轉cry1C基因抗蟲水稻吉生粳3號外源基因整合分析與品系特異性檢測
2019-05-17 09:36:42金永梅馬瑞于志晶林秀峰
生物技術通報 2019年3期
關鍵詞:水稻
金永梅 馬瑞 于志晶 林秀峰
(吉林省農業科學院農業生物技術研究所,長春 130124)
自1996年以來,全球轉基因農作物商業化種植面積和銷售收入均以倍數增長。水稻是我國乃至全世界最重要的糧食作物之一,目前,全球轉基因水稻研究進展迅速,我國轉基因水稻研發與世界同步,成功培育出抗螟蟲、抗除草劑、耐鹽、氮磷高效轉基因和優質等系列轉基因水稻品種/品系,為我國轉基因水稻的產業化提供了技術和品種儲備[1]。
通過轉基因技術可以將外源基因導入到受體中,改良目標農藝性狀。外源基因的插入改變了一個基因與其鄰近基因或其鄰近染色質的位置關系,隨著外源基因的整合區域的不同可發生位置效應,從而影響宿主植物的特定功能及表型。因此,研究T-DNA插入位點的旁側序列及其插入位點就顯得尤為重要[2]。利用農桿菌遺傳轉化方法導入的外源基因在宿主基因組的插入位點是隨機的,每個轉化事件中外源基因和基因組DNA拼接組成的旁側序列具有唯一性。因此,T-DNA旁側序列分析一直是轉基因植物研究的熱點,它對于闡述T-DNA整合方式、染色體定位、轉基因表達活性等具有重要意義。T-DNA整合過程是個復雜的異常重組過程,包括植物基因組整合位點的刪除和重復,T-DNA序列的刪除和重復,以及染色體上的易位和到位等現象[3-4]。
染色體步移(Chromosome walking)是指從生物基因組或基因組文庫中的已知序列出發,逐步探知其旁鄰的未知序列或與已知序列呈線性關系的目標序列的方法。分離旁側序列的染色體步移方法主要采用基于PCR擴增的染色體步移技術中的半隨機引物PCR策略。半隨機引物PCR策略中的熱不對稱交錯PCR(Tail-PCR)是將目標序列旁的已知序列中設計的3個嵌套的特定引物(Special primer SP1,SP2,SP3)分別與1個具有低Tm值的隨機簡并引物(Arbitary degenerate Primer,AD)相結合,根據引物的長短和特異性的差異設計不對稱的溫度循環,通過分級反應來擴增特異產物的方法,該方法準確性高、操作相對簡單[5]。
根據所分離的旁側序列,通過與NCBI的比對分析確定T-DNA在基因組中的整合位點。該序列是不同轉基因作物品系的特異性身份標識,根據其建立的品系特異性檢測方法能快速、準確地鑒定不同轉基因作物品系[6-7],保證了轉基因作物檢測的準確性和專一性,為監管轉基因作物的育種、生產、加工和銷售提供了可靠的技術支撐[8-9]。利用外源基因在受體基因組上整合位點的唯一性,可以建立品系特異性檢測方法,該方法可根據外源DNA與旁側植物基因組連接區序列為靶標設計引物,利用PCR方法擴增出對照和轉基因品系特異的PCR條帶[10]。
Bt抗蟲基因cry1C所編碼的殺蟲蛋白可以抑制和殺死鱗翅目害蟲,是在野生型Cry1Ca5基因的基
