貝氏柯克斯體九里株Ⅱ相突變體的克隆與鑒定
2019-05-07 09:46:06何澤民孫志會于永慧羅聲棟王益清王景林姜永強宋立華
生物技術(shù)通訊 2019年2期
何澤民,孫志會,于永慧,羅聲棟,王益清,王景林,姜永強,宋立華
1.病原微生物生物安全國家重點實驗室,軍事科學(xué)院軍事醫(yī)學(xué)研究院 微生物流行病研究所,北京100071;2.解放軍總醫(yī)院第五醫(yī)學(xué)中心 臨床研究管理中心,北京 100039
貝氏柯克斯體(Coxiella burnetii)俗稱 Q 熱立克次體,為革蘭陰性專性細胞內(nèi)寄生菌。貝氏柯克斯體主要通過氣溶膠傳播,引起人類Q 熱(que?ry fever,不明熱),通常表現(xiàn)為流感樣癥狀,感染劑量可低至1 個菌體[1]。九里(Nine Mile)株是貝氏柯克斯體參考菌株,于1935年分離自美國蒙大拿州的一只安氏革蜱[2]。世界各地的實驗室用不同培養(yǎng)方法對此參考菌進行了長期傳代。傳統(tǒng)的貝氏柯克斯體培養(yǎng)方法包括感染動物、雞胚培養(yǎng)和細胞培養(yǎng),直到2009年美國落基山實驗室發(fā)明了貝氏柯克斯體無細胞培養(yǎng)方法[3-4]。
在雞胚或細胞體外培養(yǎng)時,不同貝氏柯克斯體分離株均會發(fā)生不可逆的相變異。九里株的典型相變異是從強毒的Ⅰ相變異為弱毒的Crazy相[5]或無毒的Ⅱ相[6-7]。九里株的Ⅰ相、Crazy 相、Ⅱ相菌分別表達全長脂多糖(lipopolysaccharides,LPS)、含部分 O 抗原的 LPS、完全缺失 O 抗原的LPS[8]。與九里株Ⅰ相菌(RSA 493 克隆)的基因組相比,九里株Ⅱ相菌(RSA 439 克隆)基因組的主要變化是有25 997 bp的刪除區(qū),對應(yīng)20 個基因(CBU_0679~CBU_0698)的缺失[9-10],而 Crazy 相的基因組在Ⅱ相菌刪除區(qū)基礎(chǔ)上有更多的堿基缺失[8,10]。顯然九里株的刪除區(qū)與Ⅱ相變異無明顯關(guān)聯(lián)。長期以來,九里株及其他貝氏柯克斯體分離株的相變異遺傳學(xué)機制不清楚。
九里株Ⅱ相菌的培養(yǎng)特征等與九里株Ⅰ相菌高度相似[11],且致病力低,可以在生物安全Ⅱ級實驗室操作[12],在貝氏柯克斯體研究中被廣泛應(yīng)用。長期以來,研究人員用九里株Ⅱ相菌(RSA 439)開展研究,但以Ⅰ相菌(RSA 493)基因組序列為參考[14],九里株Ⅱ相菌的基因組細微變異缺乏報道。2017年美國科學(xué)家Millar 對九里株Ⅱ相菌(RSA 439,Clone 4)進行了測序,在刪除區(qū)基礎(chǔ)上鑒定了更多的單核苷酸多態(tài)性位點(single nucleotide polymorphisms,SNP)變 異[12]。 