海拔梯度變化對(duì)中亞熱帶黃山松土壤微生物生物量和群落結(jié)構(gòu)的影響
2019-05-13 02:06:54趙盼盼周嘉聰林開淼張秋芳曾曉敏徐建國(guó)陳岳民楊玉盛
生態(tài)學(xué)報(bào) 2019年6期
關(guān)鍵詞:研究
趙盼盼,周嘉聰,林開淼,張秋芳,袁 萍,曾曉敏,蘇 瑩,徐建國(guó),陳岳民,*,楊玉盛
1 福建師范大學(xué)濕潤(rùn)亞熱帶生態(tài)地理過(guò)程教育部重點(diǎn)實(shí)驗(yàn)室,福州 3500072 福建師范大學(xué)地理研究所,福州 3500073 福建戴云山國(guó)家級(jí)自然保護(hù)區(qū)管理局,泉州 362500
IPCC(2013)報(bào)告指出,21世紀(jì)末全球氣溫將平均上升1.8—4.0℃[1]。全球變暖已是不爭(zhēng)的事實(shí)。當(dāng)前,氣候變化對(duì)生態(tài)系統(tǒng)的影響是生態(tài)學(xué)研究的熱點(diǎn)問(wèn)題之一。而在全球氣候變化與陸地生態(tài)系統(tǒng)關(guān)系的研究中,由海拔高度變化引起的自然溫度梯度的變化,可以用來(lái)模擬氣候變化,這也是近年來(lái)被廣泛應(yīng)用且經(jīng)濟(jì)的方法[2]。其中,土壤微生物是土壤營(yíng)養(yǎng)源或匯,參與污染物的降解和腐殖化過(guò)程[3- 4],在森林生態(tài)系統(tǒng)能量流動(dòng)和養(yǎng)分循環(huán)過(guò)程起著重要作用[5]。因此,利用海拔梯度變化,了解其微生物多樣性的分布格局及其形成機(jī)制是未來(lái)解釋生物多樣性對(duì)氣候變化響應(yīng)的重要的參考依據(jù)。
隨海拔升高,溫度降低,這可能會(huì)影響微生物的活動(dòng)[6]。土壤微生物群落結(jié)構(gòu)組成十分復(fù)雜,數(shù)量巨大,單純的利用傳統(tǒng)培養(yǎng)方法取得的土壤微生物信息往往不能全面地反映自然狀況下的土壤微生物群落結(jié)構(gòu)變化。近年來(lái)磷脂脂肪酸(Phospholipid-derived fatty acids,PLFA)作為微生物的生物標(biāo)記,常被用于鑒定微生物種類和識(shí)別微生物類群[7- 8]。這種方法可以準(zhǔn)確反映微生物群落的實(shí)際情況[9]。
盡管微生物學(xué)方法在不斷改進(jìn),但是,目前國(guó)內(nèi)外關(guān)于土壤微生物群落隨海拔梯度的變化并無(wú)統(tǒng)一定論。如Whitaker等人[10]研究發(fā)現(xiàn)隨著安第斯山脈海拔的升高總PLFA含量,Bacteria和Fungi,GN和GP,F∶B均隨海拔升高而顯著升高。有研究者在阿爾卑斯山觀察到土壤Fungi隨海拔梯度增加也有相似的結(jié)果[11- 13]。