礎上通過序列優化人工合成的抗蟲基因[11]。目前,利用生物技術手段將cry1C基因導入到各種植物中,包括水稻[12-14]、大豆[15-16]、玉米[17-18]等作物,成功獲得了對抗鱗翅目害蟲具有抗性的轉基因植物。前期研究中,我們利用農桿菌介導的遺傳轉化方法,將cry1C基因導入到水稻品種吉粳88中,獲得了抗蟲轉基因水稻吉生粳3號[19]。本研究旨在通過染色體步移方法獲得吉生粳3號的左、右邊界旁側序列,依據旁側序列確定T-DNA在基因組中的插入位點,并建立吉生粳3號品系特異性PCR方法,為吉生粳3號的身份識別提供理論依據。
1 材料與方法
1.1 材料
轉基因水稻吉生粳3號由本課題組培育保存;非轉基因水稻吉粳88由吉林省農業科學院水稻研究所提供。
1.2 方法
1.2.1 基因組DNA的提取 將種子播種在1/2MS培養基中,7 d后取樣保存。采用CTAB方法提取0.1g水稻樣品中的基因組DNA[20]。DNA樣品的純度和濃度用NanoDrop進行檢測。
1.2.2 轉基因植株吉生粳3號的Southern Blot 利用CTAB法提取水稻葉片的基因組DNA,利用DIGHigh Prime DNA Labeling and Detection Starter Kit I(Roche公司)進行Southern雜交,方法參考試劑盒說明書。具體步驟為:取60 μg基因組DNA,用限制性內切酶SacI(TaKaRa)進行完全酶切。酶切產物用0.8% 瓊脂糖凝膠電泳分離后轉膜(Amersham),用地高辛標記的cry1C探針進行雜交,再用X-光膠片化學發光自顯影。
擴增探針引物序列為:cry1C-F:5'-TTCTACTGGGGAGGACATCG-3',cry1C-R :5'-CGGTATCTTGGGTGATTGG-3'。
1.2.3 轉基因水稻吉生粳3號的右邊界旁側序列的獲取 采用染色體步移試劑盒(TaKaRa GenomeWalking Kit,Code No.6108)分離吉生粳3號中外源載體插入位點處的旁側序列。在已知的T-DNA序列右邊界設計3條同向且退火溫度較高的特異性引物(表 1)。
以1 μL(200 ng 左右)吉生粳3號基因組DNA為模板,使用右邊界特異性引物和試劑盒中的簡并引物AP1、AP2、AP3、AP4進行巢式PCR。

表1 T-DNA右邊界巢式引物序列
巢式PCR擴增體系為:1st PCR 反應在50 μL體系中進行,以1 μL基因組DNA,0.5 μL TaKaRa LA Taq(5 U/μL),5 μL 的 10×LA PCR Buffer II(Mg2+plus),8 μL 的 dNTP Mixture(2.5 mM each),1 μL的 AP1、AP2、AP3、AP4 引 物(100 pmol/μL),1 μL的SP1引物(10 pmol/μL)進行擴增;2nd和3rd PCR反應分別將上一輪PCR反應液稀釋1 000倍后,取 1 μL作為 PCR 反 應 模 板,1 μL的 AP1、AP2、AP3、AP4 引物(100 pmol/μL)分別與 1 μL 的 SP2和SP3引物(10 pmol/μL)進行擴增PCR擴增體系同上;巢式PCR反應擴增程序按照染色體步移試劑盒(TaKaRa GenomeWalking Kit,Code No.6108) 中的方法進行。
