巨噬細胞炎癥蛋白-1α通過骨硬化蛋白抑制骨髓瘤骨病患者成骨細胞功能研究
2019-04-25 12:44:26王曉桃何玉嬋王航飛陽少芳張俊艷高彤彤吳春葉
重慶醫學 2019年8期
關鍵詞:檢測
王曉桃,何玉嬋,王航飛,陽少芳,張俊艷,高彤彤,吳春葉
(桂林醫學院第二附屬醫院血液內科,廣西桂林 541199)
骨髓瘤骨病(myeloma bone disease,MBD)是多發性骨髓瘤(multiple myeloma,MM)患者常見的并發癥。MBD以進行性骨質破壞、多發溶骨性病變等為特征,是MM患者致殘的主要原因。研究表明,有70%左右的患者因MBD而危及生活質量并增加死亡風險,給家庭及社會造成沉重負擔[1]。因此,深入研究MBD發生機制,可為靶向治療MBD奠定理論基礎。
MBD發生的主要機制是在骨髓微環境中,瘤細胞與骨髓基質細胞(bone marrow stromal cells,BMSCs)通過多條信號通路導致破骨細胞(osteoclast,OC)活性增強和成骨細胞(osteoblast,OB)活性減弱,造成溶骨性損害,阻礙新骨生成[2-3]。本課題組的前期研究已證實,巨噬細胞炎癥蛋白-1α(macrophage inflammatory protein-1α,MIP-1α)能激活OC,介導瘤細胞生長,并在MBD的發生、發展中發揮重要作用[4]。最新研究發現,MIP-1α有抑制OB的作用[5],但其具體機制不明確。筆者前期研究發現,大部分MM患者骨髓中MIP-1α和骨硬化蛋白(sclerostin,Scl)水平增高,兩者呈明顯的正相關[6],Scl是Wnt/β-catenin通路的重要抑制劑[7],β-catenin與T細胞因子相互作用,誘導OB堿性磷酸酶(alkaline phosphatase,ALP)的活性,促進前體OB的生長和早期分化、細胞外基質成熟和礦化[8]。因此,本研究擬通過體外實驗驗證MIP-1α促進瘤細胞表達Scl,從而抑制OB的分化與成熟、細胞外基質成熟和礦化促進OB凋亡,導致MBD的發生、發展。
1 資料與方法
1.1一般資料 選擇來自2015年1-12月本科室新確診的6例MBD患者,其中男4例,女2例,中位年齡64歲(47~73歲)。入選病例均按照國際骨髓瘤工作組的診斷標準確診為活動性MM,如并發MBD癥狀的MM患者均需行X線片檢查骨盆、顱骨、肱骨、股骨等確診不同程度的MBD,如無MBD相關癥狀的MM患者需行全身低劑量CT或胸椎、腰椎MRI以明確有無并發MBD。所有入選患者均簽署知情同意書。細胞株:高表達MIP-1α和Scl的人骨髓瘤H929細胞(購自天津血液病研究所[6])及MBD患者BMSCs分化成熟的第3代OB為研究對象。
1.2方法
1.2.1主要試劑和儀器 胎牛血清、小牛血清、DMEM培養基、RPMI-1640干粉均購自杭州四季青公司,淋巴細胞分離液、堿性磷酸酶(Alkaline phosphatase,ALP)測定試劑盒(南京建成生物工程研究所第一分所),Bradford法蛋白定量試劑盒(上海捷瑞生物工程有限公司),二甲基亞砜(DMSO)、地塞米松、β-甘油磷酸鈉、維生素C、茜素紅染色液(美國Sigma公司)。ALP改良鈣鈷法染液(南京建成科技有限公司);MIP-1α和Scl ELISA試劑盒(美國Gibco公司);MIP-1α和Scl蛋白,鼠抗人MIP-1α、Scl、IL-6和ALP單克隆抗體,免疫組織化學試劑盒、DAB顯色試劑盒,鼠抗人GAPDH單克隆抗體(北京中杉金橋生物技術有限公司);Annexin V FITC細胞凋亡測定試劑盒(美國R&D公司);凝膠電泳儀、酶標儀(美國 Bio-Rad公司),IX73 倒置熒光顯微鏡(日本Olympus公司)。
