紫蘇二酰甘油酰基轉移酶基因(PfDGAT1)鑒定及表達分析
2019-06-01 02:23:06周雅莉任文燕郝月茹段露露李潤植王計平
山西農業科學 2019年5期
關鍵詞:分析
周雅莉,任文燕,郝月茹,安 茜,段露露,李潤植,王計平
(1.山西農業大學農學院,山西 太谷 030801;2.山西農業大學分子農業與生物能源研究所,山西 太谷 030801)
紫蘇(Perilla frutescens(L.)Britt.),唇形科紫蘇屬,為1年生草本植物[1],是一種具有廣泛開發利用前景的經濟作物,是我國衛生部首批頒布的既是藥品又是食品的60 種“藥食同源”植物之一[2],被廣泛用作中藥材和蔬菜、調味品等[3]。紫蘇種子中含油量可達 35%~64%,而 ω-3 脂肪酸(C18∶3)和 ω-6脂肪酸(C18∶2)含量分別高達54%~64%和14%,是目前所發現的C18∶3 脂肪酸含量最高的特殊油料作物之一[4-7]。
二酰甘油酰基轉移酶(Diacylgycerol acyltransferase,DGAT)是三酰甘油(Triacylglycerol,TAG)合成的關鍵酶和限速酶,主要催化二酰甘油加上脂肪酸酰基生成三酰甘油[8]。對于大多數油料作物種子的TAG 合成積累具有重要作用。有研究發現,油料植物在其種子發育過程中,隨著DGAT 酶活性不斷加強油脂積累速度加快,而當油脂含量趨于穩定時,DGAT 活性會逐漸降低[9]。KUBIS 等[10]研究發現,在大豆(Glycine max)高油和低油品種中,大豆種子油脂積累速度、油脂含量均與DGAT 酶的活性呈正相關。HOBBS 等[11]首先在擬南芥(Arabidopsis thaliana)中克隆得到DGAT1 基因,且該基因的cDNA 在擬南芥中僅存在一個拷貝。隨后,人們相繼在油菜(Brassica napus)[12]、煙草(Nicotiana tabacum)[13]、大豆[14]、蓖麻 (Ricinus communis)[15]、旱金蓮(Tropaeolum majus)[16]和玉米(Zea mays)[17]等植物中克隆得到DGAT1 基因。
本研究依據紫蘇轉錄組數據庫,從晉紫蘇1 號中克隆得到PfDGAT1 基因全長序列,并分析該基因在紫蘇不同組織中的表達特性,旨在為進一步研究PfDGAT1 基因在紫蘇等油料作物油脂合成代謝過程中的作用提供理論基礎。
1 材料和方法
1.1 試驗材料
供試紫蘇品種晉紫蘇1 號種植于山西農業大學農學院試驗站。取不同組織器官及不同發育時期(開花后 10,20,30,40 d)的種子,液氮速凍后 -80 ℃保存備用。大腸桿菌DH5α 感受態與試驗所用克隆載體pMD18-T 均保存于山西農業大學分子農業與生物能源研究所。