2018年Beare 等闡明了 RSA 439的相變異與 CBU_0533 變異有關(guān)[8]。我們用半固體平板克隆方法對我室保存的九里株Ⅱ相菌株進行分離和全基因組測序分析,有助于理解貝氏柯克斯體的體外適應(yīng)性變異,也對今后采用遺傳背景均一的貝氏柯克斯體克隆開展相關(guān)基礎(chǔ)研究有參考意義。
1 材料與方法
1.1 九里株Ⅱ相菌的克隆分離與基因組提取
貝氏柯克斯體九里株Ⅱ相菌株為本室保存。利用國外已發(fā)表的平板克隆方法[4],隨機選擇單克隆連續(xù)克隆2 次,得到CB01 克隆株,將CB01 接種到 ACCM-2 中,在 37℃、2.5% O2、5%CO2的環(huán)境中培養(yǎng),培養(yǎng) 7 d 后 4℃、12 000 r/min離心15 min 收集菌體沉淀,用蛋白酶K 消化-酚氯仿抽提法純化基因組DNA,-20℃保存。
1.2 測序
對提取的基因組DNA 用Illumina Hiseq 2500和PacBio RSⅡ測序系統(tǒng)分別做二代和三代測序,由北京貝瑞和康生物技術(shù)有限公司完成。
1.2.1 PacBio 數(shù)據(jù)產(chǎn)出流程 利用Nanodrop/凝膠電泳技術(shù)檢測樣品,樣品合格后采用標(biāo)準(zhǔn)的PacBio 建庫流程構(gòu)建文庫,建好的文庫按PacBio標(biāo)準(zhǔn)文庫QC 流程進行質(zhì)控,按PacBio 標(biāo)準(zhǔn)上級流程進行。
1.2.2 Illumina 數(shù)據(jù)產(chǎn)出流程 利用Nanodrop 測D260nm/D280nm和凝膠電泳技術(shù)檢測樣品,樣品合格后采用標(biāo)準(zhǔn)的Illumina 建庫流程構(gòu)建文庫,建好的文庫采用AB 公司的stepONE plus QPCR 儀進行質(zhì)檢,當(dāng)文庫檢測為單峰,且濃度高于3 nmol/L,體積大于25 μL 判定為合格,按照Illumina 標(biāo)準(zhǔn)Cluster 生成、Pooling 和定量方法進行樣本文庫混合,并制備Cluster。
1.2.3 測序反應(yīng) 由 Bluepippin 系統(tǒng)(Sage Sci?ence 公司)生成 20 kb SMRTbell 庫,利用 PacBio RSⅡ系統(tǒng)進行單分子實時讀長測序。
1.3 測序結(jié)果分析
對三代測序的結(jié)果進行拼接,然后用二代測序結(jié)果清洗三代,得到全基因組序列。在Patric(https://www.patricbrc.org/)上對全基因進行整體分析。
