一種可促進重組蛋白表達量和穩定性的多功能純化標簽的開發與利用
2019-04-23 03:10:24趙偉欣劉松劉立明陳堅堵國成
生物工程學報 2019年4期
關鍵詞:融合
趙偉欣,劉松,劉立明,陳堅,堵國成
?
一種可促進重組蛋白表達量和穩定性的多功能純化標簽的開發與利用
趙偉欣,劉松,劉立明,陳堅,堵國成
江南大學 生物工程學院,江蘇 無錫 214122

趙偉欣, 劉松, 劉立明, 等. 一種可促進重組蛋白表達量和穩定性的多功能純化標簽的開發與利用. 生物工程學報, 2019, 35(4): 626–635.Zhao WX, Liu S, Liu LM, et al. Development of a purification tag to produce thermostable fused protein. Chin J Biotech, 2019, 35(4): 626–635.
自組裝雙親短肽(Self-assembling amphipathic peptides,SAPs) 是一類親疏水氨基酸按一定規律分布、具有自聚合效應的短肽,融合在酶蛋白N端時,具有促進表達和穩定化的功能。根據前期研究結論,設計一條全新的基于SAPs (S1w,HNANARARHNANARARHNANARARHNARARAR) 的可促進融合蛋白表達和穩定性,并可用于鎳柱親和層析的多功能短肽標簽,在大腸桿菌表達系統中,將S1w以PT-linker (PTPPTTPTPPTTPTP)融合在堿性果膠酶(Alkaline polygalacturonate lyase,PGL)、脂肪氧合酶 (Lipoxygenase,LOX) 及綠色熒光蛋白(Green fluorescent protein,GFP) 的N末端時,與對應的野生型相比,PGL及LOX的粗酶活分別提高了3.1倍和1.89倍,GFP的熒光強度提高了16.22倍。S1w的3種融合酶均可用鎳柱進行親和純化,并具有較高的回收率。PGL及LOX在對應的熱處理條件下,與野生型相比,半衰期分別提高了2.16倍和3.2倍。將GFP-S1w在枯草芽孢桿菌及畢赤酵母表達系統中表達,發現在枯草芽孢桿菌中融合蛋白表達量提高明顯,但在畢赤酵母中表達量幾乎沒有改變。說明在原核表達體系中,S1w可作為一種新型的促表達、穩定化及可純化的多功能標簽。
自組裝雙親短肽,融合蛋白,表達量,熱穩定性,純化,表達宿主
微生物發酵已逐漸應用于多種酶蛋白的工業化生產過程中。目前,如何獲得高產量、高穩定性及高催化效率的多功能酶蛋白是酶催化工業應用的主要瓶頸問題[1]。雖然通過基因工程和蛋白質工程等手段可以對酶蛋白本身進行改造,利用合適的表達宿主或優化培養條件可使得微生物對酶進行高效異源表達,但實際生產中還是經常面臨著蛋白低可溶表達、穩定性差、催化效率低和難分離純化等諸多問題[2]。利用分子生物學技術可以將不同酶分子融合表達成為符合工業需求的多功能酶,可應用于多功能催化體系的研究中并顯示出重要的理論和應用研究價值[3]。此外,末端融合功能肽的方法對酶蛋白的表達量及催化性質的改造已取得廣泛顯著成果[4-6]。在無需酶蛋白的晶體結構信息的條件下進行末端融合,可有效促進其異源表達產量及相關酶學性質[7]。
