ORF 序列完全一致。
2.3 AhAMA1基因植物組成型表達載體的構建
經XbaⅠ和SmaⅠ雙酶切后的AhAMA1-GFP與經XbaⅠ和SmaⅠ雙酶切的PBI121 表達載體進行連接獲得重組質粒,再用重組質粒轉化大腸桿菌感受態DH5a,獲得陽性菌落。提取質粒,用XbaⅠ和SmaⅠ進行雙酶切鑒定以及DNA 測序,結果如圖3所示,AhAMA1 基因 ORF-GFP 亞克隆到 PBI121 載體的多克隆位點,獲得了AhAMA1 基因組成型植物表達載體AhAMA1-GFP-PBI121(圖4)。

2.4 AhAMA1蛋白的生物信息學分析

使用NCBI 對AhAMA1 基因序列進行分析可知,AhAMA1 cDNA 全長為1 183 bp,起始密碼子位于cDNA 的47 bp 處,終止密碼子位于961 bp 處,開放閱讀框(ORF)為915 bp,編碼304 個氨基酸,其所編碼的蛋白命名為AhAMA1 蛋白(AAM09540.1)。將其與籽粒莧凝集素(AAD33922.1)和尾穗莧凝集素(AAQ03084.1)進行氨基酸序列多重比對顯示,其蛋白序列一致性高達99.12%,親緣關系很近(圖5)。
應用ProtParam 對AhAMA1 蛋白理化性質進行預測和分析,結果表明,其分子式為C1582H2406N412O461S12,原子總數為 4 873,相對分子質量為 34.96 ku。AhAMA1 蛋白由304 個氨基酸組成,其中,Arg+Lys(帶正電)29 個、Asp+Glu(帶負電)30 個,pI 值6.65;總平均親水性-0.343,是親水蛋白;不穩定指數值32.78,屬于穩定蛋白。
應用NCBI 相關工具對AhAMA1 蛋白保守結構域預測分析表明,該蛋白屬于Agglutinin 超家族(圖6)。SignalP 分析顯示,AhAMA1 蛋白不具有信號肽。NetPhos 潛在磷酸化位點分析顯示,AhAMA1的磷酸化位點有絲氨酸(11 個)、蘇氨酸(7 個)和酪氨酸(8 個)3 種。

