杉木人工林下原生與引入樹種葉性狀變異特征
2019-05-13 01:49:34邢文黎虞木奎成向榮
生態學報 2019年6期
關鍵詞:植物
熊 靜,邢文黎,虞木奎,成向榮,*
1 中國林業科學研究院亞熱帶林業研究所,杭州 311400 2 南京林業大學,南京 210037
人工林在全球森林資源中扮演著重要角色,是緩解采伐天然林資源提供木材供給的有效補充,但普遍存在結構不盡合理、生產力不高、生態功能較弱和生態穩定性下降的問題[1]。目前,人工林的可持續經營理論與多目標經營的優化模式仍在不斷實踐探索中。杉木(Cunninghamialanceolata(Lamb.) Hook.) 是亞熱帶地區主要造林樹種之一,其造林面積占世界人工林面積的6.5%,占我國人工林面積的19%,人工林蓄積量的25%,在我國林業生產中具有重要的地位[2]。隨著杉木人工林集約經營發展,杉木連栽導致的土壤質量退化、林分生產力降低等問題日益突出[3- 4]。改變人工純林單一林分結構,構建復層異齡林對改善林地土壤質量,提高林地生產力發揮了重要作用[5- 6]。
光是影響森林中林下植物生長和分布最主要的環境因素[7],是構建異齡復層林最主要的限制因子之一。植物對林下弱光環境的適應能力大小對其生長發育和生產力高低至關重要。葉片是植物物質生產、能量轉換及交換的主要器官,其性狀特征直接影響群落的結構和功能[8]。葉性狀是植物的重要特性之一,屬于植物功能性狀的二級性狀,具有相對的穩定性[9]。葉性狀分為功能型性狀和結構型性狀,包括葉的形態、養分和滲透調節物質等[10],易測定且可同時對大量植物種類進行比較[11]。葉性狀可直接影響植物的基本行為與功能[12- 13],能反映植物在表征生態系統功能方面的生態指示作用,強調其與生態系統過程及功能的關系[14]。在眾多植物性狀中,植物的一些葉片性狀與植物的生存對策及植物利用資源的能力緊密聯系,通常為適應環境變化而形成一系列生存對策[11],同時植物葉功能性狀之間亦存在相關性,這反映出植物對環境的趨同適應特征[8]。隨著全球環境及氣候的變化,應用葉性狀因子(如形態、解剖結構、化學計量等)研究植物在不同生境中的適應機制成為生理生態學領域近年來研究中的熱點[15- 17]。目前對葉性狀的研究主要集中在干擾和脅迫[12,18]、不同地區或流域特殊環境下植物葉性狀間關系[19- 22]或集中在某一物種或生活型水平上葉性狀的研究[12,22- 24],但葉性狀在人工林林分結構改造上的探究較少。研究小區域小尺度不同生活型植物在形態、生理及光合作用方面的葉功能性狀的差異性表達,有利于探究植物的適應機制,同時也有利于篩選與該地區植物生長及適應機制緊密聯系的關鍵性狀。本研究通過同質園試驗在浙江省開化縣林場杉木純林下套種耐陰性伴生喬木和灌木樹種,探討其與林下生境過濾后自然更新的原生喬木與灌木樹種之間葉性狀差異性及相關性,以期為亞熱帶地區杉木復層異齡林伴生樹種的篩選及其對林下環境適應機制解析提供一定理論參考與科學依據。