由親疏水氨基酸按照一定規律分布組成的自組裝雙親短肽(Self-assembling amphipathic peptides,SAPs) 被Lu等首次應用于脂肪氧合酶(Lipoxygenase,LOX,EC 1.13.11.12) 的熱穩定性及催化效率改造[5]。其中,來源于酵母Zution蛋白中的SAP (S1,AEAEAKAKAEAEAKAK) 對環糊精轉移酶[8]、腈水合酶[9]、堿性淀粉酶[10]及堿性果膠酶[11]的蛋白表達量、熱穩定性及催化效率均有一定促進作用。基于此,具有與S1類似氨基酸組成和排布方式的SAP (S11,AEAEAHAHAE AEAHAH)被開發并應用在多個酶蛋白的異源表達過程中,其融合蛋白在大腸桿菌中的表達量、穩定性及催化效率均有所提高,且S11內的組氨酸可用于融合酶的鎳柱親和層析,其純化效率較His-tag純化效率明顯提高[12]。S1與S11之間的4個氨基酸殘基的差異,說明氨基酸組成對SAPs融合酶表達量有著重要影響。此外,氨基酸的突變會引入相應密碼子的改變,諸多研究證明密碼子對異源基因轉錄和翻譯過程有重要影響[13-14]。故根據前期研究結論,對SAPs合理的改造和修飾可以進一步提高其多功能應用效果。
本研究提出一條全新的多功能SAP(S1w,HNANARARHNANARARHNANARARHNARARAR),其可促進重組融合蛋白表達和穩定性,同時可作為一種用于鎳柱親和層析的多功能蛋白標簽。并將其應用到堿性果膠酶(Alkaline polygalacturonate lyase,PGL)、脂肪氧合酶(Lipoxygenase,LOX) 及綠色熒光蛋白(Green fluorescent protein,GFP) 中驗證其作用效果。此外以GFP為模式蛋白,研究S1w在不同表達體系(大腸桿菌、枯草芽孢桿菌及畢赤酵母) 中的促表達功效。
1 材料與方法
1.1 材料
1.1.1 菌株及質粒
大腸桿菌JM109作為質粒的克隆宿主。酶/蛋白表達宿主.BL21(DE3)、枯草芽孢桿菌W600及畢赤酵母GS115由本實驗室保存。
含有PGL表達基因的質粒pET-22b(+)/、含有LOX表達基因的pET-22b(+)/質粒及含有GFP表達基因的pET-22b(+)/質粒由本實驗室前期構建并保存(圖1A)[12]。S1的PGL融合酶(PGL-S1)表達質粒pET-22b(+)/-及S11的PGL融合酶(PGL-S11) 表達質粒pET-22b (+)/-由本實驗室保存。pPIC9K質粒及pP43NMK質粒購于TaKaRa (大連) 公司。
1.1.2 試劑
Prime STAR (HS) DNA高保真聚合酶、DNA片段回收試劑盒、質粒提取試劑盒、限制性內切酶及大腸桿菌感受態制備試劑盒均購于TaKaRa (大連) 公司。異丙基-β-D硫代半乳糖苷(IPTG)、氨芐青霉素、卡那霉素及遺傳霉素G418均購于生工生物工程(上海) 股份有限公司。NuPAGE?Novex?Bis-Tris預制凝膠及所用標準蛋白marker購自Life Technologies公司。一步克隆反應ClonExpressTMⅡ試劑盒購于南京諾唯贊公司。DNA片段合成、引物合成及測序由生工生物工程(上海) 股份有限公司完成。所用相關底物等均購于Sigma公司,其他常規試劑均為國產或進口分析純。
1.2 方法
1.2.1 S1w融合酶突變體的構建
自組裝雙親短肽S1w (HNANARARHNANA RARHNANARARHNARARAR)連同PT-linker (PTPPTTPTPPTTPTP)的編碼基因由生工生物工程(上海) 股份有限公司合成并插入至pET-22b(+)/、pET-22b(+)/、pET-22b(+)/的Ⅰ和Ⅰ酶切位點之間,分別獲得PGL-S1w、LOX-S1w和GFP-S1w融合酶的表達質粒pET-22b(+)/、pET-22b(+)/和pET-22b(+)/(圖1B)。并將pET-22b(+)/質粒作為模板,以表1中所列對應引物進行聚合酶鏈式反應(PCR)獲得融合酶的表達基因片段,并用一步克隆酶融合至線性化的質粒pP43NMK及pPIC9K中,獲得在枯草芽孢桿菌及畢赤酵母中的pP43NMK/和pPIC9K/表達質粒(圖1B)。并用引物對DS-pP43-F/ DS-pP43-R和DS-9K-F/DS-9K-R分別將S1w和PT-linker表達基因從pP43NMK/和pPIC9K/中缺失,獲得在枯草芽孢桿菌及畢赤酵母中表達的野生型GFP (圖1A)。