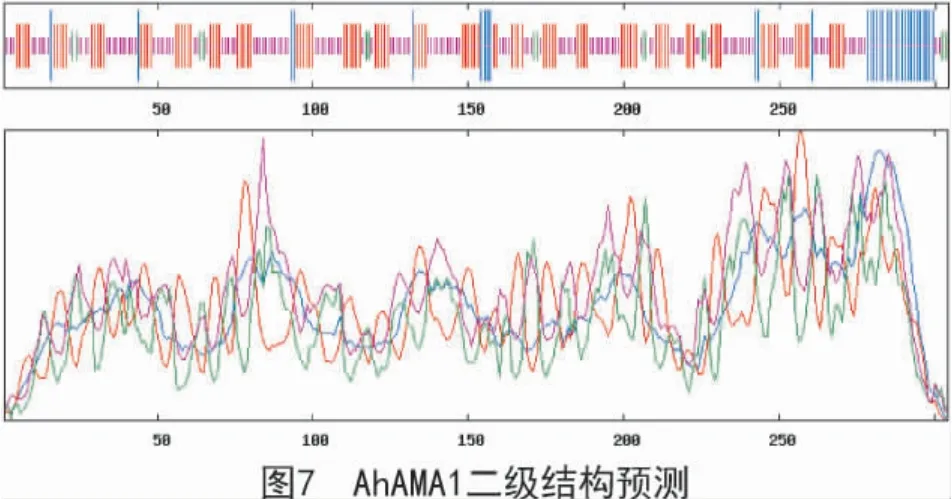
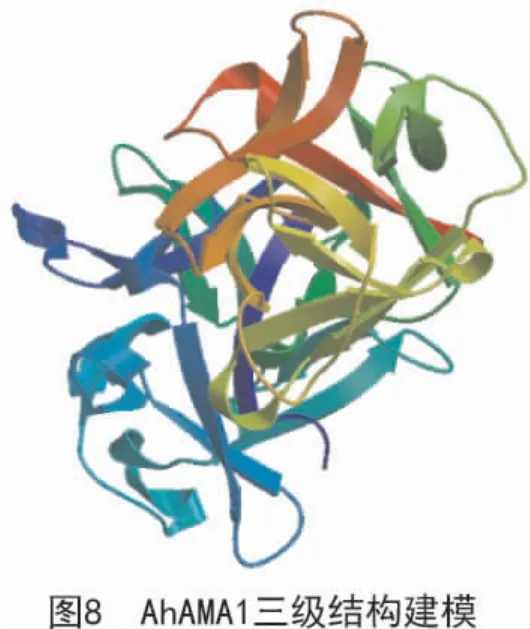
由SOPMA 軟件折疊預測分析得出(圖7),該蛋白二級結構由α 螺旋(藍色區域)、伸展鏈(紅色區域)、β 轉角(綠色區域)和無規則卷曲(紫色區域)組成,且占比分別為11.51%,36.18%,4.93%和47.37%。應用SWISS-MODEL 軟件對AhAMA1 蛋白進行三維結構圖建模(圖8),結果表明,該蛋白為單體,無配體;該蛋白的氨基酸序列與模板序列的一致性達到61%;X-射線晶體衍射發現,該蛋白是同型二聚體。
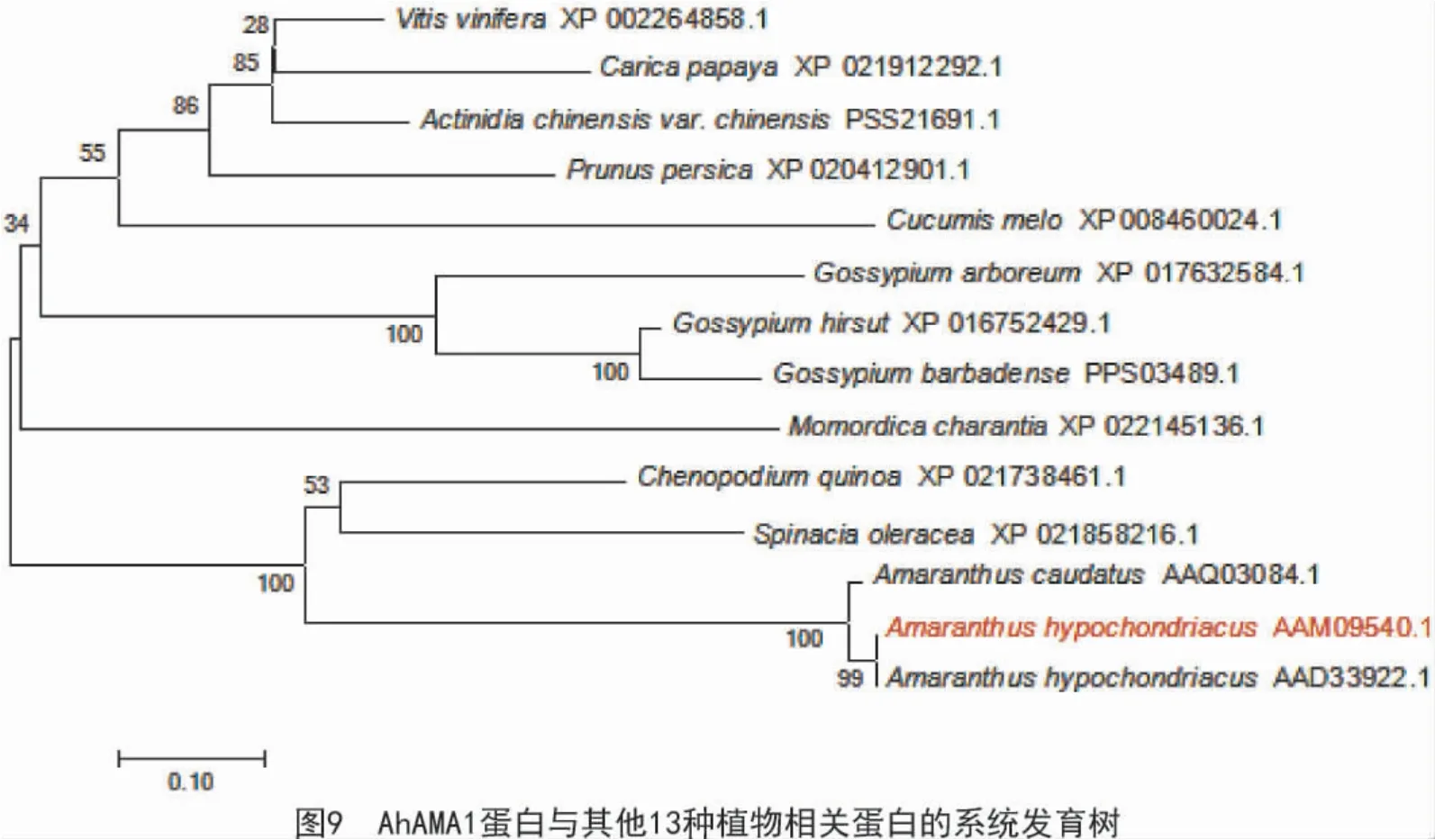
通過MEGA7 軟件構建AhAMA1 蛋白(AAM09 540.1)與其他植物相關蛋白的系統進化(圖9)結果顯示,AhAMA1 蛋白與籽粒莧凝集素親緣關系較近,這些蛋白包括籽粒莧凝集素(AAD33922.1)、尾穗莧凝集素(AAQ03084.1)、藜麥(Chenopodium quinoa willd)蛋白(XP_021738461.1)、葡萄(Vitis vinifera L.)蛋白(XP_002264858.1)、獼猴桃(Actinidia)蛋白(PSS21691.1)、番木瓜(Carica Papaya L.)蛋白(XP_021912292.1)、碧桃(Amygdalus persica L.var.persica f.duplex Rehd.)蛋白(XP_020412901.1)、菠菜(Spinacia oleracea L.)蛋白(XP_021858216.1)、甜瓜(Cucumis melo)蛋白(XP_008460024.1)、陸地棉(Gossypium hirsutum Linn.) 蛋白(XP_016752429.1)、海島棉(Gossypium barbadense Linn.)蛋白(PPS03489.1)、木本棉(Gossypium arboreum(A2))蛋白(XP_017632584.1)和苦瓜(Momordica charantia L.)蛋白(XP_022145136.1)。