1 材料與方法
1.1 試驗區概況
研究區位于浙江省開化縣林場城關分場小橋頭林區(118°25′E,29°09′N)(圖1)。該區年日照總時數1334.1 h,年平均氣溫16.4℃,年平均降雨量1814 mm,無霜期252 d,氣候屬亞熱帶季風氣候。土壤類型為紅黃壤,pH為4.0—4.9。供試杉木純林為1996年春季造林,初植密度為2500株/hm2,2006年進行1次撫育間伐,目前保留密度為1358株/hm2,平均樹高和胸徑分別為16.7 m和21.6 cm,林分郁閉度約0.85。
1.2 試驗方法
2011年3月,在小橋頭林區杉木人工林下引入13種耐陰樹種(無外來種)構建杉木復層異齡林,樹種詳細信息見表1。灌木樹種沿等高線栽植,密度3000株/hm2;喬木樹種沿杉木行間栽植,密度2500株/hm2,具體樹種配置方式見圖1。造林苗木均采用1年生健壯容器苗,每個樹種塊狀隨機栽植于杉木林下,面積0.2—0.5 hm2。為比較引入樹種與原生樹種之間葉性狀的關系,同時選擇杉木林下自然生長的優勢喬木與灌木(表1)。根據研究需要,將所有樹種分為原生樹種與引入樹種;原生樹種又分為原生喬木與原生灌木;引入樹種分為引入喬木與引入灌木。
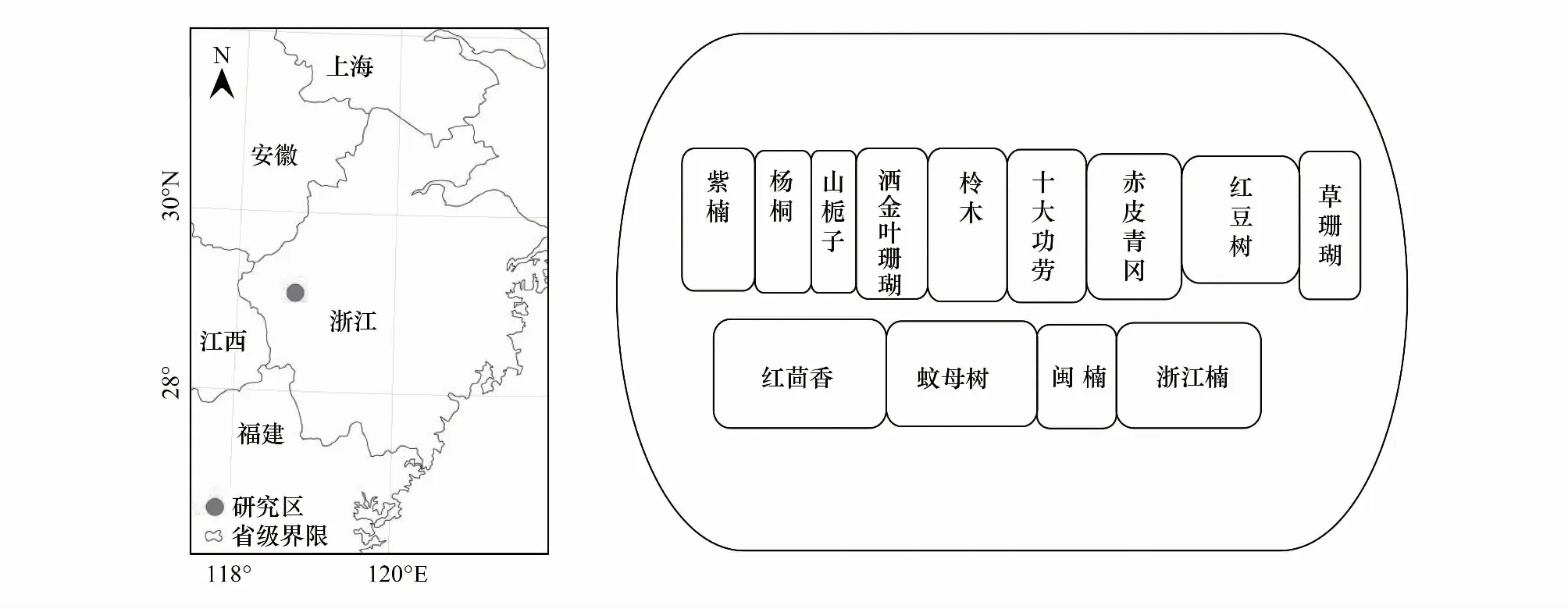
圖1 試驗區地理位置及引入樹種栽植配置圖Fig.1 Geographical location of the test area and planting configuration map of introduced tree species
1.3 葉性狀測定
2017年8月在杉木人工林下分別選取引入和原生樹種的當年生中上部小枝上的健康成熟葉片150—200片,避開林窗與林緣,采集的葉片編號后及時放入裝有冰袋的保溫箱內。將采集的葉片帶回實驗室后,一部分用于葉片形態指標分析,如葉長(Leaf length,LL) 、葉寬(Leaf width,LW)、葉長/葉寬(LL /LW)、葉面積(Leaf area,LA)等指標用LI-8400plus葉面積儀測定后采用WinFOLIA分析系統確定,掃描后的葉片烘干至恒重測葉干重(Leaf dry weight,LDW)并計算比葉面積(Specific leaf area,SLA)=掃描葉面積/掃描葉干重。剩余葉片用于生理和養分含量測定,葉綠素a(Chlorophyll a,Chla)、葉綠素b(Chlorophyll b,Chlb)含量采用乙醇-丙酮比色法測定;葉片淀粉(Starch,St)含量采用高氯酸水解-蒽酮比色法測量;可溶性糖(Soluble sugar,SS)采用蒽酮比色法量;非結構性碳水化合物(Non-structural carbohydrate,NSC)為淀粉與可溶性糖含量總和;葉碳(C)含量采用TOC分析儀測定;葉氮(N)含量用濃硫酸-過氧化氫消煮-半微量凱氏定氮法測定;葉片磷(P)含量采用HClO4-H2SO4消煮-鉬銻抗比色法測定[25]。