1.2.2實驗分組 實驗分5組:對照組:常規培養基培養;MIP-1α組:培養基加0.2 ng/mL MIP-1α標準品;Scl組:培養基加0.2 ng/mL Scl標準品;MIP-1α單抗(MIP-1α-Ab)組:培養基加0.2 ng/mL MIP-1α單抗;sclerostin 單抗(Scl-Ab)組:培養基加0.2 ng/mL Scl單抗。
1.2.3純化并原代培養OB并鑒定 從MBD患者髂后上棘處抽取骨髓5~10 mL,肝素抗凝,結合全骨髓培養法,用等量的PBS稀釋,按1.0∶1.5比例加入紅細胞裂解液,680×g離心20 min,去除上清液,移至無菌培養瓶中,補足完全培養基(15%胎牛血清,1%雙抗,DMEM)培養。5 d后首次換液,并在顯微鏡下觀察細胞開始變為梭形、貼壁,后每3天換液1次,待細胞長滿視野70%~80%后首次傳代,經0.25%胰酶消化,并接種于六孔板,改為OB誘導分化培養基(1×10-8mol/L地塞米松、50 g/mL抗壞血酸及10 mmol/L β-甘油磷酸)。每3天換液1次,并在顯微鏡下觀察細胞形態變化,行茜素紅染色,ALP免疫組織化學鑒定為OB后方可進行傳代培養并留做后續實驗。
1.2.4茜素紅染色鑒定并觀察鈣離子沉積結節 BMSCs分化的OB培養14 d后,換液,種至6孔板,PBS沖洗2次,4%多聚甲醛固定30 min后,再用PBS沖洗3次,用0.1%茜素紅-Tris-HCl染色30 min,PBS沖洗,干燥,封固后,倒置相差顯微鏡下觀察鈣結節。
1.2.5OB的定性檢查 改良Gomori鈣鉆法ALP染色,倒置相差顯微鏡觀察。ALP陽性細胞表現為細胞質出現棕黑色片狀顆粒。
1.2.6H929細胞傳代培養 H929細胞在含10%澳洲胎牛血清(美國Gibco公司)、5%小牛血清、100 U/mL青霉素、100 μg/mL鏈霉素的RPMI-1640培養基中,37 ℃、5% CO2細胞培養箱中懸浮培養。
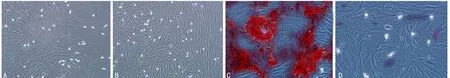
A:BMSCs分離純化培養10 d;B:BMSCs分離純化培養14 d;C:茜素紅染色BMSCs分離純化培養14 d的OB;D:ALP染色BMSCs分離純化培養14 d的OB
圖1 BMSCs分離培養純化OB(×40)
1.2.7ELISA定量檢測H929細胞上清液中的MIP-1α和Scl水平 取對數生長的H929細胞,離心棄上清液,用無血清的RPMI-1640培養基重懸調整細胞數為1×105/mL,為防止其他細胞因子的干擾,加入終濃度為1.0 ng/mL的鼠抗人IL-6單克隆抗體,靜置4 h,以中和骨髓瘤細胞分泌的主要生長因子IL-6,棄上清液,PBS沖洗3次,按實驗分組處理細胞培養24 h后,收集各組的上清液,用MIP-1α、Scl ELISA試劑盒定量檢測各組細胞上清液中的MIP-1α、Scl水平,ELISA具體檢測方法按試劑盒說明書進行。