1.2 試驗方法
1.2.1 目的基因克隆 利用EASYspin RNA 提取試劑盒(RN09,艾德萊生物)提取晉紫蘇1 號葉片總RNA,參考ABM 公司反轉錄試劑盒(5X All-In-One MasterMix(with AccuRT Genomic DNA Removal Kit))方法進行反轉錄,得到cDNA 用于后續試驗。依據GeneBank 中紫蘇 PfDGAT1 基因序列(AF298815.1)設計擴增全長cDNA 的引物PfDGAT1-F/R 進行PCR 擴增(表1)。反應體系為20μL:模板cDNA2μL,上下游引物各 0.5 μL,10×Pfu Buffer 2 μL,dNTP Mixture(2.5 mmol/L)1.6 μL,Pfu DNA Polymerase 0.5 μL,dd H2O 12.9 μL。反應條件為:94 ℃預變性3 min;94 ℃變性 30 s,58 ℃退火 30 s,72 ℃延伸2 min,循環 30 次;72 ℃終延伸 5 min。1%瓊脂糖凝膠電泳檢測。將PCR 擴增產物純化回收并連接到pMD18-T 載體上,轉入大腸桿菌感受態細胞DH5α中,鑒定陽性克隆,送至通用生物公司(安徽)測序。
1.2.2 PfDGAT1 基因生物信息學分析 利用NCBIORF(Open Reading Frame,ORF)在線軟件對 1.2.1獲得的基因序列的開放閱讀框進行識別,并將核酸、氨基酸進行比對[18]。利用在線軟件ProtParam(http://web.expasy.org/protparam/)對該蛋白質的基本理化性質進行分析;使用PSORTII(http:// psort.hgc.jp/form2.html)軟件對該蛋白質進行亞細胞定位預測;運用TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)在線軟件分析該蛋白質的跨膜區域,通過SOPMA(http://npsa-prabi.ibcp.fr/cgi-bin/npsa_autom at.pl?page=npsa_sopma.html)對其二級結構進行預測;利用NCBI-CDD(http://ncbi.nlm.nih.gov/Structure/cdd/Wrpsb.cgi)預測該蛋白的功能結構域;采用SWISSMODEL(http://swissmodel.expasy.org/)在線軟件對該蛋白進行三維結構分析與建模。運用在線軟件PRALINE(http://www.ibi.vu.nl/programs/praline-www/)對紫蘇DGAT1 蛋白與其他已公布的植物蛋白質進行多序列比對,從中找出保守域。運用MEGA 7.0 多序列比對軟件對紫蘇DGAT1 蛋白與其他多種高等植物氨基酸序列進行比對,并構建系統進化樹。