表1 本文所用引物序列表

圖1 重組質粒構建
1.2.2 大腸桿菌發酵培養條件
種子培養:挑取單菌落接入種子培養基[7]裝液量為25 mL的三角瓶(250 mL) 中,培養溫度37 ℃,搖床轉速200 r/min,培養12 h。
發酵培養:按3% (/) 的接種量接入裝液量為25 mL的三角瓶(250 mL) 的發酵培養基中,培養溫度37 ℃,當600達到0.6時加入IPTG誘導,并同時調整溫度到該酶最適宜的誘導溫度下培養。
培養條件:PGL:IPTG 0.04 mmol/L,30 ℃下培養48 h;LOX:IPTG 1 mmol/L,20 ℃下培養48 h;GFP:IPTG 0.05 mmol/L,25 ℃下培養24 h。
1.2.3 S1w的GFP融合酶在不同表達宿主中的培養條件
大腸桿菌培養參照文獻[7]完成。枯草芽孢桿菌培養參照文獻[15]完成。畢赤酵母菌培養參照文獻[16]完成。
1.2.4 酶活及熒光檢測方法
堿性果膠酶(PGL)[17]的酶活測定方法參照前期研究。
由于脂肪氧合酶(LOX) 為胞內表達,故取一定的菌體,用20 mmol/L磷酸鹽緩沖液(pH 7.5)清洗2次,用相同體積的緩沖液懸浮(使得600=5),破壁后,離心取上清液,按照前期研究方法測定LOX酶活[18]。
綠色熒光蛋白(GFP) 檢測方法:在避光條件下將熒光樣品進行一定的稀釋,采用多功能酶標儀(BioTek,Winooski,VT,USA) 進行熒光強度檢測,緩沖液A (20 mmol/L磷酸鹽緩沖液,pH 7.5) 作為空白對照。
1.2.5 酶/蛋白的分離純化
A液:20 mmol/L磷酸鹽緩沖液,500 mmol/L NaCl,20 mmol/L咪唑。
B液:20 mmol/L磷酸鹽緩沖液,500 mmol/L NaCl,500 mmol/L咪唑。
20 mmol/L的pH 7.5的磷酸鹽緩沖液配 制方法:190 mL 20 mmol/L NaH2PO4與810 mL 20 mmol/L Na2HPO4均勻混合。
野生型PGL和PGL-S1[17]、野生型LOX[18]的純化分別參照前期研究方法進行操作。野生型GFP采用鹽析方法進行初步富集和陰離子親和層析進行純化,操作過程參照前期報道[19]。
PGL-S1w、PGL-S11、LOX-S1w及GFP-S1w融合酶采用鎳柱親和層析。含有目的蛋白的胞外發酵液或胞內上清液經過0.22 μm濾膜過濾后作為待純化樣品。用緩沖液A以1 mL/min的流速平衡HisTrap 5 mL FF純化柱。將含有重組融合酶樣品以1 mL/min流速上樣,并用緩沖液A洗脫未結合蛋白,平衡純化柱。將結合在鎳柱上的蛋白分別用8%的緩沖液B、60%的緩沖液B及100%的緩沖液B進行梯度洗脫并收集含有目的蛋白的組分。
所獲得的目的蛋白溶液使用5 mL HiTrap Desalting純化柱用緩沖液A以5 mL/min的流速進行脫鹽處理,所得蛋白樣品于4 ℃保存。
1.2.6 蛋白電泳分析及蛋白濃度測定
SDS-PAGE分析,使用Life Technologies公司預制膠NuPAGE?Novex?Bis-Tris,操作步驟詳見說明書。以0.1% (/) 考馬斯亮藍R-250進行染色。