取1st,2nd,3rd PCR反應液各5 μL,使用1%的瓊脂糖凝膠進行電泳,分析PCR擴增條帶并篩選有義引物組合,切膠回收被選條帶,利用pMDTM 19-T Vector(Takara,Code No.6013)進行 TA 克隆(TaKaRa DNA Ligation Kit,TaKaRa,Code No.6022),陽性質粒進行測序獲得右邊界旁側序列,測序引物為5'-CCAGTTCAGCTAATCTTCTT-3'。引物合成及PCR擴增條帶的測序均由寶生物有限公司完成。
1.2.4 旁側序列比對分析及插入位點的確定 利用NCBI BLAST 軟件(http://www.ncbi.nlm.nih.gov/BLAST),將PCR擴增條帶的測序結果分別與水稻基因組序列和T-DNA序列進行同源比對分析,確定外源T-DNA在吉生粳3號基因組中的插入位點。
1.2.5 吉生粳3號的左邊界旁側序列獲取 在吉生粳3號基因組中T-DNA插入位點的左側設計正向引物(C10-1496F),在T-DNA左邊界設計3條同向巢式引物(C10F-SP4,C10F-SP5,C10F-SP6),進行PCR擴增以進一步確認插入位點的正確性。引物序列見表2。

表2 T-DNA左邊界巢式引物序列
PCR 擴增體系為:總體系20 μL,康為世紀2×Es Taq MasterMix 10 μL(含有 Es Taq DNA Polymerase,PCR buffer,3 mmol/L MgCl2,400 μmol/L dNTP mix),引物各 1 μL(10 μmol/L),水稻 DNA 2 μL(100 ng/μL),剩余用ddH2O 補足;PCR 擴增程序為,94℃ 1 min;94℃ 10 s,55℃ 1 min,72℃ 40 s,35 個循環 ;72℃ 10 min。PCR產物經過回收、純化、測序獲得左邊界旁側序列。
1.2.6 吉生粳3號特異性PCR檢測 提取非轉基因對照品種(吉粳88)和吉生粳3號基因組DNA進行特異性PCR檢測。PCR反應在25 μL體系中進行 :12.5 μL 的 康 為 世 紀 2×Es Taq MasterMix( 含有 Es Taq DNA Polymerase,PCR buffer,3 mmol/L MgCl2,400 μmol/L dNTP mix),1 μL 10 μmol/L 正向引物,1 μL 10 μmol/L 反向引物,1 μL T-DNA 特異性引物,水稻基因組 DNA 2 μL(100 ng/μL),剩余用ddH2O 補足。反應條件為:94℃ PCR預變性2 min,94℃變性 30 s,55℃退火 30 s,35循環,72℃10 min,引物序列如下:正向引物為C10-F:TGCACGCCTACTCCACATTA,反向引物為C10-R:TGCTTGCTAGACTCACCACA,T-DNA特異性引物為C10-TDNAR:TCACCACTCGATACAGGCAG。