1.2.3 實時熒光定量PCR 檢測 提取晉紫蘇1 號的根、莖、葉、花及不同發育時期(開花后10,20,30,40 d)種子的總RNA,反轉錄成cDNA。根據紫蘇PfDGAT1 基因的cDNA 序列設計熒光定量PCR 特異性引物PfDGAT1-q-F/R,以紫蘇18S rRNA(表1)作為內參基因[19]設計引物18S rRNA-F/R,用于實時熒光定量PCR 檢測。熒光定量PCR 反應體系為10 μL:cDNA 模板 0.5 μL,EvaGreen 2X qPCR MasterMix 5 μL,正反引物各 0.3 μL,ddH2O 3.9 μL。擴增程序為:95 ℃ 10 min;95 ℃ 15 s,58 ℃ 60 s,40 次循環;65 ℃ 5 s,95 ℃ 15 s。每個樣品進行 3 次生物學重復和3 次技術重復。采用2-ΔΔCt法計算紫蘇PfDGAT1 基因的相對表達量[20]。
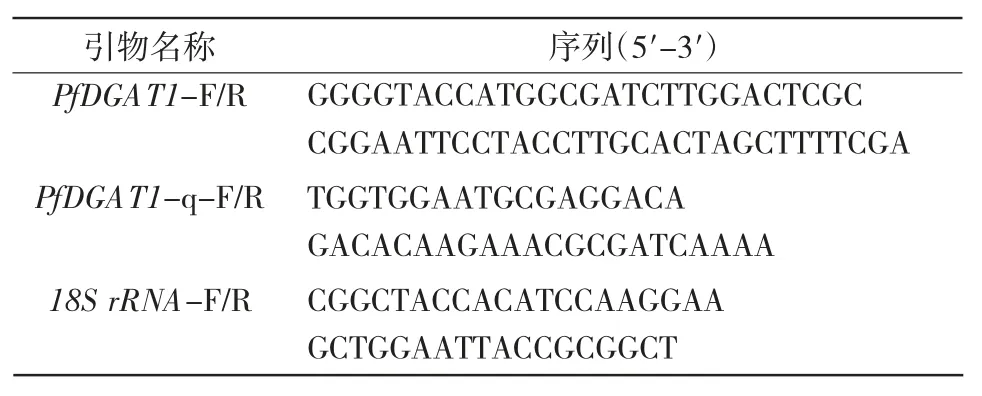
表1 引物信息
1.3 數據分析
試驗數據采用SPSS 軟件進行統計分析。
2 結果與分析
2.1 紫蘇PfDGAT1基因全長cDNA序列的克隆
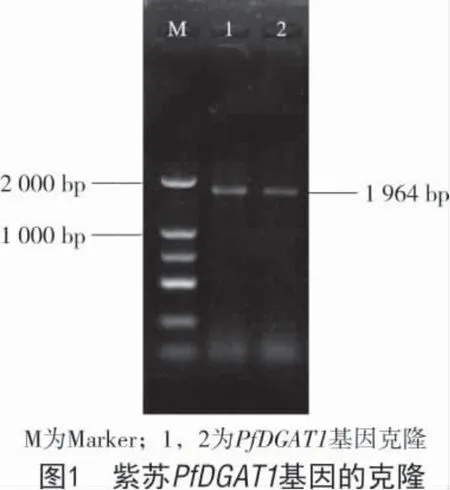
以晉紫蘇1 號葉片cDNA 作為模板進行RTPCR 擴增,擴增產物電泳結果如圖1所示,RT-PCR反應擴增得到1 條長度約為1 964 bp 的目的條帶,與預期相符。將編碼區序列與GeneBank 中紫蘇PfDGAT1 基因(AF298815.1)序列進行比對,結果表明,推測的氨基酸序列與已公布PfDGAT1 基因序列(AF298815.1)無差異,說明分離獲得了正確的PfDGAT1 基因的 cDNA 克隆。
2.2 PfDGAT1基因的生物信息學分析
本試驗克隆獲得PfDGAT1 基因序列全長為1 964 bp,開放閱讀框(ORF)長度為 1 605 bp(核苷酸序列位于69~1 673),共編碼534 個氨基酸殘基(圖2)。通過ProtParam 軟件預測分析表明,PfDGAT1蛋白的分子式為C2812H4322N714O747S35,相對分子質量為61.21 ku,理論等電點(pI)為8.34,不穩定系數為46.77,親水性系數為0.227,推導該蛋白質為疏水性不穩定蛋白。經PSORTII 亞細胞定位預測分析顯示,紫蘇PfPDAT1 蛋白定位于質膜的可信度為65.2%,預測其為膜錨定蛋白。
