轉BpTCP7白樺耐鹽堿能力分析
2019-04-10 03:11:58李芳蕊許思佳安琳君李慧玉
福建林業科技 2019年1期
關鍵詞:植物
張 園,李芳蕊,李 洋,許思佳,安琳君,李慧玉
(東北林業大學,黑龍江 哈爾濱 150040)
我國鹽漬化和次生鹽漬化土地有4000萬hm2以上,占現有耕地的四分之一,而且鹽漬化日趨嚴重,影響到我國農林業的生產。目前,鹽堿地綜合治理主要依靠生物措施,發掘更多的耐鹽植物是當前解決鹽漬化的重要手段。植物耐鹽機制研究的深入及基因工程育種的發展,為培育耐鹽植物特別是林木品種提供了新途徑。
TCP是植物特有的一類轉錄因子。TCP基因家族具60個氨基酸殘基組成的堿性區-螺旋-環-螺旋(bHLH)的TCP保守結構域(TCP domain)[1]。根據其TCP結構域不同分為2個亞家族,ClassⅠ(TCP-P)和ClassⅡ(TCP-C),BpTCP7屬于ClassⅡ亞類[2]。研究表明,TCP在植物中參與多種植物激素的應答反應,通過偶聯這些激素響應過程來調控植物生長發育過程,如分枝、株高、葉形、花形、種子萌發、維管束生長、配子體發育、光形態建成以及晝夜節律調控。另外,在植物低溫、高鹽等逆境脅迫應答反應中發揮一定的作用[3-10]。
白樺(Betulaplatyphylla)是我國東北地區蓄積量最大的鄉土速生闊葉樹種,也是天然次生林更替的先鋒樹種。前期,通過基因工程育種手段獲得了轉BpTCP7基因過表達白樺植株,本研究以轉TCP7白樺為材料,檢測NaHCO3脅迫后的各轉基因株系的生長量、保護酶類、滲透調劑物及MDA含量等指標,分析其耐鹽堿能力,并篩選抗鹽堿優良的株系,為利用植物改良鹽堿地提供種質資源。
1 材料與方法
1.1 植物材料
2年生轉BpTCP7基因過表達白樺株系4個(分別為TCP7-1,TCP7-2,TCP7-3,TCP7-4)及非轉基因對照株系(WT)。通過組培擴繁,每個株系擴繁50株。選擇規格相同的塑料花盆,裝入等量培養土,將苗木移入盆中,第2年夏,在植物生長旺盛時,株系內選擇生長基本一致的植株用于NaHCO3脅迫處理試驗。
1.2 試驗方法