2 結果
2.1 轉基因水稻吉生粳3號右邊界旁側序列及插入位點
通過農桿菌遺傳轉化法,將Bt抗蟲基因cry1C的P3300-cry1C植物表達載體,導入到吉林省水稻品種吉粳88中,獲得了抗蟲轉基因水稻獨立轉化植株。通過分子檢測和抗性分離比分析,從49個獨立轉化植株中篩選出8個單拷貝株系。分別提取和非轉基因水稻和吉生粳3號的基因組DNA,用限制性內切酶SacI和DraI分別進行酶切,地高辛標記的cry1C探針進行雜交,篩選出外源目的基因cry1C為單拷貝的轉基因水稻株系吉生粳3號(圖1)。
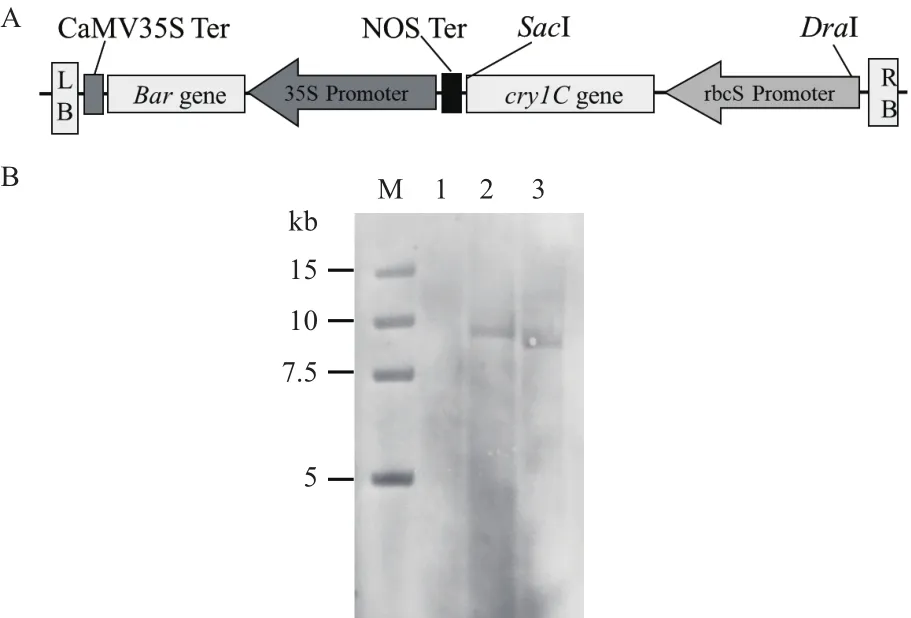
圖1 植物表達載體示意圖與Southern blot
為了獲取吉生粳3號右邊界旁側序列,提取基因組DNA,利用染色體步移方法獲得PCR特征條帶。T-DNA序列右邊界3條同向巢式引物C10R-SP1、C10R-SP2、C10R-SP3,分別與簡并引物AP1、AP2、AP3、AP4進行3輪PCR擴增。結果(圖2)表明,3輪PCR所有引物組合均獲得比較清晰的PCR條帶,但只有簡并引物AP2與3條巢式引物組合所獲得的條帶大小呈遞減趨勢(泳道4-6),因此可以判斷為吉生粳3號右邊界旁側序列的特異性條帶。
切膠回收C10R-SP3與AP2引物組合擴增出的PCR條帶(圖2-B中泳道6,約1.8 kb),通過TA克隆方法獲得包含目的片段的載體,經過目的片段的測序最終獲得目的片段的核苷酸序列。
將吉生粳3號右邊界旁側序列(圖3-A),利用NCBI數據庫中的 BLAST工具進行比對,確定了T-DNA在吉生粳3號基因組中的插入位點。比對分析結果表明,吉生粳3號右邊界旁側序列中1-67 bp區段與載體序列右邊界的部分序列有100%的相似性,68-732 bp區段與水稻基因組(日本晴)2號染色體上的基因間區(非編碼區)2 790 589-2 789 925 bp區段具有99%的相似性,因此插入位點初步可以判斷為2 790 589(圖3-B)。
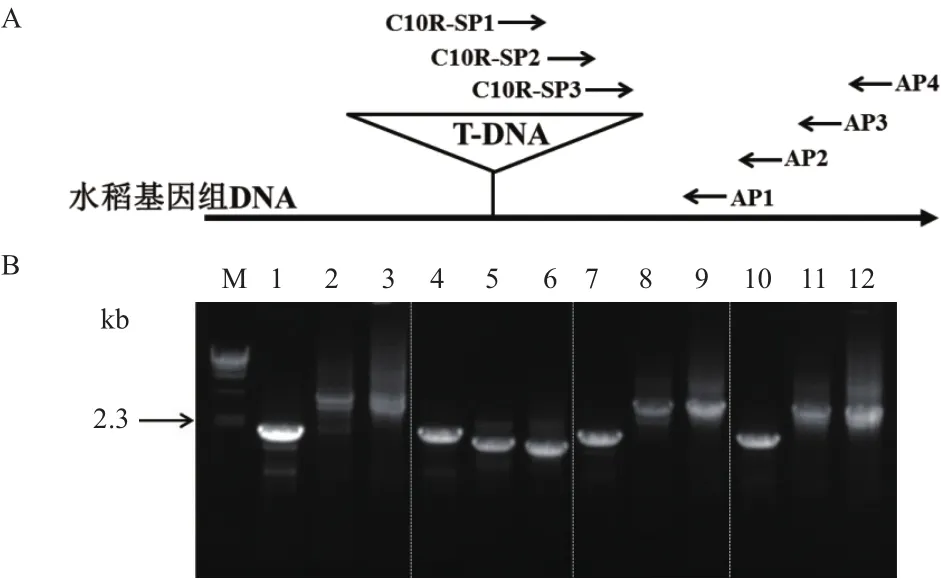
圖2 吉生粳3號右邊界旁側序列巢式PCR
2.2 吉生粳3號左邊界旁側序列及插入位點
根據吉生粳3號右邊界旁側序列確定的插入位點,在吉生粳3號插入位點左側基因組中設計正向引物(C10-Chr2-1496F),在T-DNA左邊界區段設計3條同向T-DNA特異性巢式引物(C10F-SP4,C10F-SP5,C10F-SP6)(表 2)進行 PCR 擴增,進一步確認插入位點的正確性。PCR結果獲得了3條大小遞減的PCR特征條帶,對其中一條PCR條帶(泳道3)進行了回收、克隆、測序(圖4)。測序獲得的序列分別與載體序列和水稻基因組序進行列比對確定插入位點,結果插入位點確定為水稻基因組(日本晴)2號染色體上的2790685。