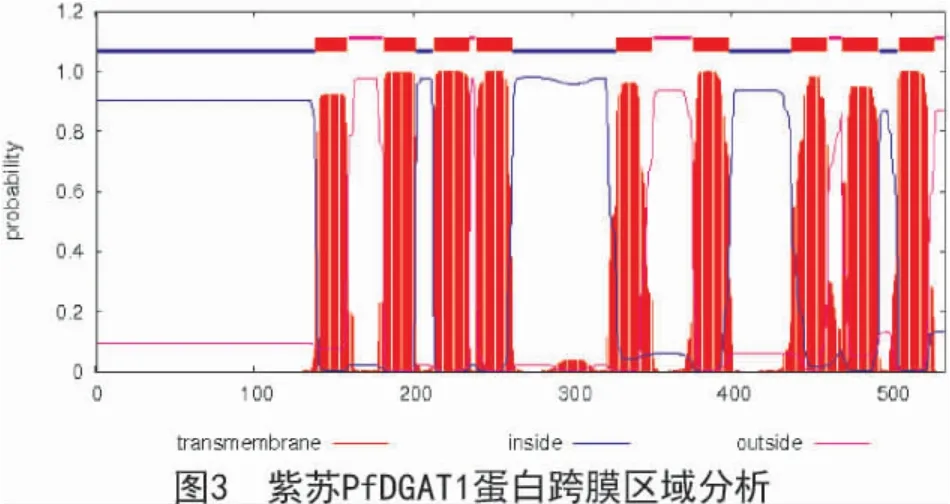
通過TMHMM 在線分析紫蘇PfDGAT1 蛋白跨膜區域,結果顯示(圖3),PfDGAT1 蛋白有 9 個典型的跨膜螺旋區,分別位于139~158,182~201,213~235,240~262,328~350,376~398,438~460,470~492,505~527 位氨基酸之間。通過NCBI-CDD數據庫分析發現,PfDGAT1 蛋白具有PLN02401(diacylglycerolo-acyltransferase)結構域,屬于MBOAT超家族中的成員。

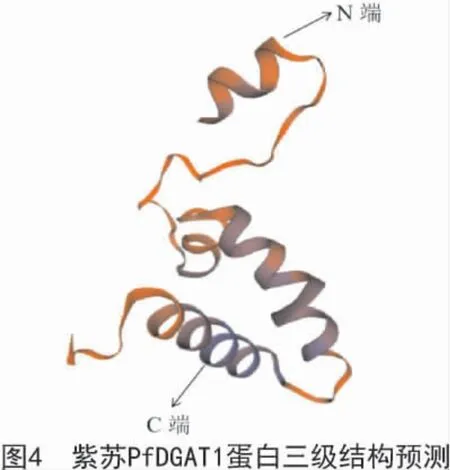
運用SOPMA 在線軟件對紫蘇PfDGAT1 蛋白的二級結構進行預測分析,結果顯示,該蛋白二級結構的主要結構元件為α-螺旋(43.63%)和無規則卷曲(41.76%),β-轉角(2.81%)和延伸鏈(11.80%)則分散于整個結構中。使用SWISS-MODEL 在線軟件對PfDGAT1 蛋白進行三維結構分析與建模,結果如圖4所示。
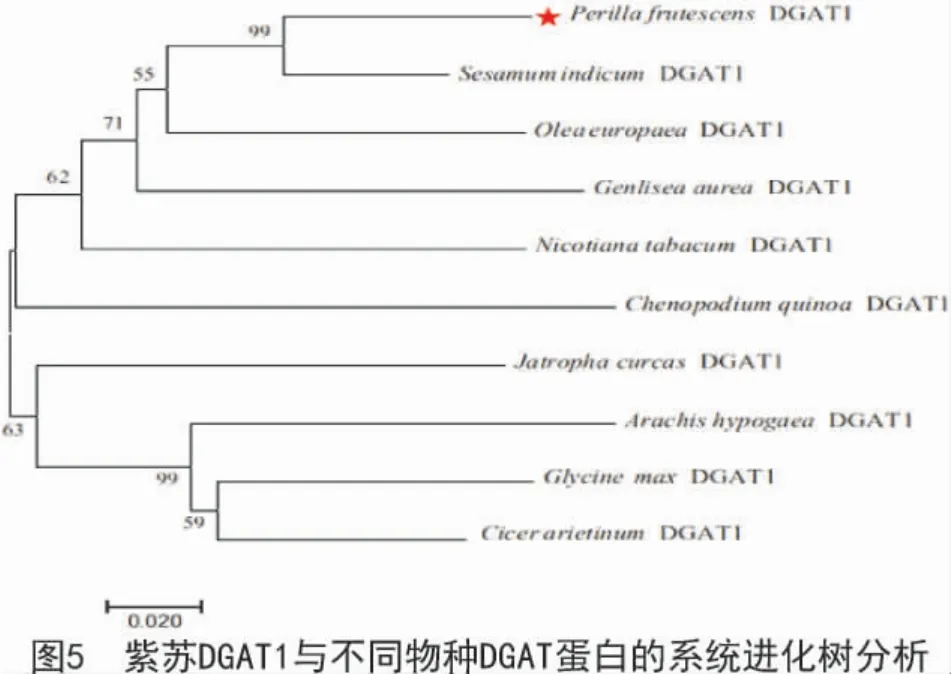
通過對紫蘇PfDGAT1 蛋白與其他植物DGAT1蛋白進行多序列比對發現,紫蘇PfDGAT1 蛋白與其他植物蛋白一樣,具有9 個典型的結構域,分別是酰基輔酶A 結合位點、擬南芥突變體AS11 串聯重復序列、催化活性位點、磷酸泛酰巰基乙胺附著位點、蛋白激酶1 靶標位點、硫酰基酶中間體結合位點、脂肪酸結合蛋白標簽、甘油二酯結合位點、內質網定位保守域。系統進化樹分析結果顯示(圖5),紫蘇PfDGAT1 蛋白與芝麻、油橄欖的親緣關系較近,且序列同源性分別為89%,83%,與大豆和鷹嘴豆的親緣關系較遠,表明DGAT1 蛋白在進化上具有高度保守性。
2.3 紫蘇PfDGAT1基因表達特性分析
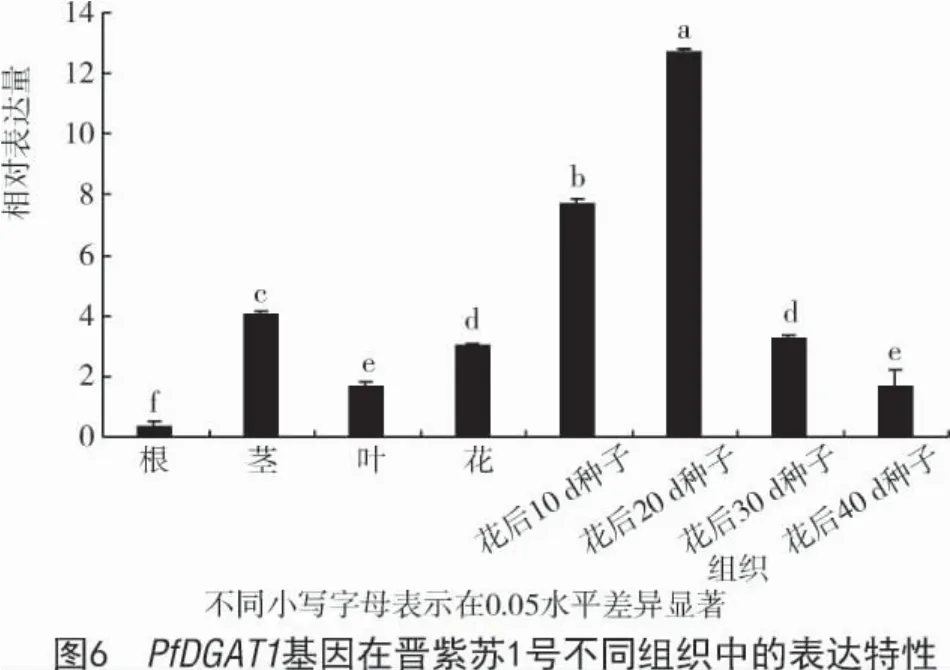
利用熒光定量PCR 技術分析紫蘇PfDGAT1 基因在晉紫蘇1 號根、莖、葉、花及不同發育時期種子中的表達特性,結果表明(圖6),PfDGAT1 基因在紫蘇不同組織中均有表達,在種子中表達量最高,在莖和花中的表達量次之,在根中基本不表達,且隨種子發育該基因表達量呈先升高后降低的變化趨勢,且在開花后20 d 達到最高。以根為對照,PfDGAT1 基因在莖、葉、花中的表達量分別是根中的 11.82 倍、1.77 倍和 8.57 倍,在開花后 20 d 的種子中該基因的表達量是根中表達量的36.29 倍。
