微粒體環氧化物水解酶1和谷胱甘肽S-轉移酶P1與中國北方漢族慢性阻塞性肺疾病及其肺功能表型的相關性
2019-04-26 06:59:40安立林英翔張鴻逯勇張曙華琳
中國老年學雜志 2019年8期
安立 林英翔 張鴻 逯勇 張曙 華琳
(1首都醫科大學附屬北京朝陽醫院呼吸與危重癥醫學科 北京市呼吸疾病研究所 北京市呼吸與危重癥診治工程技術研究中心,北京 100020;2首都醫科大學生物醫學工程學院數學與生物信息學教研室)
慢性阻塞性肺疾病(COPD)是一復雜的多基因疾病。其中,微粒體環氧化物水解酶(EPHX)1、谷胱甘肽S-轉移酶(GST)P1都屬于異生物代謝酶基因,主要參與對香煙煙霧等外來異生型物質、氧化物和反應性氧化物的中間產物在肺中的首過代謝。EPHX1有兩個常見的突變位點Try113His和His139Arg,分別被稱為慢等位基因和快等位基因,能夠引起酶活性的改變〔1〕。而GSTP1常見的突變位點位于外顯子5(Ile105Val)和外顯子6(Ala114Val)。研究證實105Ile等位基因較105Val有更高的催化活性〔2〕。近年來人們對這兩個基因與COPD易感性的關系進行了探討,發現結果存在明顯的種族差異〔3~6〕,而在中國人群中的研究中也有不一致現象,研究多集中于上述幾個突變位點〔7~9〕。本研究旨在探討在中國北方漢族人群中EPHX1、GSTP1與COPD發病及肺功能表型之間的關系。
1 材料與方法
1.1研究對象 收集2007年10月至2009年3月北京朝陽醫院及各合作醫院呼吸科門診就診的穩定期漢族COPD患者316例(病例組)。入選標準:①年齡40~75歲,男女不限;②肺功能檢查符合COPD診斷標準。且符合應用支氣管擴張劑后第1秒用力呼氣容積(FEV1)/用力肺活量(FVC)≤70%和FEV1≤80%pre;③所有患者處于疾病的穩定期,入選前6 w內沒有咳嗽、咳痰或呼吸困難加重;沒有發熱;沒有針對呼吸癥狀加重的用藥改變;④不伴有以下任何一種疾病或情況:哮喘、過敏性鼻炎、支氣管擴張、囊性纖維化、神經肌肉異常等影響運動的疾病及精神異常和認知障礙。同期在北京朝陽醫院和各合作醫院的體檢門診及周圍社區選擇年齡40~75歲,性別匹配的漢族健康對照者213例。所有的受試者在試驗前簽署知情同意書。
1.2DNA的提取及基因分型 所有受試者于采血前均禁食12 h,晨起空腹抽取靜脈血3 ml,經乙二胺四乙酸(EDTA)抗凝,離心(3 000 r/min,15 min)分離血漿,低滲法破壞血細胞沉淀中的紅細胞,將所得白細胞沉淀于-80℃保存,用于提取DNA。應用Illumina公司提供的BeadXp ress的基因分型平臺采用等位基因特異性雜交法進行EPHX1和GSTP1的SNP檢測。
1.3SNPs位點的選擇 由于在人類的基因組中天然存在連鎖不平衡(LD),因此應用較少的標簽SNPs(Tag SNP)便足以能夠獲取存在于高度連鎖區域的遺傳變異信息〔10〕。挑選Tag SNPs的步驟如下:首先我們列出在以往文獻中曾經報道的關于4個基因中和COPD易感及表型相關的SNPs位點EPHX1:rs1051740、rs2234922、rs1051741、rs2292558、rs3728042、rs3753661、rs3766934、rs1009668、rs868966、rs2292566、rs1877724、rs2260863、rs3753658;GSTP1:rs1695、rs1799811、rs612020、rs11227884〔3,5,11~15〕。其次,下載HapMap數據庫亞洲人群〔中國漢族(CHB);日本人群(JPT);選用兩個人群可以增加Tag SNP的覆蓋度〕的EPHX1和GSTP1兩個基因的基因型數據。