采用Bradford方法測定蛋白質濃度,以牛血清蛋白作為標準蛋白測定蛋白濃度標準曲線。具體操作參照試劑盒說明書。
1.2.7 酶的熱穩定性測定方法
重組PGL及其突變體的熱穩定性以60 ℃下的半衰期(1/2,min) 來表示。將純化并除鹽后的PGL用緩沖液A稀釋至一定濃度,在60 ℃下保溫,每隔3 min測定殘余酶活。
脂肪氧合酶熱穩定性測定方法:將純化后的酶用緩沖液A稀釋到蛋白濃度為100 μg/mL并在50 ℃保溫,間隔測定殘余酶活,計算1/2。1/2按照文獻所述方式進行擬合計算[20]。
2 結果與分析
2.1 多功能肽S1vw序列的設計
S1類SAPs融合在酶蛋白N端時具有促進融合酶表達和穩定化的功能[12]。前期研究結果表明[21],蛋白質翻譯過程中,新生肽鏈N端的正電荷氨基酸殘基能與帶負電的核糖體相結合,從而減緩蛋白質翻譯速率,促進蛋白質表達產量。為強化S1類SAPs的促表達功能,提出S1w短肽,在其中引入不帶電的天冬酰胺殘基(N) 和帶正電荷的精氨酸殘基(R),使其整體呈現凈電荷為正。常用的組氨酸標簽His-tag (HHHHHH) 含有6個組氨酸,關于S11的研究結果顯示[12],4個組氨酸殘基即可實現良好的純化效果,且組氨酸殘基在蛋白質分子中均勻分布,可防止其被包埋[22]。故在S1w內部均勻插入4個組氨酸(H),最終確定S1w的氨基酸序列為HNANARARHNANARA RHNANARARHNARARAR,以期融合在酶蛋白N端時,既可促進融合酶的表達量和穩定性,又可作為鎳柱純化的多功能蛋白標簽。
2.2 S1vw融合酶表達質粒的構建
S1w (HNANARARHNANARARHNANARA RHNARARAR) 是根據S1及S11序列優化而來。為研究其對不同酶蛋白的表達量、穩定性等的影響,本研究構建PGL、LOX及GFP的S1w融合酶突變體(圖1),在大腸桿菌中進行異源表達。并將GFP-S1w在大腸桿菌、枯草芽孢桿菌及畢赤酵母中進行表達,比較其表達量差異。研究S1w針對不同酶/蛋白和S1w在不同表達系統的普適性應用效果。
2.3 S1vw對不同酶/蛋白表達量的影響
在大腸桿菌表達系統中,PGL胞外表達,而LOX和GFP胞內表達。分別測定PGL的胞外酶活、LOX的胞內酶活和GFP表達菌的全細胞熒光強度。結果如圖2所示,分別在3種酶/蛋白的N端融合S1w后,融合酶/蛋白的表達量明顯提高。PGL的融合酶(PGL-S1w) 的胞外酶活提高至原野生型PGL的3.1倍,與PGL-S1和PGL-S11相比,分別提高了1.2倍和0.95倍,說明相較于S1和S11,S1w促表達效果更佳。LOX的融合酶(LOX-S1w) 的胞內酶活較野生型提高了1.89倍。在相同處理條件下,GFP-S1w的熒光強度(熒光強度/菌濃600) 是GFP的16.22倍。SDS-PAGE分析也表明融合SAPs后條帶變粗,且蛋白分子量也發生一定變化,說明N端融合S1w后,酶/蛋白表達量顯著提高。
2.4 S1vw對不同酶蛋白純化效率的影響
野生型PGL、LOX及GFP不能直接用鎳柱親和層析進行純化,由于野生型表達過程中蛋白含量較低,往往需要進行一步硫酸銨或乙醇富集的工序,導致整個過程中蛋白質回收率較低。表2詳細比較了野生型PGL、LOX及GFP與所對應的S1w融合酶在不同純化條件下的回收率。結果顯示,在不經過任何預處理的條件下,S1w的3種融合酶的回收率分別達到55.43%、34.22%和64.35%。而所對應的野生型PGL、LOX及GFP,在相同條件下幾乎不能被有效純化,經過硫酸銨和乙醇沉淀富集后用對應的方法進行純化[17-19],回收率分別為20.45%、26.22%和36.54%。