而Wagai等人[14]發(fā)現(xiàn),隨著海拔的升高,Bacteria和Fungi 含量下降。Djuki等人[15]研究發(fā)現(xiàn),TPLFA及Bacteria 并未隨海拔升高表現(xiàn)出一致的變化規(guī)律。海拔梯度的變化會(huì)影響植被類型[16],而植被類型是影響土壤微生物群落的重要因素,不同樹種凋落物和根系分泌物直接影響微生物群落組成[17],此外通過(guò)間接影響森林土壤的理化性質(zhì),如pH、有機(jī)質(zhì)含量、土壤結(jié)構(gòu)和微氣候等,進(jìn)而對(duì)土壤微生物群落產(chǎn)生重要影響[18- 19]。王淼等人[20]在罕山研究表明不同海拔下植被類型不同,顯著影響了Bacteria、Fungi、GN和GP生物量以及微生物群落結(jié)構(gòu)組成。由此可知,以往沿海拔梯度的研究大多以不同植被類型作為研究對(duì)象,可能是造成研究結(jié)果不一的原因。因此,控制植被類型變量,沿海拔梯度,探究微生物群落結(jié)構(gòu)的變化,可能更能反映未來(lái)氣候變化下土壤微生物的響應(yīng)。
戴云山國(guó)家級(jí)自然保護(hù)區(qū)是東南地區(qū)重要的生物多樣性的基因庫(kù),物種豐富,種類繁多,現(xiàn)已成為研究的熱點(diǎn)區(qū)域[21- 23]。黃山松(Pinustaiwanensis)又稱臺(tái)灣松,常綠喬木,分布于中國(guó)東南部,高山地綠化和用材的優(yōu)良樹種[24],也是亞熱帶地區(qū)的主要建群種。戴云山是中國(guó)最大的黃山松種質(zhì)基因基地[25],對(duì)保護(hù)福建省乃至我國(guó)生物多樣性和開展中亞熱帶植被的演替具有重要科學(xué)意義。因此,本研究利用PLFA技術(shù),研究3個(gè)不同海拔梯度的黃山松林土壤微生物生物量和微生物群落組成及其影響因素,以期為揭示未來(lái)全球變暖背景下微生物群落變化規(guī)律提供理論依據(jù),也為探究戴云山自然保護(hù)區(qū)土壤養(yǎng)分循環(huán)的過(guò)程提供科學(xué)依據(jù)。
1 試驗(yàn)區(qū)與試驗(yàn)設(shè)計(jì)
樣地位于福建省泉州市戴云山自然保護(hù)區(qū)(25°38′—25°43′ N,118°05′—118°05′ E),該保護(hù)區(qū)地處中亞熱帶向南亞熱帶的過(guò)渡區(qū)域,是中國(guó)單位面積生物多樣性程度最高的自然保護(hù)區(qū)之一。氣候類型為亞熱帶海洋性季風(fēng)氣候,具有冬冷夏熱,水熱同期,濕潤(rùn)多雨等特點(diǎn)。年平均氣溫20℃,年降水量達(dá)1700—2000 mm,年平均相對(duì)濕度80%[26]。