1.2.1 處理方法 分別選取每個株系各36株,分成3組,每組各12株,每盆1株,分別向盆內澆入1、2、4 g·L-13種不同質量濃度的NaHCO3溶液,以澆入等量清水為對照。每天澆施500 mL·株-1溶液,處理時間為12 d。分別在處理0、3、6、9、12 d時,取植株第3~5片功能葉,每個株系每個處理各采樣20片左右,在液氮條件下充分混合研磨成粉末,進行各指標測定。脅迫處理12 d后,對試驗苗木復水。
1.2.2 指標測定 超氧化物歧化酶(SOD) 活性采用NBT比色法[11],游離脯氨酸含量采用磺基水楊酸浸提-酸性茚三酮顯色法[12],丙二醛(MDA)含量采用TBA 比色法[13]。
生長量的測定:鹽處理前和復水20 d分別用塔尺測定主干高度,計算高生長量和相對生長量。高生長量= 處理結束后樹高-處理前樹高;相對生長量=(處理高生長量/對照生長量)×100%。
1.2.3 數據分析與處理 數據統計分析和作圖由Excel和SPSS 16.0軟件系統完成。
2 結果與分析
2.1 NaHCO3脅迫對細胞保護酶活性的影響
SOD是氧自由基代謝的關鍵酶類,可以清除植物體內的自由基,在逆境條件下其活性高低可反映機體的抗逆能力[14]。從圖1A可以看出,在0.2%NaHCO3脅迫3 d后,轉基因及對照株系的SOD活性變化不明顯;6~9 d時,除了TCP7-4外,其他各株系SOD活性逐漸上升;9 d時各株系的SOD活性最高,尤其以TCP7-1最為顯著;隨著脅迫時間進一步延長,各株系SOD活性下降。總體上看,在整個脅迫過程中,轉基因及對照株系的SOD活性差異不顯著。在0.4% NaHCO3處理過程中,3 d開始SOD活性提高,但轉基因與對照株系差異不顯著;6 d時,轉基因株系的SOD活性顯著高于對照株系;9 d開始,各株系的SOD活性下降,除TCP7-3外,其他各轉基因株系的SOD高于對照株系(圖1B)。0.8%NaHCO3脅迫過程中,3 d時,各株系的SOD活性達到了頂峰,TCP7-1、TCP7-2的SOD活性顯著高于對照株系;隨著脅迫時間延長,各株系的SOD活性明顯下降,但大部分時間點轉基因株系的SOD活性均高于對照株系(圖1C)。