2.5 農桿菌介導AhAMA1基因在煙葉組織瞬時表達分析

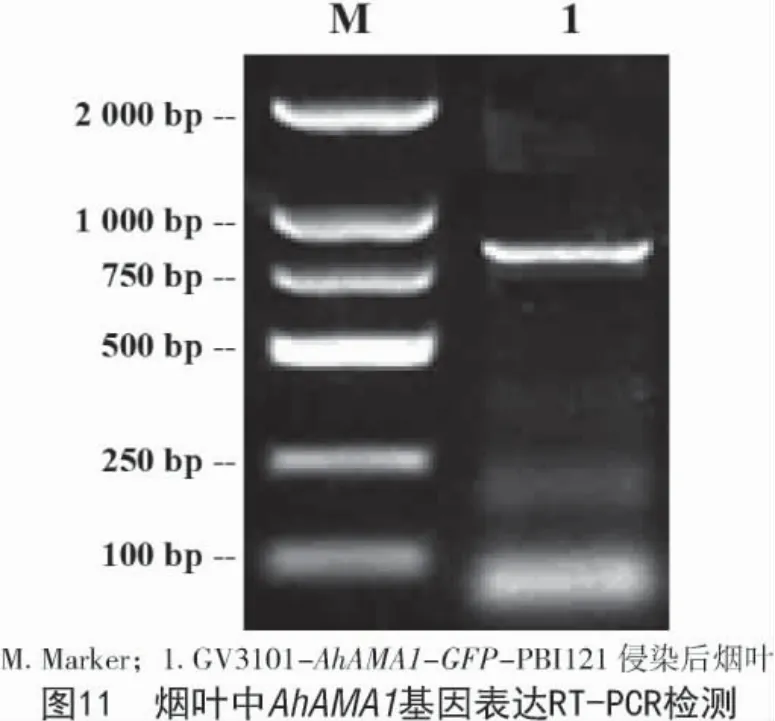
將重組表達質粒 AhAMA1-GFP-PBI121 和PBI121 空載分別通過熱激法轉化導入農桿菌GV3101 中,在含有利福平和卡那霉素的LB 瓊脂培養基上28 ℃恒溫倒置培養,挑取單克隆擴大培養后進行菌檢。將有目的條帶和含空載的農桿菌重懸后侵染煙草葉片。選用侵染2 d 后的煙草葉片提取RNA 檢測,檢測AhAMA1 基因是否成功表達,結果如圖10,11 所示,AhAMA1 基因在煙葉組織中成功表達。
2.6 瞬時表達AhAMA1基因的煙葉油脂、蛋白質和淀粉積累分析

選取侵染5 d 后的煙葉樣品,冷凍干燥后,萃取并測定蛋白質、油脂以及淀粉含量。如圖12 所示,野生型煙草及對照葉片蛋白含量為11.19%,瞬時表達AhAMA1 基因的煙草葉片蛋白含量為22.39%,是野生型煙草的2 倍;油脂及淀粉含量檢測相比野生型有所降低;必需氨基酸含量相比野生型均有增加(表2),異亮氨酸、亮氨酸、賴氨酸、甲硫氨酸、苯丙氨酸、蘇氨酸、色氨酸、纈氨酸相比野生型分別增長 42%,53%,33%,48%,47%,11%,14%,21%。

表2 煙葉中必需氨基酸質量分數 mg/g
3 結論與討論
農作物及飼草植物蛋白質含量及氨基酸組成決定其蛋白營養品質和用途。分離一些編碼優質蛋白質的基因,應用于農作物等植物的遺傳改良以提高其營養價值,近年來取得了一些可喜進展。ZHENG 等[8]利用PEG 法分別將來源于豌豆(Pisum sativum Linn)和菜豆(Phaseolus vulgaris Linn)的球蛋白基因整合到水稻基因組中,結果表明,菜豆球蛋白占總內胚乳蛋白質的4%。