不同海拔梯度的黃山松林的主要優(yōu)勢(shì)樹種均為黃山松,林內(nèi)其他喬木較少,林下植被相似,1300,1450 m海拔和1600 m海拔相比灌木較多,林下灌木層優(yōu)勢(shì)樹種以腫節(jié)少穗竹(Oligostachyumoedogonatum)為主,伴生有鈍齒冬青(LlexcrenataThunb)、鹿角杜鵑(Rhododendronlatoucheae)、窄基紅褐柃(Euryarubiginosavar.attenuata)和短尾越桔(Vacciniumcarlesii)等;草本層以里白(Hicriopterisglauca)為主,伴生有德化假衛(wèi)茅(Microtropisdehuaensis)、藜蘆(Veratrumnigrum)、狗脊蕨(Woodwardiajaponica)和黑莎草(Gahniatristis)等,土壤均為花崗巖發(fā)育的黃壤。樣地基本情況詳見表1。

表1 不同海拔土壤樣地基本情況
2 研究方法
2.1 土壤樣品采集
2017年5月,選取戴云山1300、1450、1600 m 3個(gè)海拔的黃山松林,于每個(gè)海拔梯度設(shè)置20 m×20 m的標(biāo)準(zhǔn)樣地,在每個(gè)標(biāo)準(zhǔn)樣地內(nèi)隨機(jī)布設(shè)5個(gè)2 m×2 m小樣區(qū),去除表面凋落物,每個(gè)小樣區(qū)采用“S”型混合采樣方法取樣,取表層土,土層深度約為10 cm,每片林子采集5個(gè)土壤樣品。將采集的土壤樣品迅速帶回實(shí)驗(yàn)室保鮮。去除可見根系、石粒等及動(dòng)植物殘?bào)w后,將土壤過(guò)2 mm篩。然后將土樣用四分法分為兩部分,一部分用于土壤速效養(yǎng)分、含水率、土壤微生物生物量和微生物群落結(jié)構(gòu)的測(cè)定;另一部分土壤室內(nèi)自然風(fēng)干后過(guò)0.149 mm篩,用于測(cè)定土壤全氮磷等基本理化指標(biāo)。
2.2 土壤理化性質(zhì)測(cè)定方法
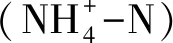
2.3 土壤微生物生物量碳、氮、磷及微生物群落結(jié)構(gòu)(PLFA)測(cè)定
微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸-K2SO4浸提法[28],最后用總有機(jī)碳分析儀測(cè)定提取液中有機(jī)碳含量,用連續(xù)流動(dòng)分析儀測(cè)定總氮含量。MBC計(jì)算公式:BC=ΔEC/kC,式中:ΔEC為熏蒸與未熏蒸土壤有機(jī)碳含量的差值,kC為轉(zhuǎn)換系數(shù),取值0.45。MBN計(jì)算公式:BN=ΔEN/kN,式中:ΔEN為熏蒸與未熏蒸土壤有機(jī)氮含量的差值,kN為轉(zhuǎn)換系數(shù),取值0.54。土壤微生物生物量磷(MBP)采用氯仿熏蒸-NaHCO3浸提法[29],MBP計(jì)算公式為:BP=ΔEP/kP,轉(zhuǎn)換系數(shù)kP取值0.40。