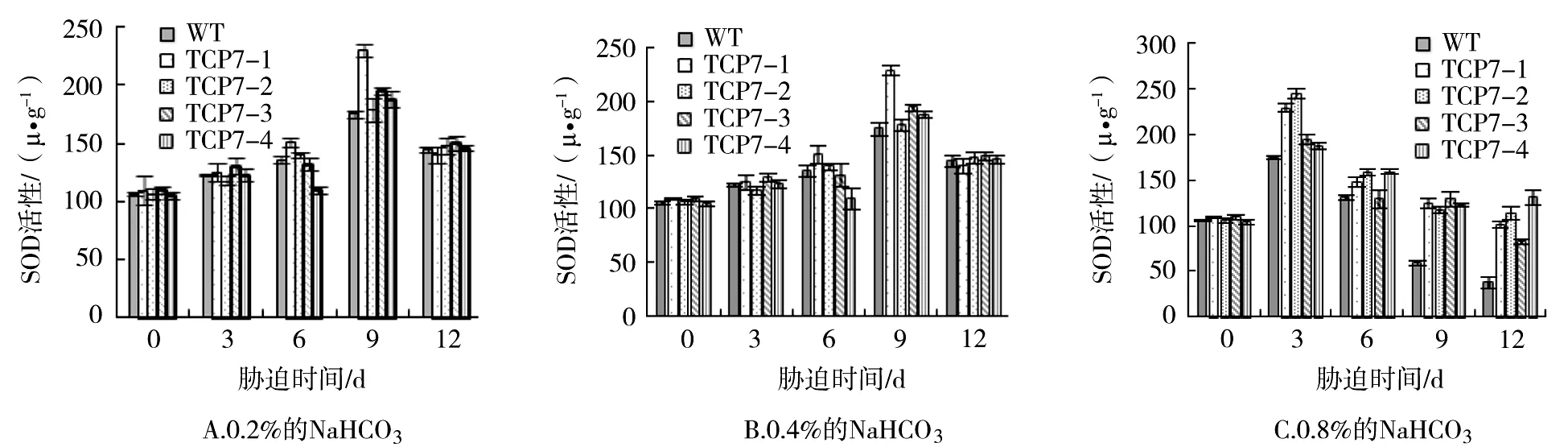
圖1 NaHCO3脅迫對轉基因及對照株系的SOD活性的影響
2.2 NaHCO3脅迫對脂膜的影響
MDA為脂膜過氧化的終端產物,其含量多少可以反映生物膜的危害程度,即MDA的含量越高,生物膜損害越嚴重[15]。從圖2A可以看出,低濃度NaHCO3脅迫6 d前,轉基因及對照株系的MDA含量變化均不明顯;脅迫9~12 d時,各株系MDA活性表現出上升的趨勢;在這2個時間點,各轉基因株系的MDA含量均低于對照株系,尤其是TCP7-2株系。在中濃度的NaHCO3處理過程中,6 d時各株系MDA的活性提高,但轉基因與對照株系差異不顯著;脅迫處理9 d時,大部分轉基因株系的MDA活性顯著高于對照株系(圖2B)。高濃度NaHCO3脅迫過程中,各株系的MDA含量隨著脅迫時間的延長逐漸上升,各株系的MDA含量均低于對照株系,尤其是TCP7-4株系(圖3C)。
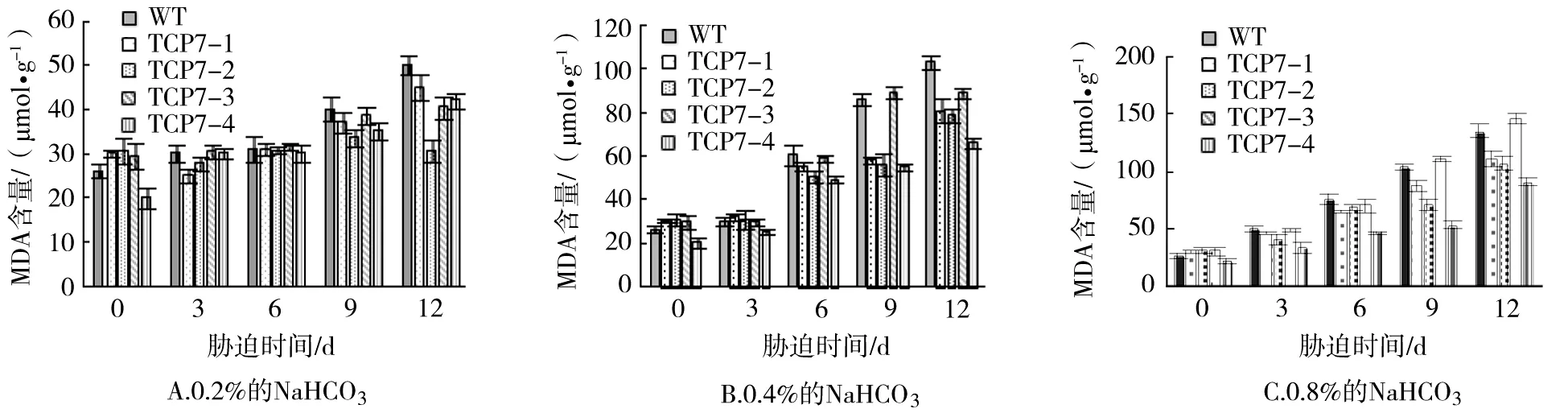
圖2 NaHCO3脅迫對轉基因及對照株系的MDA含量的影響
2.3 NaHCO3脅迫對滲透調節物質的影響
由圖3A可以看出,在0.2%NaHCO3脅迫3 d時,轉基因及對照株系的脯氨酸含量提高,但各株系間差異不顯著;脅迫6 d時,轉基因株系TCP7-3、TCP7-4顯著高于對照株系;脅迫9 d時,2個轉基因株系的脯氨酸含量下降,與其他各株系間無顯著差異。在0.4%NaHCO3脅迫3 d時,各株系的脯氨酸含量急劇上升,轉基因株系的含量高于對照株系;6 d時,除TCP7-3外,其他各轉基因株系的脯氨酸含量均高于對照株系,并達到最高峰;隨著脅迫時間的繼續延長,脯氨酸含量降低。0.8%NaHCO3脅迫后,3 d及6 d時各株系的脯氨酸含量均上升,除TCP7-1外,其他轉基因株系的上升幅度明顯高于對照株系。
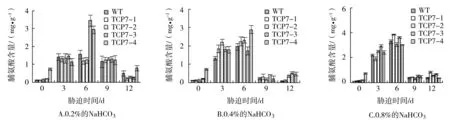
圖3 NaHCO3脅迫對轉基因及對照株系的脯氨酸含量的影響