1.4 基因組比對
從NCBI下載RSA 439和RSA 493基因組序列及注釋(https://www.ncbi.nlm.nih.gov/genome/ge?nomes/543),用Snippy軟件(https://github.com/tseemann/snippy)將CB01序列分別與RSA 493和RSA 439的基因組序列進行比較分析,找出SNP并對差異基因功能進行注釋。在NCBI(https://blast.ncbi.nlm.nih.gov)上分別將CB01 基因組與RSA 439 基因組和RSA 493 基因組進行比較,找到大片段差異。
2 結(jié)果
2.1 CB01全基因組測序結(jié)果
貝氏柯克斯體是專性胞內(nèi)寄生菌,但其基因組富含插入序列[14]。以插入序列IS1111 為例,該序列編碼轉(zhuǎn)座酶,其拷貝數(shù)高達56 個。由于貝氏柯克斯體基因組中重復(fù)序列多,結(jié)構(gòu)復(fù)雜,普及應(yīng)用的二代測序技術(shù)無法獲得完整基因組序列。為了解CB01的完整基因組結(jié)構(gòu),首先利用PacBio 三代測序技術(shù)進行測序并獲得完整的基因組框架。意外的是,我們發(fā)現(xiàn)該測序技術(shù)在分析貝氏柯克斯體序列中的重復(fù)堿基時易發(fā)生錯誤的單堿基缺失。因此,又采用二代測序技術(shù)對三代測序結(jié)果進行單堿基缺失校正。校正后的CB01 基因組全長2.024 Mb,GC 含量為42.68%,包含1 個1.987 Mb的環(huán)狀染色體和1 個37.321 kb的質(zhì)粒。染色體包含2213 個蛋白編碼區(qū)(CDS)、87 個重復(fù)區(qū)域、41 個 tRNA 編碼區(qū)和 3 個rRNA 編碼區(qū);質(zhì)粒包含54 個CDS。環(huán)狀染色體基因組模式如圖1所示。
2.2 與九里株Ⅰ相菌RSA 493基因組的比較分析
與RSA 493的基因組進行比對,CB01 染色體上存在23 個 SNP,包括14 個堿基替換(表1)、7 個堿基缺失(表2)和1 個位于基因間區(qū)的堿基插入。CBU_0533 基因中的單堿基缺失是九里株發(fā)生Ⅱ相變異并缺失O 抗原的遺傳學(xué)原因[9]。因此測序結(jié)果在遺傳學(xué)上證實了CB01 為Ⅱ相菌克隆。另外,質(zhì)粒上存在1 個同義突變(表1)。
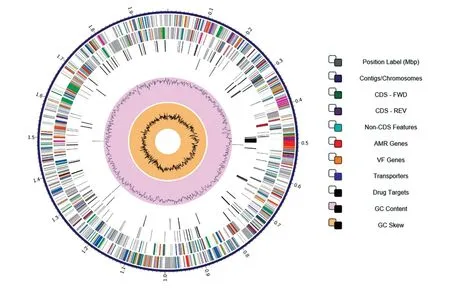
圖1 貝氏柯克斯體九里株CB01克隆的染色體模式圖
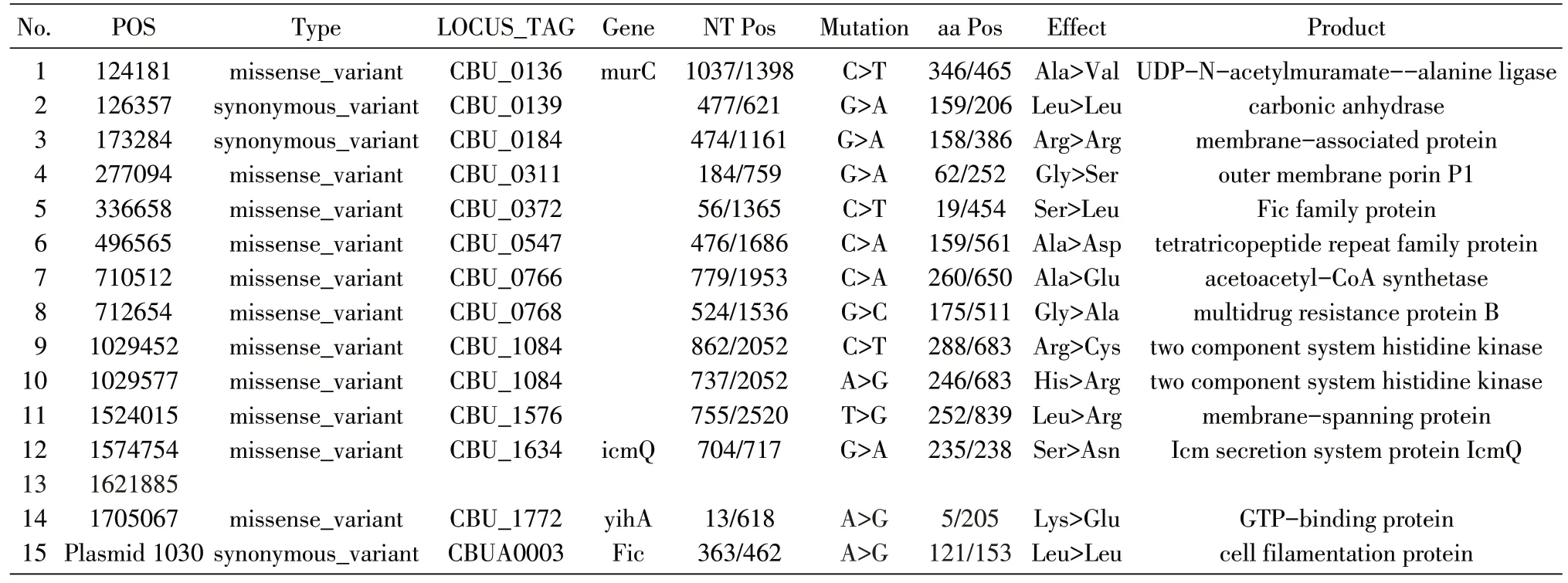
表1 堿基替換統(tǒng)計表
表2 缺失堿基統(tǒng)計表