1.4 數據處理
采用Excel 2013和SPSS 22進行數據分析和處理。以SPSS 22統計分析軟件對實驗數據進行方差分析、并對不同功能群進行Duncan多重比較檢驗,對葉性狀的相關性進行Pearson相關分析,不同功能群葉性狀的線性相關圖采用對數函數轉換后的數據進行繪制。以Canoco 4.5對葉性狀數據進行主成分分析;表中數據為平均值(Mean)±標準差(Standard of deviation,SD);變異系數(Coefficient of variation,CV)=(標準差/平均值)×100%。研究區位置圖采用ArcGIS 10.2繪制。

表1 引入樹種與原生樹種植物名錄及功能群分類
2 結果與分析
2.1 原生樹種與引入樹種葉性狀特征
根據原生樹種與引入樹種葉性狀分析(表2)可知:總體上引入樹種各葉形態參數數值略大于原生樹種,其中引入與原生樹種的葉寬、葉長寬比、葉面積與葉干重無顯著差異(表2)。引入樹種的葉長比原生喬木大32%(P<0.05),而與原生灌木無顯著差異。引入樹種比葉面積顯著高于原生喬木(P<0.05),而與原生灌木無顯著差異。無論是引入樹種或原生樹種,其葉面積、葉干重及比葉面積均表現出較高的變異性,其中原生喬木總體葉片形態指標變異系數普遍較低。
引入樹種的總葉綠素含量顯著高于原生樹種(P<0.05)。引入喬木的總葉綠素含量顯著高于引入灌木(P<0.05),原生灌木的總葉綠素含量顯著高于原生喬木(P<0.05),引入喬木與原生灌木的總葉綠素含量無顯著差異。引入喬木的葉綠素a含量顯著高于其他樹種(P<0.05),原生灌木的葉綠素b含量顯著高于其他樹種(P<0.05);引入樹種與原生灌木的葉綠素a及葉綠素b含量的變異系數高于原生喬木。引入樹種的葉綠素a/b顯著大于原生植物(P<0.05),引入喬木和引入灌木的葉綠素a/b比值接近,分別比原生喬木和原生灌木高60%和45%。
引入樹種葉碳含量顯著高于原生樹種(P<0.05),尤其是引入喬木葉碳含量最高。原生灌木葉氮含量最高,其次為引入喬木,原生喬木和引入灌木的葉氮含量沒有顯著差異。引入喬木和灌木樹種與原生灌木的葉磷含量沒有顯著差異,但引入喬木和原生灌木葉磷含量顯著高于原生喬木(P<0.05)。引入樹種與原生樹種的葉氮、磷含量的變異系數高于葉碳含量。引入樹種與原生喬木的葉碳/氮均顯著大于原生灌木(P<0.05),引入樹種與原生灌木的葉碳/磷顯著小于原生喬木(P<0.05),引入喬木與原生樹種的葉氮/磷顯著大于引入灌木(P<0.05)。