使用軟件Haploview(Haploview4.2;release 23;CHB+JPT panel),在每個基因中依據它們自身能夠探及LD區塊的能力,挑選出可以覆蓋該基因的Tag SNPs,并將其作為背景數據庫。再次,將陽性位點強行納入Tag SNPs列表中(EPHX1:rs868966、rs1877724、rs3766934、rs1051740、rs2234922、rs2292558、rs1009668;GSTP1:rs1695)。最后,在pairwise 模型下,設定r2=0.8,然后運行Haploview,計算出覆蓋整個基因所需的所有Tag SNPs。
1.4統計分析 采用SPSS16.0軟件進行獨立樣本t檢驗、χ2檢驗。Hardy-Weinberg平衡檢驗采用χ2檢驗。各SNPs位點與COPD發病風險的相關性采用二分類Logistic 回歸分析,與肺功能表型的相關性采用直線回歸分析,并對年齡、性別和吸煙指數等混雜因素進行校正。回歸模型分別采用顯性遺傳模型(AA/AC vs.CC)、隱性遺傳模型(AA vs.AC/CC)和加性遺傳模型(AA vs.AC vs.CC)。對各SNPs位點與COPD發病及肺功能表型之間的所有相關性分析皆進行Bonferroni 多重檢驗〔16〕。上述檢驗均采用全基因組關聯分析軟件PLINK1.06來完成。
2 結 果
2.1受試者一般臨床資料 根據基因分型的結果,病例組最后納入研究的COPD患者為310例,對照組為203例。兩組FEV1、FEV1%pre、FEV1/FVC、FVC有統計學差異(P<0.001)。病例組年齡和吸煙指數顯著高于對照組,體重指數顯著低于對照組(P<0.05)。見表1。
2.2EPHX1及GSTP1的基因多態性與COPD易感性的關系 兩個基因13個SNPs的等位基因頻率分布皆符合Hardy-Weinberg平衡,各SNPs的最小等位基因頻率在病例和對照組中的分布情況見表2。EPHX1的1個SNP位點(rs3766934),GSTP1的1個SNP位點(rs36211088)的等位基因分布在兩組中有明顯差別(P<0.05)。之后以是否發病為因變量,以各個SNPs位點的基因型作為自變量進行二分類Logistic回歸分析,并對年齡、性別和吸煙指數進行校正,結果顯示:在顯性、隱性和加性遺傳模型下EPHX1的2個SNPs位點(rs3738040和rs3766934)、GSTP1的1個SNPs位點(rs36211088)皆與COPD易感性相關(P<0.05)。經Bonferroni多重檢驗后各SNPs位點與COPD發病的相關性消失,見表3。
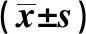
表1 研究對象的臨床特征

表2 兩基因SNP標簽物理位置及等位基因頻率在病例組和對照組中的分布情況

表3 兩基因各SNPs位點與COPD易感性的相關性分析
PBon:經過Bonferroni校正后的P值;NA:等位基因頻率太低而無法進行分析;表4、5同
2.3EPHX1和GSTP1的基因多態性與基線肺功能的關系 如表4、5所示:在顯性模型下,EPHX1的2個SNPs位點(rs3738040和rs3766934)、GSTP1的1個SNP位點(rs36211088)分別與FEV1值和FEV1/FVC值顯著相關(P<0.05)。經Bonferroni多重檢驗后發現EPHX1的rs3766934仍然與FEV1和FEV1/FVC顯著相關(P<0.05),rs3738040則僅與FEV1/FVC顯著相關(P<0.05)。即攜帶rs3766934T基因的患者與更高的FEV1和FEV1/FVC值相關,攜帶rs3738040A的患者僅與更高的FEV1/FVC值相關。顯性模型下GSTP1的rs36211088仍顯示與基線FEV1明顯相關(P<0.05),而與FEV1/FVC的相關性消失(P>0.05)。