1.2.8各組OB活性定量檢測 OB按1×105/mL種于六孔板中培養2 d,PBS沖洗2次,根據實驗分組予以更換相應的培養液,培養3 d后棄上清液,用0.25%胰酶消化細胞,置于離心管,680×g離心10 min,去上清液,加TritonX-100裂解液1 mL,反復凍融3次(凍約30 min,融約3 min)取出樣品,按ALP試劑盒說明書配制測定管、標準管和空白管,每個管混勻后37 ℃水浴10 min,再加上顯色劑,立即混勻,用分光光度計在波長520 nm下進行比色,測定各組細胞ALP活性,并用Bradford法進行細胞蛋白標準化(按蛋白檢測試劑盒說明操作),最后用ALP/總蛋白的值計數ALP[單位:比活性(U/mg)]。
1.2.9Western blot檢測各組細胞中的ALP蛋白的表達 按1.2.3方法鑒定OB后,取對數生長期的第3代OB按1×105/mL加入6孔培養板,按實驗分組處理細胞繼續培養24 h后,按凱基全蛋白提取試劑盒說明書提取第3代OB的總蛋白,再按BCA蛋白定量試劑盒說明書定量后于-20 ℃保存。取30 μg總蛋白行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE),然后轉至聚偏氟乙烯(PVDF)膜,5%脫脂奶粉封閉1 h,加入抗ALP(1∶500)一抗、GAPDH(1∶600),冰箱4 ℃孵育過夜,室溫培育1 h,TBST溶液清洗3遍,每遍5 min,TBST洗膜后用ECL法顯影。條帶用Imαge J軟件分析,測定灰度值。目的條帶與對應的GAPDH條帶的灰度比值為目的蛋白相對表達水平。
1.10流式細胞儀(FCM)檢測OB凋亡率 將對數生長期的第3代OB置入10%小牛血清培養基中培養過夜,調節細胞密度至1×106/mL,按實驗分組再在含10%胎牛血清的RPMI-1640培養基中共同培育24 h后,收集細胞約5×106個進行如下操作:用Binding Buffer洗1次,室溫804.96×g離心10 min,收集細胞;洗滌后用100 μL Binding Buffer重懸細胞,加5 μL Annexin V-FITC,避光孵育20 min,再加10 μL碘化丙啶(PI,濃度為1 mg/mL)室溫下避光孵育15 min,Binding Buffer洗滌、重懸,立即上FCM檢測,每個樣品至少檢測5×106個細胞。利用Cell Quest功能軟件進行參數獲取和數據分析。每組樣品重復3次。
2 結 果
2.1OB鑒定及傳代 原代BMSCs培養5 d換液后,即可觀察到貼壁細胞,細胞形態短小,呈長梭型,細胞之間無突起連接,培養至第10天,細胞明顯增多、變長、變大,長梭型形態更加明顯,見圖1。培養至第14天時,細胞基本鋪滿皿底,細胞排列呈螺旋狀,長梭型。經茜素紅染色發現培養14 d后的OB能見到少部分鈣離子沉淀結節,ALP染色可發現OB細胞細胞質中有棕褐色顆粒或大片狀沉淀物體,顯示 ALP 染色陽性,符合OB的特征。
2.2H929細胞上清液中MIP-1α和Scl的水平 按實驗分組檢測各組H929細胞上清液中MIP-1α和Scl蛋白水平,結果顯示,MIP-1α組上清液中的MIP-1α和Scl的水平明顯高于對照組(t=6.42、3.56,P=0.000、0.012),MIP-1α-Ab組上清液中的MIP-1α、Scl水平明顯低于對照組(t=3.42、2.78P,P=0.001、0.012);而Scl及Scl-Ab組中的MIP-1α水平較對照組增減不明顯(t=0.618、0.744,P=0.552、0.474)、Scl水平較對照組增減明顯(t=6.97、3.17,P=0.000、0.013),見表1。