灌木樹種的非結構性碳水化合物總量顯著大于喬木樹種(P<0.05),引入灌木顯著小于原生灌木(P<0.05)。引入樹種與原生喬木之間的淀粉含量差異不顯著,但均顯著小于原生灌木(P<0.05)。引入喬木與原生樹種間的可溶性糖含量無顯著差異,但均顯著小于引入灌木(P<0.05)。引入樹種的可溶性糖與淀粉含量的比值顯著高于原生樹種(P<0.05),其中引入灌木的可溶性糖與淀粉含量的比值取得最大值。引入樹種與原生樹種的可溶性糖及淀粉均表現出較高的變異性。
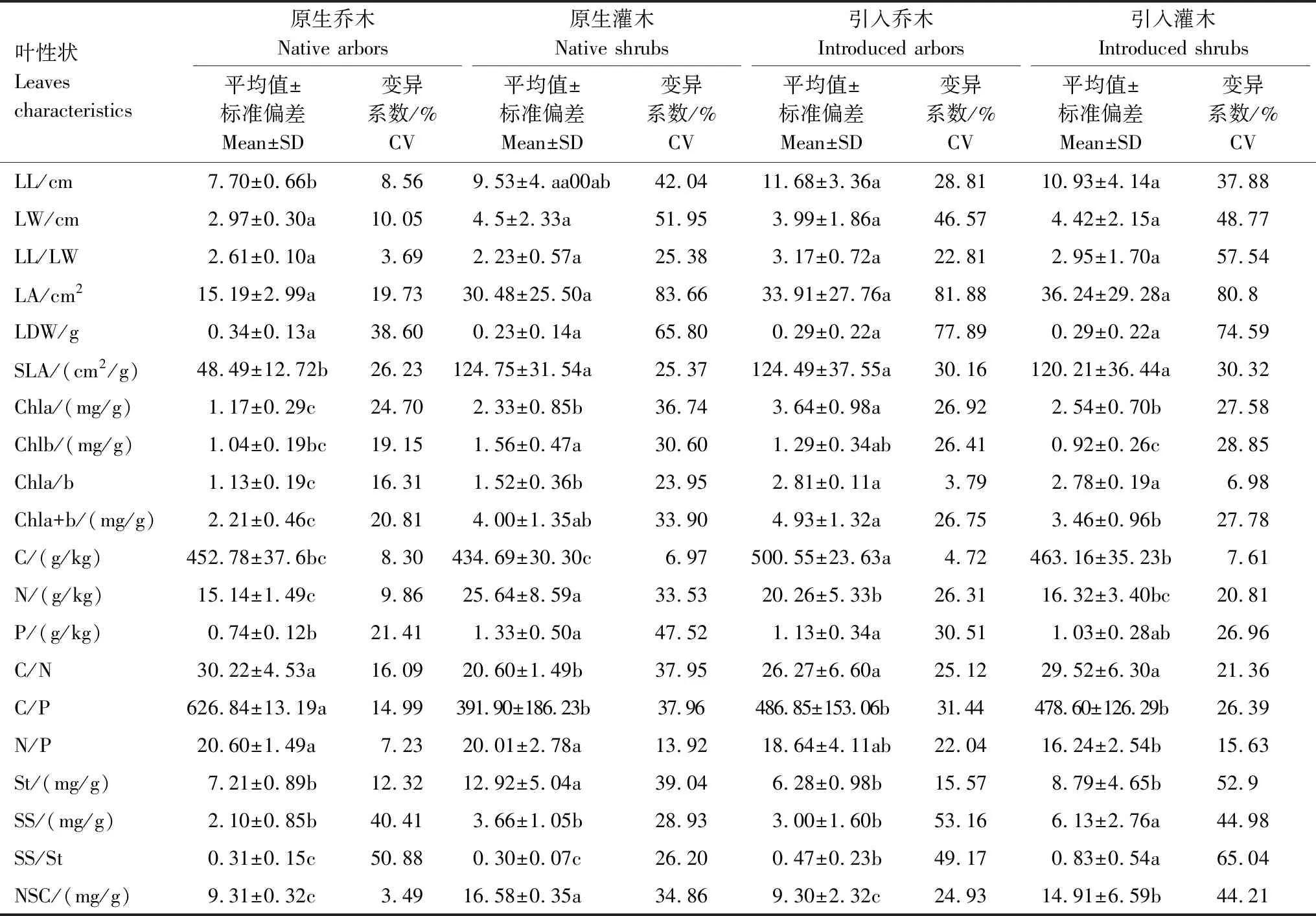
表2 原生樹種與引入樹種葉性狀特征
不同小寫字母表示同一葉性狀在不同功能群之間具有顯著差異(P<0.05);SD:標準偏差,Standard of deviation;CV:變異系數,Coefficient of variation;LL:葉長,Leaf length;LW:葉寬,Leaf width;LL/LW:葉長/葉寬,Leaf length/Leaf width;LA:葉面積,Leaf area;LDW:葉干重,Leaf dry weight;SLA:比葉面積,Specific leaf area;Chla:葉綠素a,Chlorophyll a;Chlb:葉綠素b,Chlorophyll b;Chla/b:葉綠素a/b,Chlorophyll a/b;Chla+b:葉綠素a+b,Chlorophyll a+b;C:碳,Carbon;N:氮,Nitrogen;P:磷,Phosphorus;C/N:碳/氮,Carbon/Nitrogen;C/P:碳/磷,Carbon/Phosphorus;N/P:氮/磷,Nitrogen/Phosphorus;St:淀粉,Starch;SS:可溶性糖,Soluble sugar;SS/St:可溶性糖/淀粉,Soluble sugar/Starch;NSC:非結構性碳水化合物,Non-structural carbohydrate
2.2 原生樹種與引入樹種的葉性狀相關性
引入樹種與原生樹種的葉性狀具有顯著的正相關關系(R2=0.2354,P=0.032,圖2),其中原生喬木與原生灌木(R2=0.96,P<0.001,圖2)、引入喬木與引入灌木(R2=0.9789,P<0.001,圖2)的葉性狀之間呈高度線性正相關,表明原生樹種之間、或引入樹種之間葉性狀的相關度均較強。引入喬木與原生喬木(圖2)、引入喬木與原生灌木(圖2)、引入灌木與原生喬木(圖2)、引入灌木與原生灌木(圖2)的葉性狀之間也呈現良好的正相關關系(P<0.05),表明在杉木人工林下喬木與灌木的葉性狀特征變化趨勢類似。