此外,前期研究表明S11短肽也具有純化功能[12],如表2所示,相同條件下PGL-S11與PGL-S1w的回收率相近,說明S1w保持了S11的純化功能。純化后蛋白電泳分析如圖3所示。且需要說明的是,前期研究已經證明PGL和LOX的His-tag融合酶在相同條件下不能用鎳柱純化,GFP的His-tag融合酶在與GFP-S1w相同的純化條件下回收率僅有8%[12]。
2.5 S1vw對不同酶蛋白穩定性的影響
為比較S1w對不同酶的穩定性及催化活性的影響,測定純化后的野生型PGL、LOX及融合酶的比酶活及半衰期。結果顯示(圖4和圖5),PGL-S1w在60 ℃下的半衰期由5.2 min提高至16.43 min,比酶活279.14 U/mg提高至758.9 U/mg。LOX-S1w在50 ℃下半衰期由9.4 min提高至29.78 min,比酶活由32.5 U/mg提高至82.3 U/mg。此外,與PGL-S1和PGL-S11相比[12],S1w進一步促進了PGL的熱穩定性和催化活性 (圖4)。
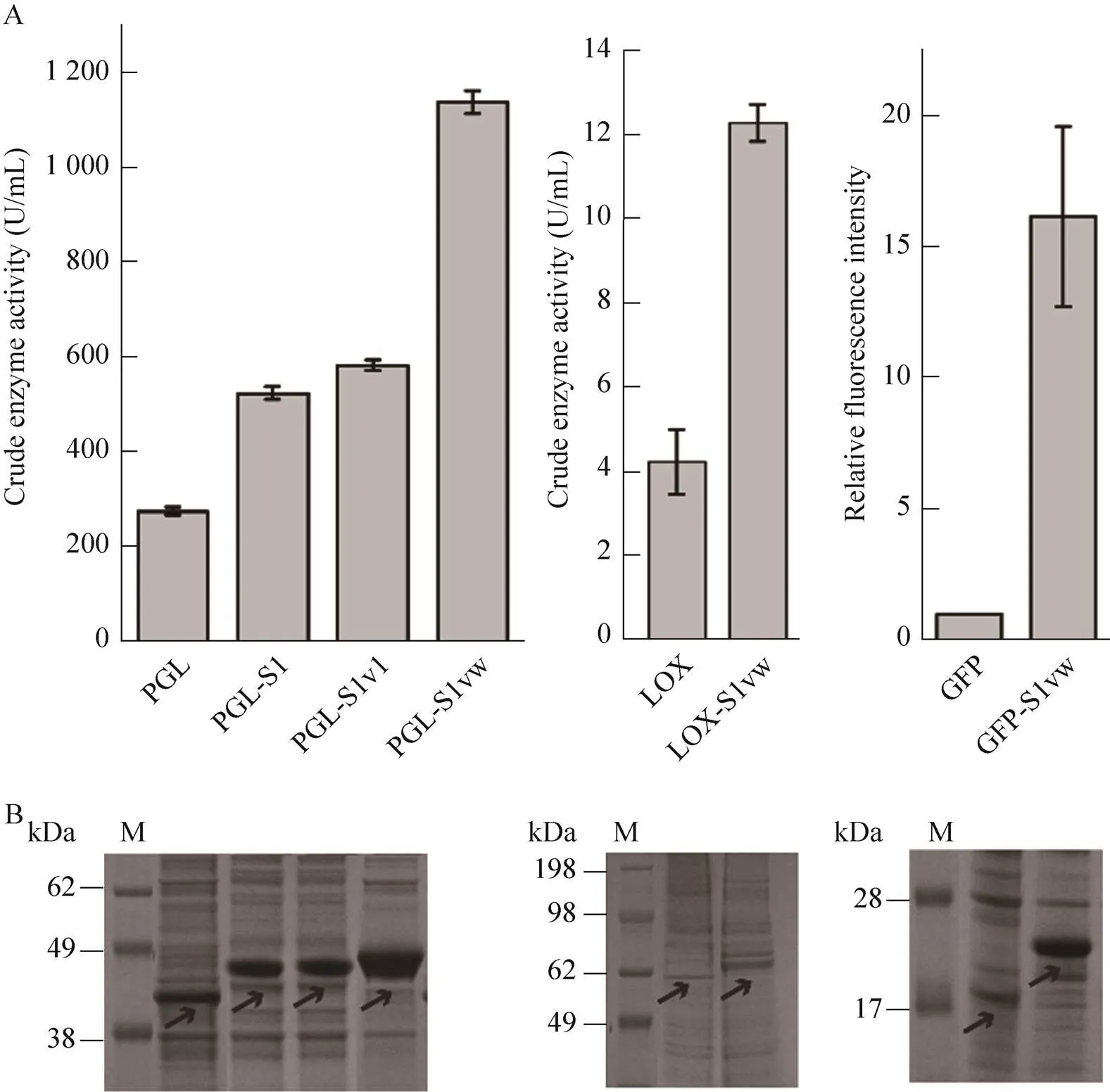
圖2 酶/蛋白及其對應的SAPs融合酶表達量分析
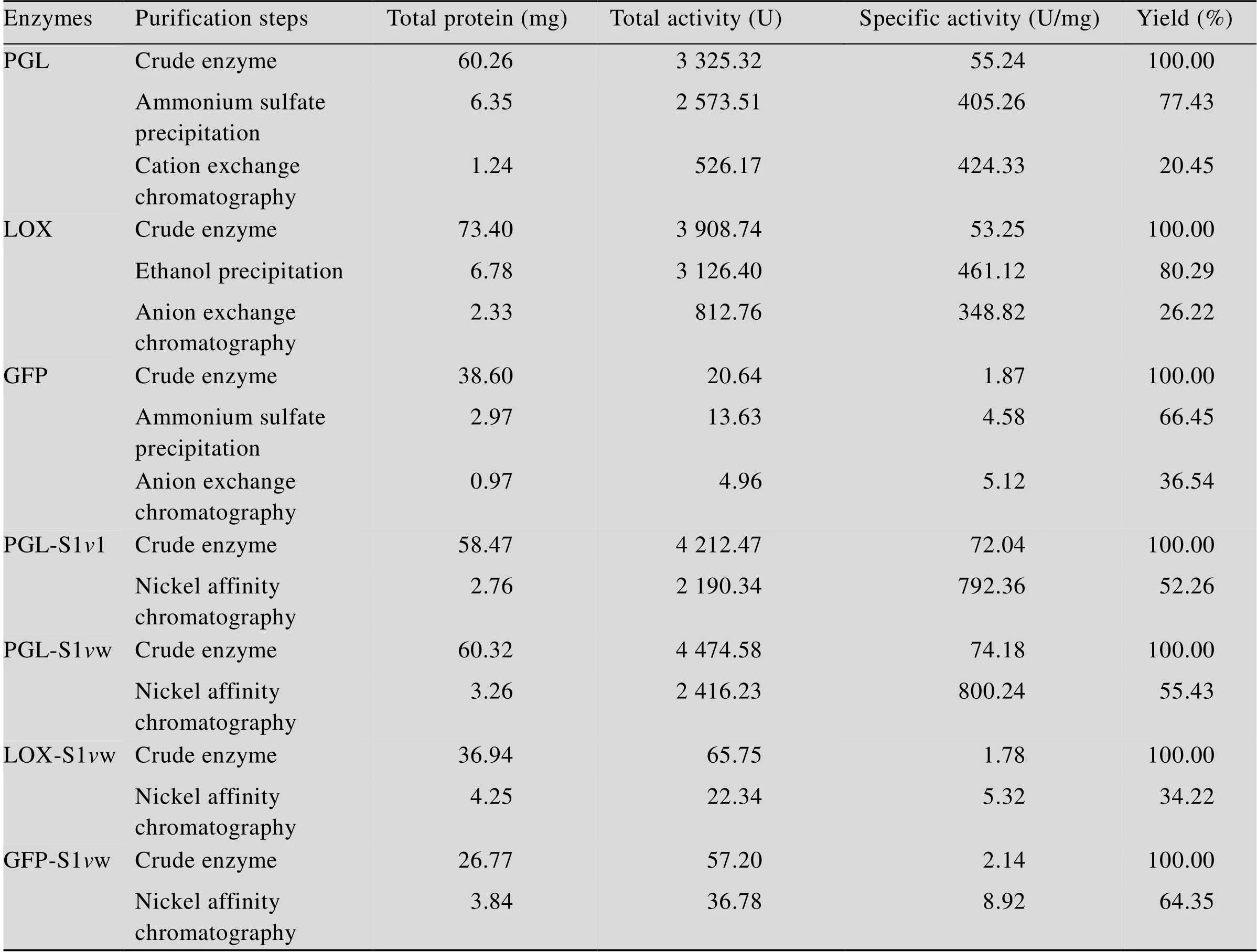
表2 酶/蛋白純化收率表

圖3 酶/蛋白純化后的SDS-PAGE圖譜

圖4 PGL及其SAPs融合酶的半衰期與比酶活測定
2.6 S1vw在不同表達宿主中對融合酶表達量的影響
由于不同表達系統中異源蛋白的表達機制存在差異,故本研究以GFP為模式蛋白,比較S1w在不同宿主中(大腸桿菌、枯草芽孢桿菌和畢赤酵母)對GFP的表達量的影響。如圖6所示,在大腸桿菌和枯草芽孢桿菌中,融合S1w后熒光強度顯著提高,但在畢赤酵母表達系統中,GFP和GFP-S1w的熒光強度相近,說明S1w在原核表達體系中對其融合酶的表達量有促進作用,但對真核表達系統酶/蛋白異源表達的產量沒有明顯影響。

圖5 LOX及其SAPs融合酶的半衰期與比酶活測定
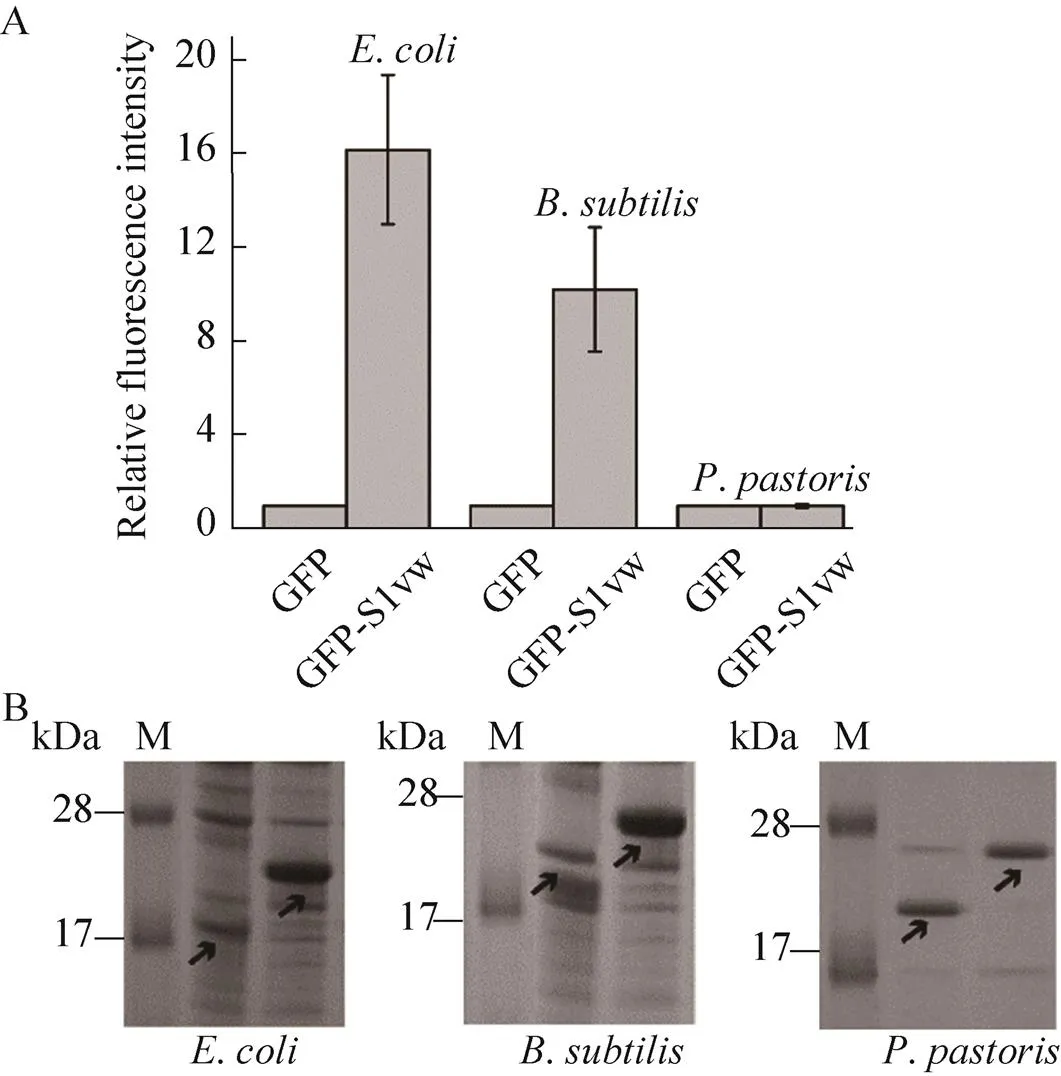
圖6 GFP和GFP-S1vw在不同表達宿主中的表達量
3 討論
酶的高表達、高穩定性、高效純化工藝和循環利用技術是目前酶工程領域的主要研究方向。融合功能標簽技術作為一種便捷、高效的酶蛋白改造策略也已經在該領域取得一定實效成果[23]。本研究在SAPs的前期研究的基礎上[7, 12, 24],提出一種可促進在原核表達宿主中酶蛋白表達產量、提高融合酶穩定性并可用于鎳柱純化的多功能標簽S1w。分別將S1w以PT-linker融合在PGL、LOX和GFP的N端,在大腸桿菌中培養發酵,與對應的野生型相比,粗酶活或熒光強度分別提高了3.1、1.89和16.22倍,且用鎳柱進行親和層析純化達到了可觀的回收率。PGL和LOX的融合酶在對應熱處理條件下的半衰期分別提高了2.16倍和3.2倍。將GFP-S1w在枯草芽孢桿菌及畢赤酵母表達系統中表達,發現在枯草芽孢桿菌中融合酶表達量提高明顯,但在畢赤酵母中表達量幾乎沒有改變。說明在原核表達體系中S1w可作為一種新型的促表達、穩定化和純化的多功能標簽。
S1類SAPs融合在酶蛋白N端時具有促進融合酶表達和穩定化的功能。為優化S1類SAPs的促表達特性,在S1w中引入不帶電的天冬酰胺殘基和帶正電荷的精氨酸殘基,并插入正電荷的組氨酸以便用于鎳柱純化,結果表明,優化之后的S1w促表達效果更加明顯,這很有可能是S1w整體呈現凈電荷為正的結果。但由于真核原核表達系統的差異,S1w在真核畢赤酵母中沒有促表達的功能,這也為后續SAPs的促表達研究提供一定的研究方向。