2.3各組OB的鈣離子沉積結節分析 運用茜素紅染色鈣離子沉積結節以檢測OB外基質成熟情況,按實驗分組干預后繼續培養至第6天,發現MIP-1α、Scl組較對照組鈣離子沉積結節減少(P<0.05)。MIP-1α-Ab、Scl-Ab組鈣離子沉積結節較對照組明顯增多(P<0.05)。但各組形態變化不顯著,見表2、圖2。
表1 各組細胞上清液中的MIP-1α、Scl水平
a:P<0.05,b:P<0.01,與對照組比較

A:對照組;B:MIP-1α組;C:Scl組;D:MIP-1α-Ab組;E:Scl-Ab組
圖2 不同組OB的鈣離子沉積結節(×40)
2.4各組OB的ALP活性 MIP-1α-Ab、Scl-Ab組的OB較對照組細胞生長快,細胞呈長梭形更明顯,有的呈纖維樣生長,細胞體積增大、而MIP-1α-Ab、Scl-Ab組的OB較對照組生長慢,細胞較短小,有的呈多角形,呈樹枝狀突起。經ALP 活性檢測試劑盒檢測后,ALP蛋白表達陽性細胞為細胞質和核均出現棕黑色顆粒。定量檢測發現MIP-1α-Ab、Scl-Ab組中OB的ALP的活性明顯高于對照組(均P<0.01)。而MIP-1α、Scl組中OB的ALP的活性明顯低于對照組(均P<0.01),見表2。
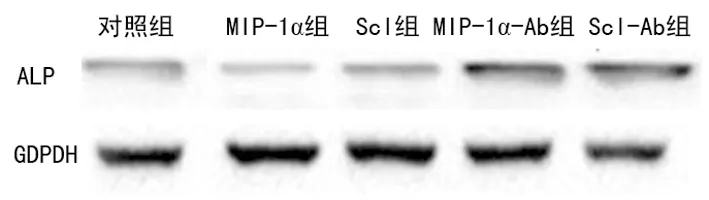
圖3 Western blot檢測各組OB的ALP蛋白質表達
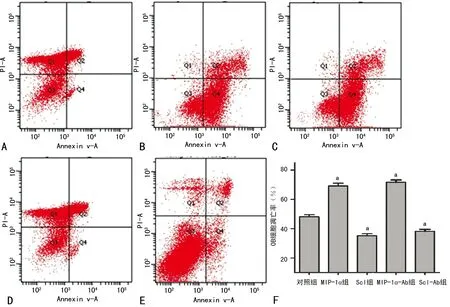
A:對照組;B:MIP-1α組:C Scl組;D:MIP-1α-Ab組;E:Scl-Ab組;F:凋亡率分析;a:P<0.05,與對照組比較
圖4 各組OB細胞的凋亡率
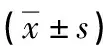
表2 不同組成骨細胞的鈣沉積結節
a:P<0.05,b:P<0.01,對照組比較
2.5各組OB的ALP蛋白表達水平 Western blot檢測結果進一步發現,培養至第6天發現MIP-1α、Scl組OB中ALP蛋白的灰度值較對照組表達減少,MIP-1α-Ab、Scl-Ab組OB ALP蛋白的灰度值較對照組表達明顯增多,見圖3。
2.6各組OB的凋亡率 FCM檢測顯示,MIP-1α、Scl組的OB凋亡率明顯高于對照組(t=2.19,P=0.02),MIP-1α-Ab、Scl-Ab組的OB凋亡率明顯低于對照組(t=3.12,P<0.01),見圖4。
3 討 論
MBD的發生、發展是多因子、多信號通路相互作用的結果,本團隊既往研究發現MIP-1α在MBD的發生、發展中有重要作用,并與Scl具有一定的關聯性。但它們之間到底如何影響并不清楚。體外研究發現高表達MIP-1α和Scl的MM細胞株H929,加入MIP-1α試劑后,H929細胞株表達更多的Scl。但當加入Scl后,細胞表達的MIP-1α并不增加,說明MIP-1α可能通過不同方式促進骨髓瘤細胞表達Scl,而Scl并不促進骨髓瘤細胞表達MIP-1α。反之運用同樣的方法,在高表達MIP-1α和Scl的H929細胞中加入MIP-1α-Ab中和MIP-1α后,細胞的MIP-1α和Scl水平均下降,而加入Scl-Ab中和Scl后,細胞的MIP-1α水平卻并不下降,這說明了抑制MIP-1α表達,也會影響骨髓瘤細胞表達Scl;而抑制骨髓瘤細胞表達Scl,則并不影響骨髓瘤細胞分泌MIP-1α。這也間接說明了骨髓瘤細胞表達Scl可能受MIP-1α調控。