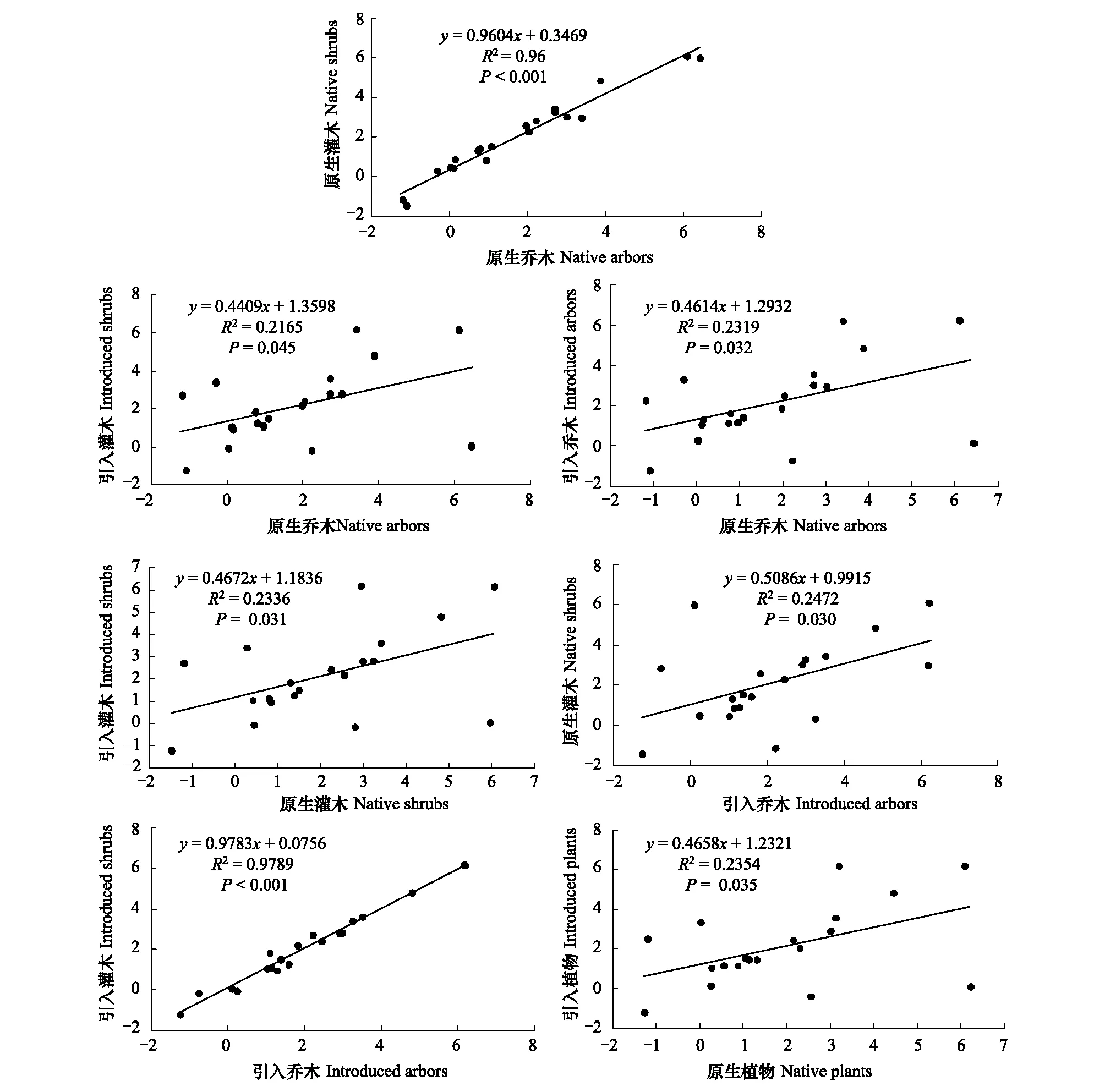
圖2 不同功能群間葉性狀的線性關系Fig.2 Linear relationship of leaf traits among different functional groups
從表3可以看出,總體上引入喬木與原生喬木的葉綠素含量與葉長寬比及葉干重無顯著相關性,而與葉長、葉寬、葉面積及比葉面積呈顯著正相關(P<0.05)。葉片養分含量中,葉碳、氮、磷含量與比葉面積呈極顯著正相關(P<0.01),與色素含量呈顯著正相關(P<0.05),葉碳/氮、葉碳/磷、葉氮/磷與比葉面積、葉綠素含量呈極顯著負相關(P<0.01)。非結構性碳水化合物中可溶性糖含量與其他葉性狀無顯著相關性,葉片淀粉含量、總非結構性碳水化合物含量與色素含量、葉氮含量和葉磷含量呈極顯著負相關(P<0.01),與比葉面積呈顯著負相關(P<0.05)。
引入灌木與原生灌木間Pearson相關性分析顯示:總體上葉綠素含量與葉形態性狀呈顯著正相關(P<0.05),而與葉干重無顯著相關性。葉片養分含量中,葉碳含量與葉長寬比呈極顯著負相關(P<0.01),而與其他形態性狀及色素含量無顯著相關性;葉氮含量和葉磷含量與葉寬、比葉面積及色素含量呈極顯著正相關(P<0.01);總體上葉碳/氮、葉碳/磷、葉氮/磷與葉形態性狀及葉綠素含量呈顯著負相關(P<0.05)。非結構性碳水化合物與比葉面積、葉綠素a+b含量無顯著相關性;淀粉與葉形態性狀及葉綠素a含量呈負相關(P<0.05);可溶性糖含量與葉綠素b含量呈顯著負相關(P<0.05);非結構性碳水化合物總量與葉綠素b含量、葉氮含量、葉氮/磷呈顯著正相關,與葉綠素a/b、葉碳含量、葉碳/氮呈顯著負相關(P<0.05)。
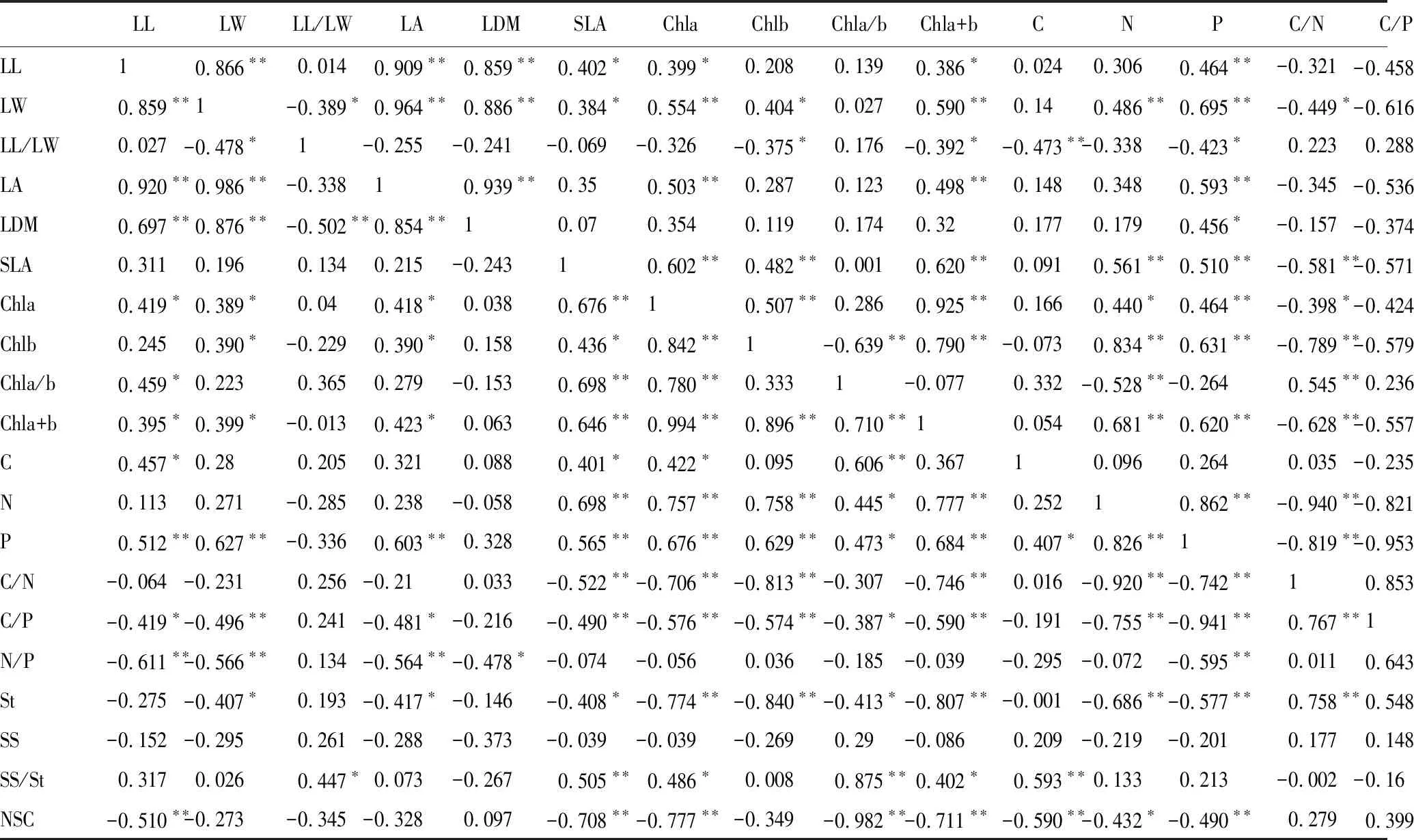
表3 喬木之間及灌木之間葉性狀Pearson相關性
表左下角為原生喬木與引入喬木間Pearson相關性,右上角為原生灌木與引入灌木間Pearson相關性。表中*表示在0.05水平上達到顯著水平(P<0.05),**表示在0.01水平上達到極顯著水平(P<0.01)
基于葉性狀和功能群的主成分分析(PCA)表明,前2個主成分可以解釋葉性狀變異總方差的87.9%,其中PC1可以解釋總方差的48.8%,PC2可以解釋總方差的39.1%(圖3)。葉長、葉寬、葉面積、比葉面積、葉綠素a、葉綠素a+b、可溶性糖等對PC1軸的貢獻率較大;葉碳/氮、葉長寬比、葉碳含量、葉綠素b、淀粉、總非結構性碳水化合物總量等對PC2軸的貢獻率較大。引入喬木(IA)與引入灌木(IS)樹種在葉碳含量、葉綠素a/b、可溶性糖、葉綠素a、葉面積等與其他功能群植物差異較大。原生喬木(NA)在葉碳/磷、葉碳/氮、葉干重等與其他功能群植物差異較大。原生灌木(NS)在淀粉、葉綠素b、葉氮、葉磷含量等與其他功能群植物差異較大。引入喬木與引入灌木的葉性狀較為接近;引入樹種與原生灌木的葉性狀差異相對較小,而與原生喬木差異較大;原生喬木與原生灌木樹種的葉性狀差異相對較大。