盡管S1w整體帶電情況發生改變,但其親疏水性交替分布的形式仍然與S1和S11類似,并且保持了S1類SAPs的穩定化效果。SAPs內的疏水氨基酸殘基對其融合酶形態至關重要,強疏水性的SAPs可直接導致活性包涵體的產生[24]。相比于S1和S11,S1w引入更多的疏水丙氨酸殘基,故其穩定化功效進一步提高。
生物活性酶在工業生產中具有廣泛的應用,酶的分離純化是工業酶成本計算的重點考量指標之一。在所有酶的分離純化方法中[25],親和層析是相對簡單和節約成本的方法,其中最常用的就是組氨酸標簽親和層析方法。在S1w中引入4個組氨酸,且均勻分布在S1w中,可提高組氨酸殘基與鎳柱的結合效率。此外,Slw融合酶的熱穩定性較高,其在純化過程中活性損失較小,有利于提高目的蛋白的回收率[26]。
多功能標簽的融合可以在融合單一蛋白或短肽的同時實現多個酶/蛋白性質的優化,另一個明顯的優勢是可以減少由外來融合標簽帶來的空間組織(結構域)之間的相互影響,保持酶/蛋白原有催化或功能特性。因此,基于SAPs的多功能肽的開發將為酶或功能蛋白的改造研究提供新的技術手段和研究方向。
[1] Yu K, Liu CC, Kim BG, et al. Synthetic fusion protein design and applications. Biotechnol Adv, 2015, 33(1): 155–164.
[2] Kaur J, Kumar A, Kaur J. Strategies for optimization of heterologous protein expression in: roadblocks and reinforcements. Int J Biol Macromol, 2018, 106: 803–822.
[3] Zhang Q, Wu XY, Jiang XK, et al. Trend of hybrid enzyme design in the big data era. Chin J Biotech, 2018, 34(7): 1033–1045 (in Chinese). 張群, 吳秀蕓, 蔣緒愷, 等. 大數據時代雜合酶的設計及其新趨勢. 生物工程學報, 2018, 34(7): 1033–1045.
[4] Takano K, Okamoto T, Okada J, et al. Stabilization by fusion to the C-terminus of hyperthermophileRNase HI: a possibility of protein stabilization tag. PLoS ONE, 2011, 6(1): e16226.
[5] Lu XY, Liu S, Zhang DX, et al. Enhanced thermal stability and specific activity oflipoxygenase by fusing with self-assembling amphipathic peptides. Appl Microbiol Biotechnol, 2013, 97(21): 9419–9427.
[6] Zhao Q, Xu WH, Xing L, et al. Recombinant production of medium-to large-sized peptides inusing a cleavable self-aggregating tag. Microb Cell Fact, 2016, 15: 136.