表3 堿基替換統(tǒng)計表
表4 堿基缺失統(tǒng)計表
2.3 與九里株Ⅱ相菌RSA 439基因組的比較分析
與RSA 439的基因組進行比對,CB01 存在6個SNP,包括4 個堿基替換(表3)和2 個堿基缺失(表4)。CB01的質(zhì)粒與Ⅰ相 RSA 493 相比有 1 個同義SNP,與Ⅱ相菌RSA 439 相比有2 個非同義SNP 差異和1 個同義SNP。測序結(jié)果說明CB01的適應(yīng)性生長更大的可能性與染色體變異有關(guān),而國外Ⅱ相菌RSA 439的體外適應(yīng)性生長可能同時與染色體和質(zhì)粒變異相關(guān)。
2.4 大片段差異
與九里株Ⅰ相菌RSA 493的基因組相比,CB01 基因組有一段刪除序列,大小為25 997 bp,對應(yīng) RSA 493 基因組的 620 667~646 663 堿基,導(dǎo)致 CBU_0679~CBU_0698 基因缺失。CB01 基因組上還存在5 個重復(fù)序列,大小分別為1562、4998、4117、3351 和 3831 bp,共包含 20 個基因。與九里株Ⅱ相菌RSA 439 相比,CB01 同樣多出這5 個重復(fù)序列。
3 討論
貝氏柯克斯體九里株自1935年被首次培養(yǎng)后在國際上廣泛流傳,是貝氏柯克斯體研究用參考菌株。我國的九里株引自前蘇聯(lián)。由于培養(yǎng)困難,易發(fā)生交叉污染等原因,有必要對研究用菌株進行遺傳學(xué)鑒定。我們利用高通量測序技術(shù)完成了貝氏柯克斯體九里株CB01 克隆的完整基因組測序,發(fā)現(xiàn)CB01的菌株甄別位點與九里株Ⅰ相完全相同,可鑒定CB01 為九里株克隆。與九里株Ⅰ相RSA 493 克隆相比,CB01 存在刪除區(qū)變異和CBU_0533 基因變異,這些變異與九里株Ⅱ相菌RSA 439 克隆的變異完全相同,這可以將CB01 進一步鑒定為Ⅱ相菌。九里株Ⅱ相菌CB01 克隆可用做國內(nèi)貝氏柯克斯體研究用參考菌株。
與九里株Ⅱ相菌RSA 439 相比,CB01 至少有6 個SNP 差異和5 個重復(fù)序列差異,其中3 個SNP位于質(zhì)粒。可以明確CB01 是不同的九里株Ⅱ相克隆。貝氏柯克斯體菌的體外適應(yīng)性變異機制還缺乏研究,不同的適應(yīng)性變異可能與不同的培養(yǎng)體系有關(guān)。我們的測序結(jié)果可說明九里株體外適應(yīng)性變異的多樣性。國外已對貝氏柯克斯體脂多糖O 抗原的合成機制進行了較詳細的遺傳學(xué)分析。考慮到其他分離株的相變機制還不清楚,以及存在LPS 合成基因之外的適應(yīng)性變異,本研究也提示有必要深入探討貝氏柯克斯體Ⅰ相菌的相變異和適應(yīng)性變異機制。