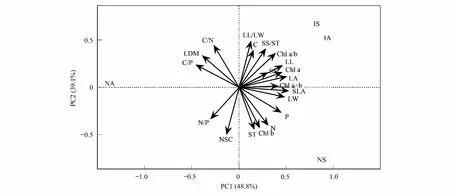
圖3 原生植物與引入植物葉性狀的主成分分析Fig.3 Principal component analysis (PCA) of traits of native and introduced plantsNA:原生喬木,Native arbors;NS:原生灌木,Native shrubs;IA:引入喬木,Introduced arbors;IS:引入灌木,Introduced shrubs
3 討論
植物葉性狀受環境的影響很大[26],而植物自身作為自組織有機體,對外界環境適應并非單一性狀的適應,通常是和其他性狀協同適應[27]。形態性狀是植物適應環境變異最直接的外在表現,比葉面積則是植物功能生態學研究中最常用性狀,是植物比較生態學研究中的首選指標[11- 12],可以反映植物獲取資源能力,亦可作為葉遮陰度的指數使用[28]。通過對山梔子(Gardeniajasminoides)和浙江楠(Phoebechekiangensis)在不同光照下葉片特征研究發現,比葉面積隨有效光照強度降低而顯著增加[29- 30]。此外,一般在低光環境下生長的植物,會出現葉片更加寬大,葉長寬比也往往較小的現象[31]。隨著杉木林分郁閉度增加,林下植物自然更新受到弱光限制,從而發生生境過濾作用,通過長期適應與環境選擇出葉性狀與林下弱光環境相適應的原生樹種。本研究中引入喬木的葉長及比葉面積顯著大于原生喬木,表明其受林下弱光環境影響,引入喬木樹種通過增加比葉面積協同葉長調整,來加強捕獲光能,以促進光合作用和維持自身的正常生長發育。本研究表明引入喬木與引入灌木的比葉面積沒有顯著差異,Chen等[32]、劉貴峰等[14]研究也發現喬灌木間的比葉面積沒有明顯差異;而原生灌木的比葉面積大于原生喬木,這與呂金枝等[33]對山西霍山30種不同功能型植物間的葉面積關系一致。本研究中引入樹種與原生樹種比葉面積變化的差異可能與引入灌木在低光環境的自我調節能力的大小和適應策略有關。