微生物群落結(jié)構(gòu)(PLFA)具體參考Bossio和Scow[30]的方法進(jìn)行樣品的浸提、分離、酯化、萃取,采用MIDI軟件系統(tǒng)分析PLFA各組分含量。具體步驟:稱取相當(dāng)于8 g干重的新鮮土壤,置于離心管中,加入提取液(氯仿∶甲醇∶磷酸緩沖液體積比為1∶2∶0.8)后振蕩2 h,然后在2000g離心10 min,將上清液倒入加有12 mL氯仿和12 mL磷酸緩沖液的分液漏斗中;下層沉淀再用23 mL提取液重復(fù)提取1次。兩次的提取液混合搖勻,靜置過(guò)夜。第2天將分液漏斗中的氯仿層小心接入試管中,在30—32℃水浴中用高純N2吹干。濃縮的脂肪酸用1 mL氯仿分5次轉(zhuǎn)移到活性硅膠柱中,分別用氯仿、丙酮和甲醇洗提,棄去氯仿和丙酮洗提液,收集甲醇洗提液,在32℃水浴中用高純N2吹干后,依次加入1 mL甲醇∶甲苯(體積比1∶1)混合液和1 mL 0.2 mol/L KOH 甲醇溶液,振蕩混勻,在37℃水浴中處理15 min。之后加入0.3 mL 1 mol/L醋酸、2 mL正己烷和2 mL純水,低速振蕩10 min,提取上層溶液;剩余溶液用2 mL正己烷重復(fù)提取1次,兩次提取的上層溶液混合后,用高純N2吹干,得到甲酯化的脂肪酸樣品。該樣品用200 μL正己烷溶解,以19:0甲酯作為內(nèi)標(biāo)物,在氣相色譜儀(Hewlett-Packard 6890 series GC,FD)上采用MIDI軟件系統(tǒng)(MID,IInc.,Newark,DE)進(jìn)行分析,測(cè)定磷脂脂肪酸各組分的含量。具體的土壤微生物種群磷脂脂肪酸標(biāo)志物及其特征比值詳見表2[31-35]。

表2 土壤微生物種群磷脂脂肪酸標(biāo)志物
2.4 數(shù)據(jù)處理
數(shù)據(jù)經(jīng)過(guò)Microsoft Excel 2013軟件處理后,采用SPSS 21.0統(tǒng)計(jì)軟件對(duì)不同海拔土壤各指標(biāo)進(jìn)行統(tǒng)計(jì)分析。采用單因素方差分析(one-way ANOVA)檢驗(yàn)不同處理之間土壤理化性質(zhì),土壤微生物生物量及土壤微生物群落結(jié)構(gòu)的差異顯著性(Duncan法,α =0.05)。采用Canoco Software 5.0軟件,以土壤微生物PLFA為響應(yīng)變量,以土壤理化性質(zhì)及微生物生物量為解釋變量做冗余分析(Redundancy Analysis,RDA);繪圖由Origin 9.0軟件完成。
3 結(jié)果與分析
3.1 土壤基本理化性質(zhì)

3.2 不同海拔對(duì)微生物生物量及其比值的影響