張秀君等[9-11]將馬鈴薯(Solanum tuberosum L.)花粉特異水溶性蛋白的cDNA 用基因槍法轉入玉米(Zea mays L.)中,結果顯示,獲得的轉基因玉米成熟種子蛋白含量提高,賴氨酸提高10%~54%。王為民等[12]將馬鈴薯高賴氨酸基因用基因槍法轉入水稻(Oryza sativa)龍特甫B和中優早5 號,結果顯示,龍特甫B 種子中蛋白質含量提高27.69%、賴氨酸含量提高16.15%;中優早5 號種子中蛋白質含量提高35.18%、賴氨酸含量提高了45.09%。高越峰等[13]將四棱豆(Psophocarpus tetragonolobus(L.)DC.)高賴氨酸蛋白基因 Lys 通過基因槍法轉入水稻,結果顯示,轉基因植株葉片中賴氨酸含量均提高,最高增幅16.04%。王逸群等[14]將從辣椒(Capsicum annuum L.)成熟花粉中克隆的高賴氨酸蛋白基因CFLR 通過農桿菌介導法轉入谷稈兩用水稻,結果發現,9 個轉基因植株葉片的賴氨酸含量均有提高,最高增幅22.71%。蔣家煥等[15]將四棱豆Lys 基因用農桿菌介導法轉入水稻臺粳9 號,并獲得了轉基因抗性植株,野生水稻種子賴氨酸含量僅為0.27%,而轉基因水稻種子為0.349%,相比野生型提高了29.3%;轉基因水稻種子蛋白含量提高了27.4%,轉基因水稻秸稈蛋白含量提高了30.9%。孔維文等[16]將抗蟲基因半夏(Pinellia ternata)凝集素基因PTA 和馬鈴薯的高賴氨酸基因SB401通過農桿菌介導法同時轉入雜交水稻1826 中,獲得了含PTA,SB401 基因的轉基因水稻植株。吳超等[17]將四棱豆高賴氨酸基因通過農桿菌介導法轉入脆莖粳稻中,結果發現,轉基因水稻秸稈中賴氨酸含量最高為0.350%,粗蛋白含量最高為7.27%。李科等[18]將馬鈴薯中高賴氨酸蛋白基因SB401 通過農桿菌介導法導入優質水稻品種日本晴中,結果發現,9 個純合的T4水稻轉化株系種子的蛋白質、賴氨酸和其他氨基酸含量均提高。