[7] Zhao WX, Liu S, Liu LM, et al. Analysis of the factors influencing the expression of enzymes fused with self-assembling amphipathic peptides. Food Ferment Ind, 2017, 43(12): 1–6 (in Chinese). 趙偉欣, 劉松, 劉立明, 等. 自組裝雙親短肽氨基酸組成及連接肽對其融合酶表達量的影響. 食品與發酵工業, 2017, 43(12): 1–6.
[8] Han RZ, Li JH, Shin HD, et al. Fusion of self-assembling amphipathic oligopeptides with cyclodextrin glycosyltransferase improves 2--D-glucopyranosyl-L-ascorbic acid synthesis with soluble starch as the glycosyl donor. Appl Environ Microbiol, 2014, 80(15): 4717–4724.
[9] Liu Y, Cui WJ, Liu ZM, et al. Enhancement of thermo-stability and product tolerance ofnitrile hydratase by fusing with self-assembling peptide. J Biosci Bioeng, 2014, 118(3): 249–252.
[10] Yang HQ, Lu XY, Liu L, et al. Fusion of an oligopeptide to the N terminus of an alkaline α-amylase fromsimultaneously improves the enzyme’s catalytic efficiency, thermal stability, and resistance to oxidation. Appl Environ Microbiol, 2013, 79(9): 3049–3058.
[11] Liu S, Wang MX, Du GC, et al. Improvement of thermal stability of alkaline polygalacturonate lyase by fusing with self-assembling amphipathic peptides. Food Ferment Ind, 2015, 41(11): 1–6 (in Chinese).劉松, 汪明星, 堵國成, 等. 融合自組裝雙親短肽提高堿性果膠酶熱穩定性. 食品與發酵工業, 2015, 41(11): 1–6.
[12] Zhao WX, Liu LM, Du GC, et al. A multifunctional tag with the ability to benefit the expression, purification, thermostability and activity of recombinant proteins. J Biotechnol, 2018, 283: 1–10.
[13] Goodman DB, Church GM, Kosuri S. Causes and effects of N-Terminal codon bias in bacterial genes. Science, 2013, 342(6157): 475–479.