MBC、MBN和MBP含量均隨海拔升高而降低,但是不同海拔之間的MBC含量的差異性未達(dá)顯著水平,而海拔1300 m的MBN和MBP含量顯著高于海拔1450 m和1600 m的MBN和MBP含量(P<0.05)(表4)。隨海拔升高,MBC∶MBN 并無(wú)顯著變化。和1300 m海拔相比,1450 m和1600 m海拔MBC:MBP分別顯著增加了2.68倍,2.12倍,MBN∶MBP分別顯著增加了1.33倍,1.44倍(圖1)。
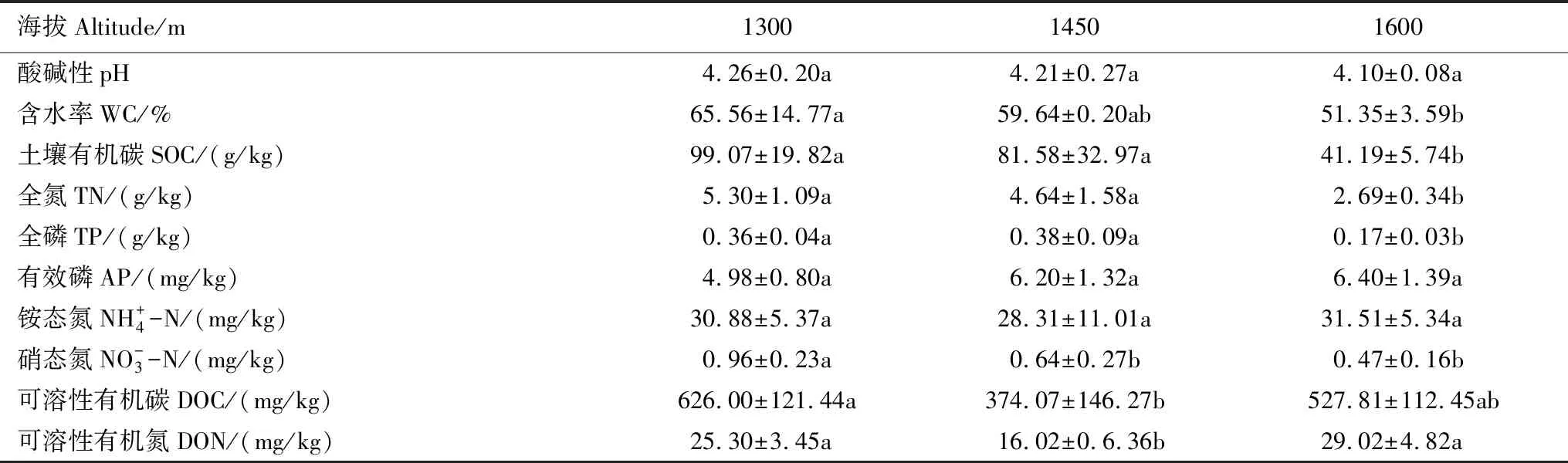
表3 不同海拔土壤基本理化性質(zhì)
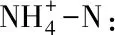

表4 不同海拔對(duì)微生物生物量的影響
表中數(shù)據(jù)為平均值±標(biāo)準(zhǔn)差(n=5),同一列不同小寫字母表示同一土層不同海拔間差異顯著(P<0.05)
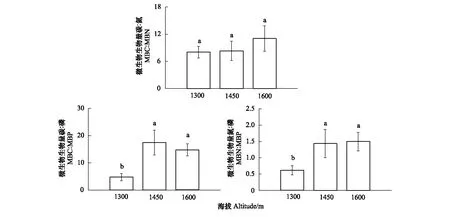
圖1 不同海拔處理下對(duì)土壤MBC∶MBN,MBC∶MBP,MBN∶MBP比值的影響Fig.1 Effects of different altitudes on soil MBC∶MBN,MBC∶MBP,MBN∶MBP ratiosMBC:微生物生物量碳,Microbial biomass carbon; MBN:微生物生物量氮,Microbial biomass nitrogen; MBP:微生物生物量磷,Microbial biomass phosphorus
3.3 不同海拔土壤中特征微生物類群PLFA 總量及其比率
不同微生物相對(duì)生物量在不同海拔土壤中分布不同。其中Bacteria生物量最大,其次是Fungi,ACT。隨海拔升高,AMF,GN,Fungi,TPLFA含量隨之下降(表5),均為1300 m>1450 m>1600 m,和1300 m海拔相比,1600 m海拔GN含量下降了0.37倍。而海拔變化對(duì)GP和ACT無(wú)顯著影響。和1300 m海拔相比,1600 m海拔的GP∶GN顯著增加了34.69%。F∶B比值在1450 m海拔最低,和1300 m海拔的F∶B比值相比,1450 m海拔的F∶B比值顯著下降了33.73%。
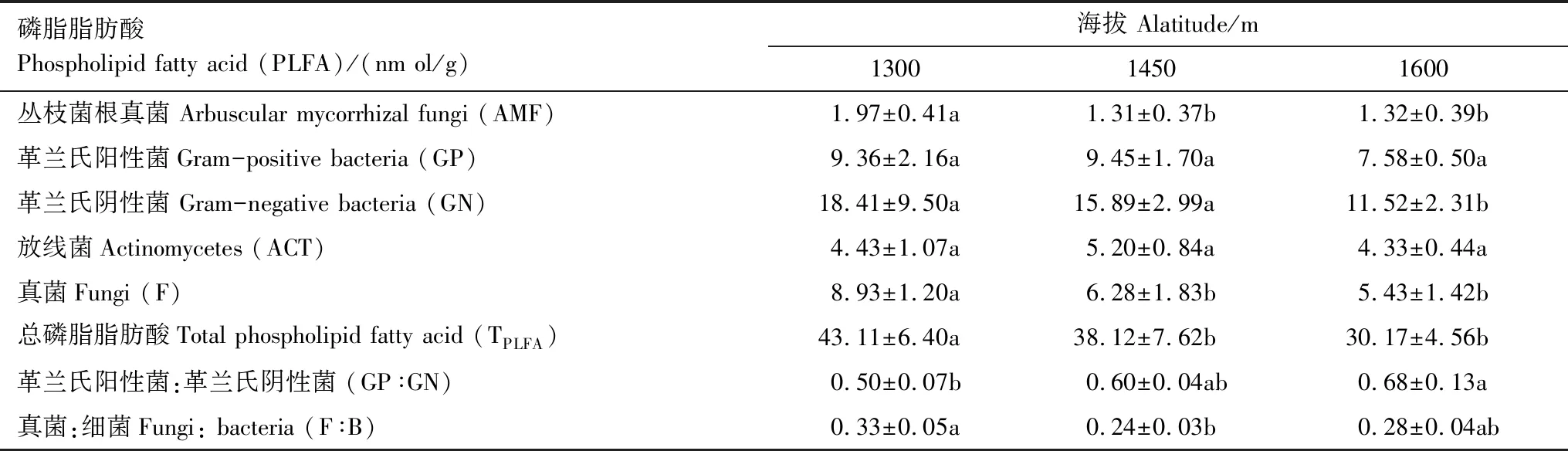
表5 不同海拔對(duì)磷脂脂肪酸及其特征比值的影響
表中數(shù)據(jù)為平均值±標(biāo)準(zhǔn)差(n=5),同一行不同小寫字母表示不同海拔間差異顯著(P<0.05)
3.4 土壤微生物群落結(jié)構(gòu)變化的RDA分析
以土壤微生物PLFA作為響應(yīng)變量,土壤的理化性質(zhì)作為環(huán)境解釋變量進(jìn)行RDA分析(圖2)。不同海拔梯度都按照一定的規(guī)律進(jìn)行了很好的聚類,說(shuō)明不同海拔處理顯著影響了表層土壤微生物PLFA。RDA分析表明T和DON是影響表層土壤PLFA變化的關(guān)鍵因子。RDA第一軸和第二軸分別解釋了變量的56.48%和9.90%,其中氣溫(T)作用最明顯,解釋了土壤微生物群落結(jié)構(gòu)變化的41.1% (P=0.002),且與AMF,Fungi,TPLFA呈顯著正相關(guān),與F∶B顯著負(fù)相關(guān)。