MOMMA 等[19]將大豆球蛋白基因通過電擊法轉入水稻,結果發現,轉基因水稻種子中大豆球蛋白含量為40~50 mg/g,占種子總蛋白含量的4%~5%。這些研究表明,轉基因提高植物蛋白含量和改良蛋白營養品質是可行的,但不同蛋白基因異源表達產生的效果不同。
許明等[20]采用 RT-PCR 方法,從籽粒莧(cv.千穗谷1 號)幼嫩種子克隆出AMA1 基因的完整開放閱讀框序列,結果表明,該序列與GenBank 登錄的籽粒莧AMA1 基因的核苷酸序列只有一個堿基的差異,但是不影響所編碼氨基酸,原核表達獲得了相應的目標蛋白;進一步將AMA1 基因在水稻胚乳中特異性表達,可一定程度提高水稻種子3 種限制性必需氨基酸含量。彭雪娟[7]用胚乳特異性啟動子驅動AMA1 基因在水稻種子中表達,改進了稻米的蛋白營養品質。TAMáS 等將[21]AMA1 基因通過基因槍轉化法成功導入小麥中,結果使小麥氨基酸含量提高,而且面粉中有關面粉質量的功能參數得以改善。
本研究從籽粒莧種植材料(cv.SX-04)發育種子分離到 AhAMA1 的 ORF 序列(916 bp),與 Gen Bank 登錄的籽粒莧AMA1 基因序列完全一致;AhAMA1 編碼304 個氨基酸,分子量為34.96 ku。表達分析揭示該基因在發育種子后期高表達,與籽粒莧種子蛋白質快速富集期相吻合。成功構建了AhAMA1的組成型植物表達載體AhAMA1-GFP-PBI121,并通過農桿菌介導侵染使其在本氏煙葉組織中瞬時表達。煙葉生化測試顯示,AhAMA1 瞬時表達導致煙葉組織中蛋白質含量顯著提高,含量達22.39%,是野生型煙草葉片蛋白含量的2 倍;8 種必需氨基酸均有顯著增加,且對油脂及淀粉含量影響未達顯著水平。
總之,本研究表明,源于籽粒莧編碼優質貯藏蛋白(8 種必需氨基酸含量高且均衡)的AhAMA1基因可以應用于高生物量植物營養器官的蛋白營養品質遺傳改良,從而提高這些蛋白及8 種必需氨基酸用于功能飼料/餌料或加工其他產品的市場價值。