[14] Zhong C, Wei P, Zhang YHP. Enhancing functional expression of codon-optimized heterologous enzymes inBL21(DE3) by selective introduction of synonymous rare codons. Biotechnol Bioeng, 2017, 114(5): 1054–1064.
[15] Chen X. Study on the expression and fermentation optimization of L-asparaginase inWB600[D]. Wuxi: Jiangnan University, 2015 (in Chinese).陳璇. L-天冬酰胺酶在WB600中的表達與發酵過程優化研究[D]. 無錫: 江南大學, 2015.
[16] Ren CH, Zhang J, Du GC, et al. Enhancing thermal stability of glucose oxidase by fusing amphiphilic short peptide. Chin J Biotech, 2018, 34(7): 1106–1116 (in Chinese). 任春慧, 張娟, 堵國成, 等. 基于融合雙親短肽提高葡萄糖氧化酶的熱穩定性. 生物工程學報, 2018, 34(7): 1106–1116.
[17] Wang MX, Liu S, Liu L, et al. Fusions of amphipathic peptide to the alkaline polygalacturonate lyase fromsp. WSHB04-02 improves the production. J Food Sci Biotechnol, 2016, 35(5): 504–509 (in Chinese).汪明星, 劉松, 劉龍, 等. 融合短肽促進堿性果膠酶的高效表達. 食品與生物技術學報, 2016, 35(5): 504–509.
[18] Qiu FF. Study on the high-level expression and thermal stability oflipoxygenase[D]. Wuxi: Jiangnan University, 2017 (in Chinese).邱芳芳. 重組脂肪氧合酶的高效表達和熱穩定性研究[D]. 無錫: 江南大學, 2017.
[19] Lu XY, Zhang J, Liu S, et al. Overproduction, purification, and characterization of extracellular lipoxygenase ofin. Appl Microbiol Biotechnol, 2013, 97(13): 5793-5800.
[20] O'Fágáin C. Enzyme stabilization-recent experimental progress. Enzyme Microb Technol, 2003, 33(2/3): 137–149.
[21] Charneski CA, Hurst LD. Positively charged residues are the major determinants of ribosomal velocity. PLoS Biol, 2013, 11(3): e1001508.
[22] Mei YZ, Chen YR, Zhai RY, et al. Cloning, purification and biochemical properties of a thermostable pectinase fromM29. J Mol Catal B: Enzymat, 2013, 94: 77–81.
[23] Kosobokova EN, Skrypnik KA, Kosorukov VS. Overview of fusion tags for recombinant proteins. Biochemistry (Moscow), 2016, 81(3): 187–200.
[24] Lin ZL, Zhou BH, Wu W, et al. Self-assembling amphipathic alpha-helical peptides induce the formation of active protein aggregates. Faraday Dis, 2013, 166: 243–256.
[25] Becker K, Van Alstine J, Bülow L. Multipurpose peptide tags for protein isolation. J Chromatogr A, 2008, 1202(1): 40–46.
[26] Farchaus JW, Ribot WJ, Jendrek S, et al. Fermentation, purification, and characterization of protective antigen from a recombinant, avirulent strain of. Appl Environ Microbiol, 1998, 64(3): 982–991.
Development of a purification tag to produce thermostable fused protein
Weixin Zhao, Song Liu, Liming Liu, Jian Chen, and Guocheng Du
School of Biotechnology, Jiangnan University, Wuxi 214122, Jiangsu, China
Self-assembling amphipathic peptides (SAPs) have alternating hydrophilic and hydrophobic residues and can affect the thermal stabilities and catalytic properties of the fused enzymes. In this study, a novel multifunctional tag, S1w (HNANARARHNANARARHNANARARHNARARAR) was developed to modify fused enzymes. After fusing S1w at the enzymes/proteins N-terminus through a PT-linker, the crude enzymatic activities of polygalacturonate lyase and lipoxygenase were enhanced 3.1- and 1.89-fold, respectively, compared to the wild-type proteins. The relative fluorescence intensity of the green fluorescent protein was enhanced 16.22-fold. All the three S1w fusions could be purified by nickel column with high purities and acceptable recovery rates. Moreover, S1w also induced the thermostabilities enhancement of the fusions, with polygalacturonate lyase and lipoxygenase fusions exhibiting 2.16- and 3.2-fold increase compared with the corresponding wild-type, respectively. In addition, S1w could enhance the production yield of green fluorescent protein inandwhile the production of GFP and its S1w fusion changed slightly in. These results indicated that S1w could be used as a multifunctional tag to benefit the production, thermal stability and purification of the fusion protein in prokaryotic expression system.
self-assembling amphipathic peptides, fusion enzymes,production yield, thermal stability, purification, expression host
10.13345/j.cjb.180363
September 9, 2018;
November 5, 2018
National Natural Science Foundation of China (No. 31401638).
Song Liu. Tel: +86-510-85918307; Fax: +86-510-85918309; E-mail: liusong@jiangnan.edu.cn
Liming Liu. Tel/Fax: +86-510-85197875; E-mail: mingll@jiangnan.edu.cn
國家自然科學基金 (No. 31401638) 資助。
(本文責編 陳宏宇)
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