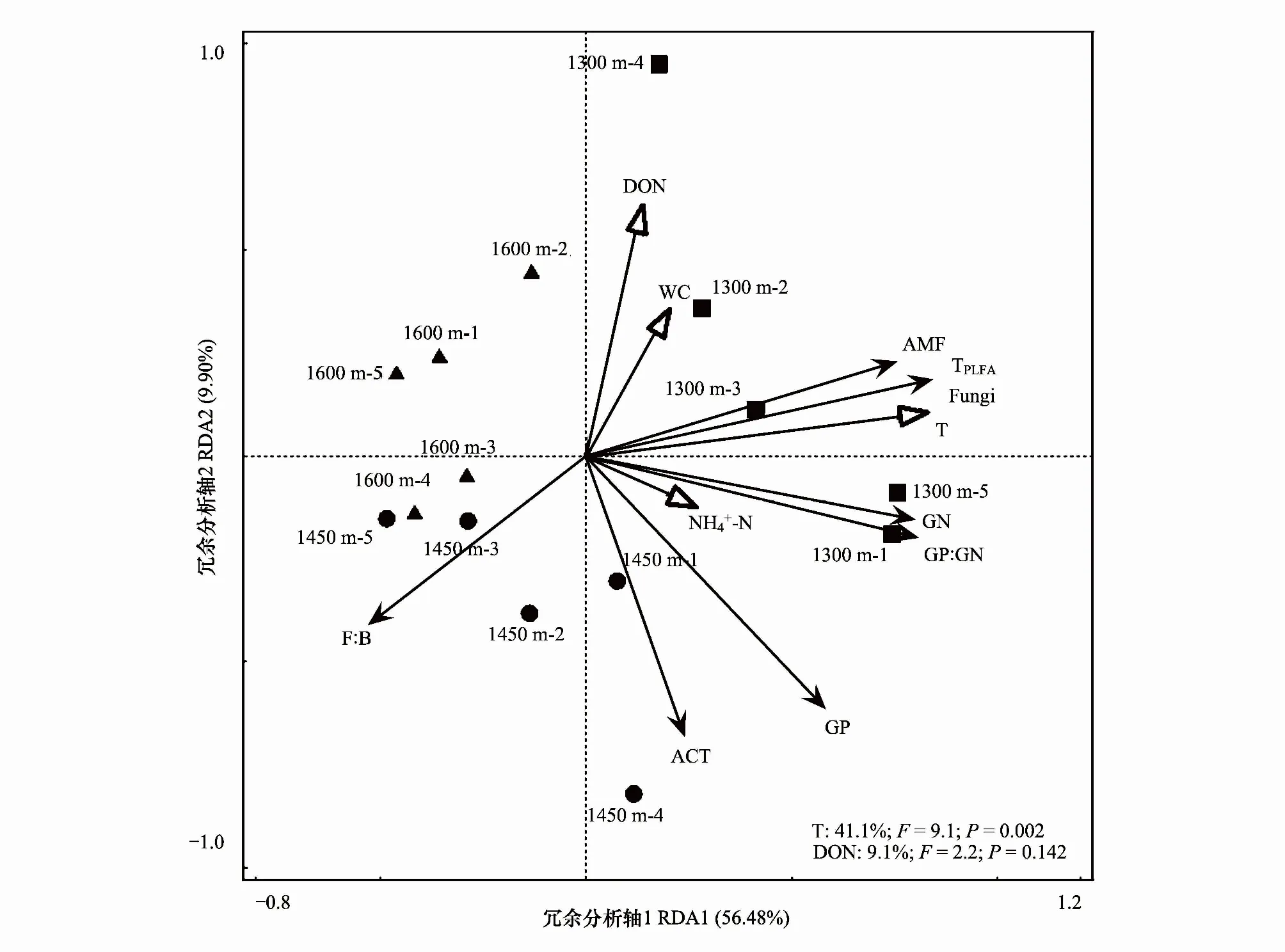
圖2 不同海拔對(duì)土壤微生物群落結(jié)構(gòu)影響的冗余分析Fig.2 Correlations of soil phospholipid fatty acid to soil properties as determined by redundancy analysis (RDA)GN:革蘭氏陰性菌,Gram-negative bacteria; ACT:放線菌,Actinomycetes; AMF:叢枝菌根真菌,Arbuscular mycorrhizal fungi; TPLFA:總磷脂脂肪酸,Total phospholipid fatty acid; GP∶GN:革蘭氏陽(yáng)性菌∶革蘭氏陰性菌,Gram-positive bacteria∶Gram-negative bacteria; F∶B:真菌∶細(xì)菌,Fungi:bacteria; DON:可溶性有機(jī)氮,Dissolved organic nitrogen; WC:含水率,Water content; T:平均年氣溫,Temperature; 銨態(tài)氮,Ammonium nitrogen
4 討論
4.1 不同海拔下微生物生物量及其比值的差異
土壤微生物量是土壤微生物發(fā)揮作用的物質(zhì)基礎(chǔ),由于其周轉(zhuǎn)快、靈敏度高,可以直接或間接反映土壤肥力和土壤環(huán)境質(zhì)量的微小變化[36-37]。MBC和MBN分別是有機(jī)碳和有機(jī)氮中最活躍、最易變化的部分,雖然所占比例很小,卻是整個(gè)生態(tài)系統(tǒng)養(yǎng)分和能源循環(huán)的關(guān)鍵和動(dòng)力[38],而MBP則反映著土壤中有機(jī)-無(wú)機(jī)磷素的轉(zhuǎn)化過(guò)程。本研究中,MBN和MBP均隨海拔降低而顯著升高。這是因?yàn)楹0?300 m地區(qū)溫度較高,有效促進(jìn)了微生物的生長(zhǎng)及活性,加速其對(duì)無(wú)機(jī)養(yǎng)分的利用。同時(shí)促進(jìn)了MBC的積累,但隨海拔下降MBC含量有升高趨勢(shì)并不顯著。這與劉秉儒[39]的研究結(jié)果并不一致,其結(jié)果表明賀蘭山東坡(1400—3100 m)土壤MBC和MBN隨海拔的升高而增加。土壤有機(jī)質(zhì)含量是影響土壤微生物量的重要因素[40- 41],賀蘭山高海拔地區(qū)SOC、TN含量較高,進(jìn)而增加MBC、MBN含量,然而本研究海拔1600 m地區(qū)的SOC、TN含量相比于海拔1300 m地區(qū)更低,因此MBC和MBN含量隨海拔升高下降。