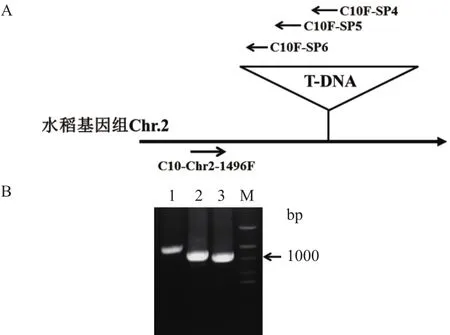
圖4 吉生粳3號左邊界PCR
2.3 吉生粳3號的T-DNA插入位點及品系特異性PCR檢測
根據右邊界旁側序列可以判斷T-DNA在吉生粳3號基因組中的插入位點為2號染色體上的2 790 589位,根據左邊界旁側序列可以插入位點為2號染色體上的2 790 685位。因此,根據左、右邊界旁側序列可以確定T-DNA的插入位點為2號染色體上的2 790 685-2 790 589位點,水稻基因組序列發生了96個堿基的缺失(圖5-A)。
在吉生粳3號基因組中T-DNA的插入位點兩側分別設計正向引物和反向引物(C10-F和C10-R),在T-DNA的左邊界設計特異性引物(C10F-TDNAR),進行吉生粳3號特異性PCR檢測。提取非轉基因對照、吉生粳3號、和其它兩個轉基因水稻株系的基因組DNA,用上述3條引物同時進行PCR擴增。在PCR擴增反應程序中延伸時間限定為40 s,因此在吉生粳3號中引物C10-F和C10F-TDNAR可進行擴增,在非轉基因對照和其它基因轉基因水稻株系中引物C10-F和C10-R可進行擴增。PCR結果(圖5-B)發現,轉基因水稻吉生粳3號中擴增出一條428 bp的轉基因特異性片段,非轉基因對照和其它兩個轉基因株系中擴增出一條1 023 bp的非轉基因特異性片段,與預期大小一致,說明引物C10-F、C10-R、C10F-TDNAR能夠特異性地識別吉生粳3號水稻。
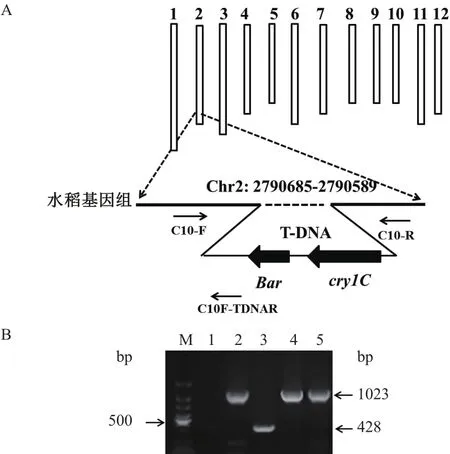
圖5 T-DNA在吉生粳3號基因組中的插入位點及品系特異性PCR
3 討論
在轉基因生物中外源基因的T-DNA旁側序列具有非常重要的意義,利用它可以分析T-DNA在宿主生物基因組中的整合方式、染色體定位、對內源基因的表達影響等情況。目前,分離旁側序列的方法有Tail-PCR、反向PCR、質粒拯救、染色體步移、全基因組測序等。李亞麗等[21]通過Tail-PCR方法分離了轉mCherry基因水稻T-DNA旁側序列并確定了它在基因組中的整合位點;Cao等[22]利用染色體步移方法分離轉基因小麥品系B10201-2的旁側序列,并建立了品系特異性PCR方法;Guo等[23]利用全基因組測序方法,在轉基因大豆中分離了G2EPSPS基因和GAT基因的旁側序列并確定了基因組中的插入位點。