變化環境中,植物主要通過兩條適應策略,其一為改變葉性狀,其二則調整植物性狀間的關系[34]。葉綠素與植物光合作用密切相關,本研究中引入喬木的葉綠素a及總葉綠素含量顯著高于原生喬木,說明引入喬木通過增加葉綠素a含量加快光合產物的合成提高自身生產力,以及協同較高的葉面積與比葉面積來適應林下弱光環境,這與梯度弱光脅迫下楊桐(Adinandramillettii)及紫楠(Phoebesheareri)的生理響應一致[35- 36]。然而有研究指出隨著光照減弱,光合色素調節主要以葉綠素b的提高為主[37],低光環境中耐陰植物葉綠素b含量增加的幅度往往高于葉綠素a,從而形成較低的葉綠素a/b,以提高植物對藍光的利用效率[38]。本研究中灌木樹種之間葉形態性狀及葉綠素a含量無顯著變化,這可能反映了引入灌木樹種受林下弱光環境的影響較小,但引入灌木的葉綠素b含量顯著低于原生灌木,說明引入灌木與原生灌木的比葉面積—葉綠素關系格局發生了位移現象,引入灌木對林下弱光的捕獲和利用能力小于原生灌木,原生灌木的適應性可能更強。
碳是植物體內干物質最主要的構成元素,葉碳含量高則表明其比葉重大,光合速率低,生長速率慢,具有較強的防御能力[39]。本研究中引入樹種的葉碳含量顯著大于原生樹種,根據葉碳含量適應策略可初步將耐陰性排序為:引入喬木<原生喬木、引入灌木<原生灌木。葉片氮、磷含量是植物與環境長期適應結果,研究表明在弱光環境中植物可通過增加氮在捕光組分中的含量和分配來提高光能捕獲能力,或通過增加磷的投資比例來增大細胞的分裂能力,進而獲得較大的比葉面積[37]。本研究中比葉面積、葉綠素、葉氮含量、葉磷含量之間的極顯著正相關關系在不同功能群中變化趨勢一致,表明林下植物主要通過形態適應或生理調節促進比葉面積及色素含量的增加以適應林下弱光環境。氮和磷是陸地生態系統植物生長的主要限制因子,可以直接反映植物受養分脅迫的狀態[40]。一些研究者采用氮/磷來診斷植物的元素受限狀況[41],在對歐洲濕地植物的研究發現,當氮/磷<14,植物受氮制約,當氮/磷>16,植物受磷制約[42]。本研究中,大多數植物的氮/磷>16,說明該地區植物主要受磷制約;引入樹種的氮/磷顯著小于原生樹種,表明引入樹種的氮、磷內穩性可能較原生樹種強。有研究表明,內穩性較弱的樹種的適應性更強[43],進一步說明原生樹種的適應性高于引入樹種;而內穩性較高的樹種的養分利用方式可能更加保守[44],因而內穩性較高的引入樹種在弱光環境中更傾向于生存而非生長,這種保守性策略可能是導致其光合產物低于原生樹種的主要原因。
非結構性碳水化合物是植物光合作用的主要產物,包括淀粉和可溶性糖,不僅可以反映植物碳收支狀況[45],亦可表征植物生長和存活的能力以及應對外界脅迫的緩沖能力[46]。淀粉是植物主要的長期能量儲存物質之一,是植物儲存器官中最重要的碳水化合物[47]。可溶性糖是光合作用的直接產物,是植物通過生理調節適應環境變化的重要機制[48]。引入喬木與原生喬木的淀粉、可溶性糖含量及非結構性碳水化合物總量無顯著差異,但引入喬木的可溶性糖含量與淀粉的比值顯著大于原生喬木,說明引入喬木主要以可溶性糖形式參與機體代謝及抵御外界脅迫為主,而原生喬木的光合產物則以淀粉的形式用于能量儲存及自身生長,這一結論輔證了引入喬木的耐陰性小于原生喬木。