4.2 不同海拔下微生物PLFA的變化及其驅(qū)動(dòng)因子
土壤微生物是土壤生態(tài)系統(tǒng)中的重要組成部分,參與土壤有機(jī)質(zhì)分解,養(yǎng)分循環(huán)及氮的固定[43],是研究和預(yù)測(cè)陸地生態(tài)系統(tǒng)對(duì)全球變化響應(yīng)的重要對(duì)象。隨著全球變化的進(jìn)程,預(yù)計(jì)在本世紀(jì)末,全球高海拔和高緯度生態(tài)系統(tǒng)的氣溫將增加0.6℃[44]。海拔梯度的變化會(huì)導(dǎo)致溫度、光照、水分等多種環(huán)境因子的改變,引起山地區(qū)域小氣候、土壤理化特征等環(huán)境系統(tǒng)的梯度效應(yīng),進(jìn)而影響土壤微生物生物量、活性及群落結(jié)構(gòu)[13,45]。目前,關(guān)于土壤微生物的海拔梯度分布規(guī)律的相關(guān)研究還很少,土壤微生物分布規(guī)律有待進(jìn)一步揭示[13]。
本研究表明,隨海拔升高,AMF,GN,Fungi,Bacteria,TPLFA含量顯著下降。同時(shí),海拔梯度影響土壤養(yǎng)分含量(表3)。土壤養(yǎng)分含量對(duì)微生物群落起著重要的作用,其數(shù)量和質(zhì)量可以影響真菌生長(zhǎng)[46],被認(rèn)為是決定土壤微生物生存、物種組成和代謝的關(guān)鍵因素之一[47]。有研究表明微生物的特征PLFAs含量與養(yǎng)分的可利用性呈現(xiàn)正相關(guān)關(guān)系[48]。本研究中發(fā)現(xiàn)隨海拔梯度變化微生物PLFA與SOC,TN,TP,MBC,MBN,MBP變化趨勢(shì)一致,其含量均隨海拔升高而降低。這與Wagai等人[14]研究結(jié)果一致,隨海拔升高,土壤Fungi、Bacteria含量也下降。其主要原因是高海拔地區(qū)微生物受到底物的限制作用。然而Siles和Margesin[49]研究表明,高海拔地區(qū),細(xì)菌相對(duì)豐度增加,這與該地區(qū)土壤有機(jī)質(zhì)及養(yǎng)分含量更加豐富有關(guān)。本研究中,高海拔地區(qū)C、N、P養(yǎng)分含量減少,這是因?yàn)楹0?600 m處黃山松植株矮小,黃山松凋落物歸還較少,同時(shí)植被的稀疏增加土壤養(yǎng)分淋失的風(fēng)險(xiǎn)。Bardelli等人[50]研究表明不同海拔地區(qū)表層土壤的細(xì)菌和真菌群落均受到總碳含量的顯著影響。因此,微生物豐度的變化可能主要受到養(yǎng)分的限制。