轉基因植物中外源基因的整合區域與外源基因的穩定表達及宿主基因組中上下游基因的正常表達密切相關。如果T-DNA整合進入內含子,外源基因容易造成基因沉默,導致目的基因不表達;如果T-DNA整合進入功能基因內部,會影響功能基因的正常表達并會對基因功能造成影響。因此,轉化事件篩選過程中的眾多指標中選擇T-DNA整合進入基因間區的轉化事件是重要指標之一。有研究表明,外源基因片段易整合進入植物基因組中轉錄活躍區域或染色體的末端[24-25];在轉基因擬南芥中T-DNA整合位點與染色體上基因分布的密度相關,T-DNA插入位點傾向于基因間區域,整合在外顯子與內含子區域的頻率沒有顯著性差異[26]。為了分析外源基因T-DNA在抗蟲轉基因水稻吉生粳3號中的整合區域與插入位點,我們通過染色體步移方法分離了吉生粳3號的右邊界旁側序列。根據右邊界旁側定位外源基因在轉基因水稻吉生粳3號基因組中的插入位點并分離出右邊界旁側序列。由于吉生粳3號轉基因水稻的外源基因T-DNA區域插入到水稻基因組2號染色體的基因間區,因此,外源片段的插入不會使吉生粳3號產生位置效應。通過對轉基因水稻吉生粳3號多個世代的外源目的基因遺傳穩定性分析表明,cry1C基因在DNA水平、RNA水平和蛋白水平上表達穩定;農藝性狀和目標性狀鑒定分析發現,除了抗蟲性之外其他農藝性狀與對照品種沒有顯著差異,并且保持穩定,說明T-DNA在吉生粳3號基因組中的插入位點對外源基因自身的穩定表達和水稻內源基因的表達均沒有造成不利影響(未發表數據)。
吉生粳3號的左右邊界旁側序列分析表明,在外源基因的整合過程中T-DNA右邊界旁側序列的斷裂點在A、T堿基處,左邊界斷裂點在C、T堿基處,水稻基因組序列缺失了96 bp,載體序列未發生堿基缺失或易位等變化。有研究表明轉基因水稻T-DNA左右邊界及邊界內側序列的斷裂點主要集中在A、T堿基處[20];Tzfira等[27]認為T-DNA 整合進宿主基因組的主要路徑為雙鏈斷裂修復機制,T-DNA整合過程中農桿菌中的稀有位點限制性內切酶在宿主中表達,使宿主基因組DNA產生雙鏈斷裂,并且在宿主體內消化T-DNA,導致宿主基因組序列的缺失和T-DNA的丟失。
轉基因作物品系特異性檢測方法特異性高,適合轉基因產品的定性和定量檢測,是對轉基因植物進行監督管理、保障其健康發展的重要技術基礎。郭超等[28]利用hiTail-PCR方法獲得了轉基因水稻BarKasalath-01中外源基因在基因組上的旁側序列并建立了該轉化事件特異性檢測方法;金蕪軍[29]等人于2013年基于轉基因抗蟲水稻品系TT51-1的旁側序列建立了鑒定純合型轉基因抗蟲水稻品系TT51-1 的方法。本研究中,根據外源基因T-DNA在吉生粳3號基因組中的整合位點,在整合位點兩側和T-DNA左邊界共設計3條引物,建立了吉生粳3號品系特異性PCR檢測方法。本研究所建立的cry1C轉基因吉生粳3號品系水稻特異性檢測方法,對提高該品系的鑒定效率及準確性具有重要意義。
4 結論
本研究利用染色體步移方法,獲得轉cry1C基因抗蟲水稻吉生粳3號的外源基因旁側序列及其水稻基因組中的插入位點,并建立了吉生粳3號品系特異性PCR檢測方法。該結果為吉生粳3號的身份識別提供了準確、快速的檢測技術手段。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