2.4LD特征分析與單體型分析
2.4.1LD分析 將對照組中各基因所有樣本的基因型數據導入HaploView(V4.2),分析各基因的所有SNPs位點之間的LD特征并用其構建出每個基因的單體型圖(圖1)。EPHX1上的2個陽性位點rs3738040和rs3766934之間處于強連鎖(D′=0.83)。由于GSTP1的位點數太少,只能看到各SNPs位點間的LD關系,而無法構建連鎖區塊。

表4 兩基因各SNPs位點與FEV1的相關性分析
僅顯示了P<0.1 的SNPs;表5同

表5 兩基因各SNPs位點與FEV1/FVC的相關性分析

共使用了8個常見變異位點(common SNP)構建了EPHX1的LD圖譜。連鎖區塊內的數值顯示的是兩兩位點間的D′值。紅色代表D′=1,白色代表D′=0,而中間的過渡色代表 0 2.4.2單體型分析 應用滑動窗方法對相鄰3個SNPs構成的單體型進行分析,以探討可能與COPD發病和肺功能表型相關的區域。結果顯示,在EPHX1基因中CAT和CAA個單體型與COPD的發病相關,其中CAT與COPD發病更為密切(Pspecific=0.014)。在GSTP1基因中AGG與COPD易感性密切相關(Pspecific=0.010,Pglobal=0.036)。見表6。在與肺功能表型相關的分析中發現EPHX1基因中有4個單體型(GGA、GAA、AAG、AGA)分別與FEV1和FEV1/FVC顯著相關(P<0.05)。GSTP1中有1個單體型(AGG)與FEV1和FEV1/FVC明顯相關(P<0.05),見表7。上述單體型中都包含有前述的單點分析的陽性位點。 表6 兩基因與COPD發病的單體型分析 僅列出了P<0.05的單體型,Pspecific值是指在3個位點構成的單體型中最小的P值;Pglobal值是指3個位點構成的單體型中總體P值;單體型組合中的SNPs的序號與表2中各基因的SNPs序號一致;下表同;F_A:在COPD組的單體型頻率,F_U:在對照組中的單體型頻率 表7 兩基因與基線肺功能的單體型分析 EPHX1是由支氣管上皮細胞表達的具有解毒功能的異物代謝酶。EPHX1基因位于1q42.1染色體,已經較為明確的基因多態性表現在外顯子3(rs1051740)和外顯子4(rs2234922),外顯子3的基因突變型為T-C替換,組氨酸取代113位的酪氨酸(Try113-His),酶活性減少50%(慢等位基因型);外顯子4的突變為A-G替換,精氨酸取代139位的組氨酸(His-Arg),酶活性增強25%(快等位基因型)。Smith等〔3〕首次報道EPHX1的基因多態性與肺氣腫的發病相關,并發現慢等位基因頻率在肺氣腫患者中的發生率(28%)明顯高于對照組(6%)。然而,之后其他研究均未發現外顯子3和4的等位基因頻率在COPD組和對照組中有明顯差別〔17~19〕。目前對于中國北方漢族人的研究亦有不同的結果,張榮葆等〔8〕的研究表明EPHX1的基因多態性與中國COPD的易感性無相關性,但肖丹等〔9〕的研究表明EPHX1的基因多態性與中國COPD的易感性相關,并且EPHX1的基因型與吸煙之間有交互作用。但是鑒于這2個研究的樣本量較少,EPHX1是否為中國人群中COPD的易感基因尚需更大的樣本來驗證。 對于上述較為關注的2個SNPs位點,本研究未發現其與COPD易感性及肺功能表型相關。而另2個以前鮮有報道的SNPs位點rs3738040和rs3766934則分別顯示與COPD發病及肺功能表型顯著相關。在經過Bonferroni多重檢驗后,除rs3738040與FEV1相關性不顯著外,這2個位點仍顯示與肺功能表型顯著相關。