F∶B和 GP∶GN的比值常被用作指示微生物群落結(jié)構(gòu)變化的重要指標(biāo)[51]。F∶B可表征細(xì)菌和真菌兩個(gè)種群的相對(duì)豐富程度消長(zhǎng)狀況[52],F∶B比值越高,表明生態(tài)系統(tǒng)越穩(wěn)定[53]。研究表明,海拔梯度并非是驅(qū)動(dòng)土壤F∶B的關(guān)鍵因子,F∶B隨海拔高度有相似[11]或相反[13,54- 55]變化趨勢(shì)。許多研究認(rèn)為pH的變化是土壤中優(yōu)勢(shì)真菌和細(xì)菌的主要驅(qū)動(dòng)力[49,56]。而Kotas等人[57]研究表明除pH值外,F∶B與土壤有機(jī)質(zhì)和含水量也有很大的關(guān)系。本研究地區(qū),3個(gè)不同海拔pH并無(wú)顯著差異,但有機(jī)質(zhì)含量則顯著不同,1300 m海拔地區(qū)SOC含量富集促進(jìn)真菌生物量增長(zhǎng),進(jìn)而提高真菌菌絲體與土壤有效養(yǎng)分的接觸面積,減少土壤養(yǎng)分流失,因而1300 m海拔地區(qū)土壤生態(tài)系統(tǒng)更穩(wěn)定。
GP和GN對(duì)底物的傾向性及生長(zhǎng)策略的不同,常用于指示環(huán)境變化下,微生物群落結(jié)構(gòu)變化的指標(biāo)[58]。與GN相比,GP更容易適應(yīng)底物有效性低的環(huán)境[59],主要以K策略物種為主。高海拔地區(qū),土壤碳、氮、磷以及速效養(yǎng)分含量較少,資源相對(duì)貧瘠,GP可以較好的適應(yīng)這種不利環(huán)境,其生物量并未發(fā)生較大的改變。而GN生長(zhǎng)則受到抑制,其生物量隨海拔升高而降低。因此造成,隨海拔升高,GP∶GN顯著增加。這與Huang等人[60]在大安山地區(qū)研究結(jié)果一致。
5 結(jié)論
在3個(gè)不同的海拔梯度下,WC、SOC、TN、TP、MBN、MBP和Fungi、Bacteria、TPLFA隨海拔升高顯著下降。隨海拔升高GN:GP顯著增加,在資源相對(duì)貧瘠的1600 m海拔地區(qū),GP比GN更具競(jìng)爭(zhēng)力,而F∶B比值隨海拔升高有下降趨勢(shì),說(shuō)明真菌比細(xì)菌對(duì)海拔的響應(yīng)更敏感。冗余分析表明,T和DON是影響不同海拔梯度表層土壤微生物群落結(jié)構(gòu)發(fā)生變化的主要環(huán)境因子,因此它們制約了1600 m海拔地區(qū)微生物活動(dòng)及分解作用。未來(lái)全球氣候變暖背景下,將改變土壤碳氮磷養(yǎng)分含量及微生物生物量,從而影響土壤微生物群落結(jié)構(gòu)發(fā)生變化,這對(duì)進(jìn)一步深入了解氣候變化對(duì)山地生態(tài)系統(tǒng)土壤養(yǎng)分循環(huán)具有重要意義。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