引入灌木與原生灌木的非結構性碳水化合物總量接近,但引入灌木的可溶性糖含量與淀粉的比值顯著大于原生灌木,表明引入灌木的耐蔭性較原生灌木弱,其亦發動了可溶性糖調節機制,通過增加可溶性糖含量來調節滲透壓、緩沖弱光脅迫壓力。喬木一般處在森林群落中的優勢層,接收到的光照最強,光合作用也最為充分,植物葉片的碳同化能力最大,而灌木處于次優勢層,由于喬木層的遮擋其碳同化能力較弱,因此灌木的非結構性碳水化合物較喬木低[49- 50]。本研究中不論是引入樹種或原生樹種,其中喬木非結構性碳水化合物均顯著低于灌木,這可能是引入或原生的喬木與灌木樹種均處在林下層,灌木能夠進行正常生長,而喬木并未處在適宜的生態位上,林下光照不足制約了光合作用。
在植物生長發育過程中,由于受到物理、生理等因素的綜合作用,不同葉性狀間存在相互促進或制約關系[51]。本研究也發現,引入樹種與原生樹種的葉性狀呈顯著線性相關,喬木或灌木的葉綠素含量與葉形態性狀、葉氮、葉磷含量與葉綠素含量均呈顯著正相關,其淀粉含量均與比葉面積呈顯著負相關,說明相同生活型植物葉性狀對弱光環境的響應具有高度相似性[52],表現出較高的趨同適應特征。另外植物類群和生活型對性狀分異產生一定影響[53],PCA排序結果顯示葉性狀在引入喬木與引入灌木間的差異較小,但原生喬木與原生灌木間葉性狀的差異較大,這反映了不同生活型植物的葉性狀仍具有一定差異。
4 結論
本研究探討了杉木人工純林改造為杉木復層異齡林7年后,林下引入喬灌木樹種與原生喬灌木樹種的葉性狀變異特征,研究發現引入樹種與原生樹種的葉形態性狀差異較小,引入樹種及原生灌木的比葉面積及色素含量顯著高于原生喬木,引入樹種主要通過高比葉面積—葉綠素a來適應林下弱光環境,原生樹種主要采用增加比葉面積—葉綠素b適應策略。引入喬木的葉碳氮磷含量均大于引入灌木,而原生喬木均小于灌木,引入與原生樹種均受土壤磷元素制約。引入灌木的可溶性糖含量最高,原生喬木的淀粉含量最高,灌木的非結構性碳水化合物顯著大于喬木,灌木之間光合產物較為接近,喬木之間無顯著差異。引入樹種與原生樹種的葉性狀高度相關,表現出趨同適應特征。引入喬木與灌木的葉性狀較為接近,引入樹種與原生喬木和灌木葉性狀存在一定差異,這不僅與植物遺傳特性密切相關,也與植物耐陰性和所處生態位有關。綜合葉性狀分析,本研究中引入灌木樹種的適應性優于喬木樹種。因此,在研究區構建杉木異齡復層林需根據林木生態適應特征,進行適當間伐,降低林分郁閉度,將有利于林下植被層的快速恢復,尤其是伴生喬木樹種需要更多的光照資源和生長空間。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