MIP-1α如何促進骨髓瘤細胞分泌Scl,經檢索國內外文獻均未見明確報到。有研究報道將人骨髓瘤細胞系移植給鼠構建鼠骨髓瘤模型,發現其血漿中鼠源性Scl水平增高,說明Scl可能是由骨髓微環境產生的[9-10]。也可能是在MM患者中,骨髓瘤細胞與局灶的破骨祖細胞直接接觸、相互黏附后,通過多種途徑促進骨髓瘤細胞表達Scl[11]。也有可能骨髓瘤細胞通過多種途徑激活毗鄰的破骨祖細胞,局灶增多的破骨細胞(OC)會促進骨細胞分泌Scl增多。還有學者認為Scl是由破骨祖細胞產生的[12],而MIP-1α是一種非常重要的OC激活因子。因而,根據上述實驗結果,骨髓瘤細胞通過自分泌或旁分泌途徑分泌MIP-1α,當MIP-1α與受體結合后,使核因子κB(NF-κB)受體激動劑配體ligand of receptor activator of nuclear factor kappa B,RANKL)表達增高,增高的RANKL與骨髓巨噬細胞(bone marrow macrophages,BMM)膜上的RANK受體結合,再通過膜內部分特定區域激活下游信號通路,促進BMSCs分化破骨祖細胞增多,導致Scl增多。
Scl是SOST基因精細編碼的一種糖蛋白,是骨細胞分泌的重要信號分子,參與骨形成調節,決定骨重建過程中的骨量和結構。在MBD患者BMSCs分化OB的體外實驗中,當加入MIP-1α和Scl干預時,OB的凋亡率增加,鈣離子沉積及ALP活性均下降;而加入MIP-1α和Scl單抗干預時OB凋亡率則減少,鈣離子沉積及ALP的活性則增加,這說明MIP-1α和Scl可能抑制成骨祖細胞向OB分化、增殖,減弱細胞外基質的成熟及礦化作用,促進OB凋亡。結合上述實驗可推測MIP-1α抑制OB分化與成熟的機制可能是通過其促進骨髓瘤細胞分泌Scl來實現的。Scl是 Wnt/β-catenin信號通路抑制劑,Scl水平增高可直接減弱骨形態發生蛋白(BMP)的活性,抑制骨細胞的生成[13],Scl還可抑制Fra-1基因表達,抑制轉錄因子AP-1的活性,從而導致APL、骨鈣素(BGP)、Ⅰ型膠原和膠原酶等基因活性下降影響骨基質的形成[14];Scl可直接抑制Wnt/β-catenin及下游的信號通路,導致OB數量下降,骨量降低,細胞外基質成熟障礙、礦化降低。Scl還上調RANKL的表達,增加OC形成,抑制OB的分化成熟。本團隊在后續的實驗發現,在晚期MBD患者中,骨髓瘤細胞負荷較高時,骨髓瘤細胞分泌的Scl可使RhoA/ROCK1信號途徑失活。ROCK1蛋白質序列中含有Erk1/2結合結構域,當Scl抑制ROCK1時,就會暴露Erk1/2,從而啟動Bax凋亡通路,促進OC凋亡。從而導致MBD的發生、發展[15]。當加入MIP-1α和Scl單抗時,可能克服骨髓瘤細胞抑制Runx2表達,防止OB和骨細胞凋亡[16];加入單抗后,還可降低骨髓瘤細胞分泌RANKL,直接抑制破骨細胞前體分化,從而潛在地有助于OB的功能恢復[17]。這也間接地說明MIP-1α和Scl可導致MBD的發生發展。
綜上所述,MIP-1α可通過促進MM患者的骨髓瘤細胞分泌Scl,Scl通過多種途徑抑制MM患者BMSCs向OB的分化、成熟,促進其凋亡,參與MBD的發生發展,MIP-1α和Scl單抗可促進MBD患者OB的功能恢復和數量增加,促進MBD的改善,為MIP-1α和Scl單抗治療MBD奠定基礎,但其詳細的作用途徑及具體機制有待進一步研究。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