3 結論與討論
二酰甘油酰基轉移酶(DGAT)是TAG 合成最后一步反應的關鍵限速酶,有研究表明,植物DGAT1 蛋白一般具有 9~10 個跨膜結構域[13,21]。本研究使用含油量較高的紫蘇品種晉紫蘇1 號作為試驗材料,克隆獲得PfDGAT1 基因后,分析該基因序列特征及編碼蛋白基本性質,結果發現,紫蘇PfDGAT1 蛋白具有9 個典型的跨膜螺旋區,這與前人研究結果一致[21]。多序列比對結果發現,PfDGAT1蛋白具有酰基輔酶A 結合位點、擬南芥突變體AS11 串聯重復序列、催化活性位點、磷酸泛酰巰基乙胺附著位點、蛋白激酶1 靶標位點、硫酰基酶中間體結合位點、脂肪酸結合蛋白標簽、甘油二酯結合位點、內質網定位保守域等9 個功能結構域,這與 HOBBS 等[11],ZOU 等[22],HE 等[23],BOUVIER 等[13],NYKIFORUK 等[24]對植物DGAT1 蛋白功能位點預測結果一致。
MURPHY 等[25]研究發現,DGAT 廣泛分布于植物的各個組織中,其活性在植物的花及發育的種子中最高。本試驗利用熒光定量PCR 分析PfDGAT1基因在紫蘇不同組織器官中的表達特性,結果顯示,PfDGAT1 基因在紫蘇不同組織中均有表達,其中,在種子中表達量最高,在莖和花中的表達量次之,在根中基本不表達,且隨種子發育,該基因表達量呈現先升高后降低的變化趨勢。王計平等[19]研究發現,在紫蘇種子發育過程中,其總脂肪酸含量逐漸升高,且PfDGAT1 基因表達量先升高后下降,與本研究結果基本一致,表明PfDGAT1 基因在紫蘇脂肪酸合成積累中發揮重要作用。
本研究克隆得到紫蘇PfDGAT1 基因cDNA,該序列全長 1 964 bp,其中,CDS 為 1 605 bp,共編碼534 個氨基酸,預測PfDGAT1 基因編碼定位于細胞質膜,具有典型的DGAT1 功能結構域,包含9 個典型的跨膜螺旋區。多序列比對和系統進化樹分析結果表明,PfDGAT1蛋白與芝麻、油橄欖的DGAT1 蛋白的親緣關系較近。qRT-PCR 結果表明,PfDGAT1基因在紫蘇根、莖、葉、花及種子發育不同階段均有表達,且在種子中表達量最高,PfDGAT1 基因表達量隨種子發育呈先升高后降低的變化趨勢。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