Bonferroni多重檢驗是目前在多位點關聯分析中為降低統計學方面的假陽性而進行多重檢驗的常用方法〔16,20〕。Rs3738040位于啟動子區,而rs3766934位于內含子1,由本研究數據(研究樣本和基因型)所構建的包含8個SNPs位點的EPHX1基因LD圖譜看,兩者處于強連鎖,D′為0.83,上述2個SNPs位點均位于第1個強連鎖區塊內(Block 1)。上述2個位點所在的區域,位于RNA選擇性轉錄剪切區〔21〕。研究發現〔21〕,位于此區域的一剪切突變位點(外顯子1)在肝臟中呈高表達,而另一變異體也同時在機體的其他部位包括肺臟中表達。每一個剪切變異體都有自己的5′端啟動子區,提示這個區域在調節EPHX1 mRNA水平方面存在組織特異性。目前已有研究表明,此區域與COPD患者的某些數量性狀密切相關〔11〕。本研究中的2個陽性SNPs位點具體通過何種途徑對COPD及其肺功能表型進行影響尚需進一步探討。 此外,單體型分析對識別某些不能直接觀察到的致病位點有一定的優勢。研究表明,探討基因和疾病相互關系時,應用單體型信息和用SNPs為基礎的單點分析相比,其可信度可增加15%~50%〔22,23〕。因此,在相關分析中聯合應用單點分析和單體型分析對結果的判斷更有效〔24〕。綜合單點分析和單體型分析的結果推測,中國北方漢族人群中與肺功能表型相關的位點可能位于Block 1。 GSTP1屬于GSTs同工酶家族,也是一種重要的抗氧化解毒酶。由于在肺臟組織中的表達較其他GSTs更豐富,因此被認為是在肺臟局部對異生物質解毒的主要GSTs同工酶〔25〕。它的外顯子5、6存在多態性。在第5外顯子(rs1695)發生的單個堿基替換(GTC/ATC,G-A)會導致105位異亮氨酸(Ile)替換為纈氨酸(Val);第6外顯子發生(GCG/GTG,C-T)替換,導致114位的氨基酸由丙氨酸(Ala)替換為Val。研究進一步證實GSTP1純合野生型(105 ILe/ ILe)的催化活性顯著高于雜合型(105 Ile/ Val)和純合變異型(105 Val/ Val)。GSTP1的基因型分布在不同種族間差異極大。Ishii等〔5〕在病例-對照研究中發現GSTP1 105位Ile純合子的比例在COPD患者較對照組明顯升高。而Yim 等〔6〕在研究中卻沒有發現陽性。本研究同樣未發現rs1695與COPD易感相關,但發現另一與COPD發病及肺功能表型顯著相關的SNP位點——rs36211088,且在經過Bonferroni多重檢驗后仍有統計學意義。本研究發現,在HapMap數據庫及本研究數據構建的LD圖譜中,此位點與其他SNPs位點的連鎖程度較弱。此外,雖然單體型分析表明包含rs36211088A,rs4147581G,rs1695G的單體型頻率在病例和對照組中有明顯差別(Pspecific=0.010),但AGG單體型頻率極低(Fre=0.02)。而且與肺功能表型的相關性分析中也是同樣的結果。故此位點在COPD發病中的作用尚需進一步探討。 綜上,本研究發現在中國北方漢族中,異生物代謝酶EPHX1和GSTP1的基因多態性可能與肺功能表型相關。本研究結果與其他研究結果不一致的原因可能有以下幾點:①不同種族的遺傳背景不同,不同人群各SNPs位點間的LD程度和形式不同等。②在不同研究人群內部基因與基因,基因與環境等之間的相互作用都是其影響因素。本研究結果有望進一步深化COPD發病的分子機制研究,豐富國人COPD遺傳易感機制方面的知識。本研究尚存在一定缺陷,樣本量不夠大,同時未能進行重復人群的驗證。

3 討 論
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