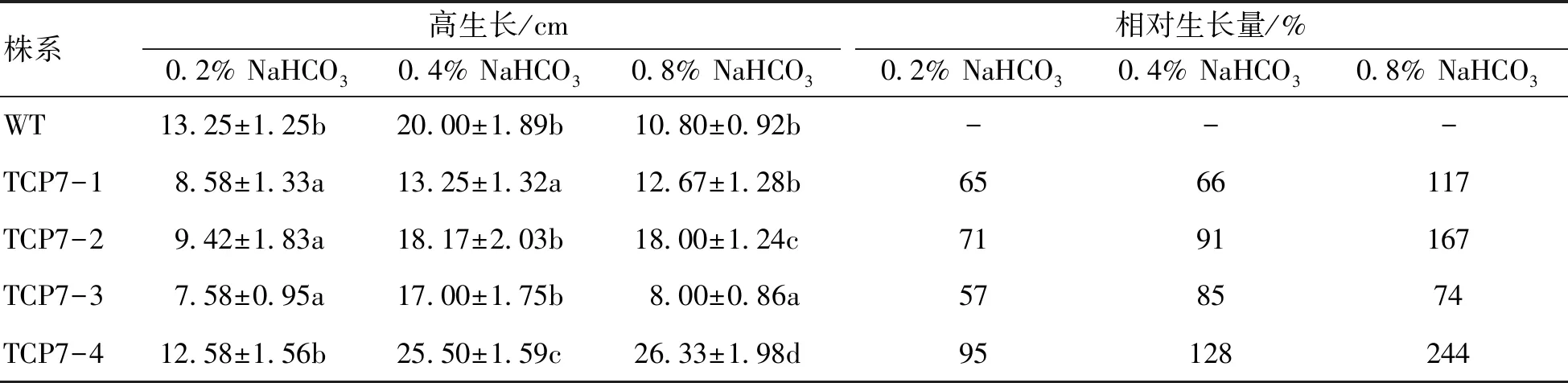
株系高生長/cm0.2% NaHCO30.4% NaHCO30.8% NaHCO3相對生長量/%0.2% NaHCO30.4% NaHCO30.8% NaHCO3WT13.25±1.25b20.00±1.89b10.80±0.92b---TCP7-18.58±1.33a13.25±1.32a12.67±1.28b6566117TCP7-29.42±1.83a18.17±2.03b18.00±1.24c7191167TCP7-37.58±0.95a17.00±1.75b8.00±0.86a578574TCP7-412.58±1.56b25.50±1.59c26.33±1.98d95128244
2.4 NaHCO3脅迫對植株高生長的影響
從表1可以看出,0.2%、0.4%的NaHCO3處理后,只有TCP7-4株系的高生長高于對照株系,其他3個轉基因株系均低于對照株系;但0.8%的NaHCO3處理后,除TCP7-3株系外,其他各株系的樹高生長量均高于對照株系,最高的是TCP7-4,相對生長量達到244%,TCP7-2次之,相對生長量是167%。
3 結論與討論
目前針對TCP的研究集中于發育調控,在抗逆方面的研究鮮見報道。OsTCP19在調節非生物脅迫中發揮了重要的作用,鹽和干旱條件下水稻中OsTCP19基因的表達量上調了5~6倍,并且在擬南芥中過量表達的OsTCP19引起ABI3、ABI4、IAA3的上調表達和LOX2下調表達,導致植株的發育異常[16]。在水曲柳中,研究FmTCP4對冷、鹽及旱的響應發現,FmTCP4明顯響應了這3種非生物脅迫,并且其響應模式并不相同。NaCl處理后,FmTCP4基因的表達呈現先上調后下調的趨勢,在6 h出現顯著上調,達到峰值,為對照的70.69倍[17]。在水稻中證明了過量表達或抑制OsTCP14(PCF6)和OsTCP21顯著降低或增加植物的冷脆弱性,且OsTCP14(PCF6)和OsTCP21在冷脅迫下負調控ROS介導的逆境脅迫反應[18]。在前期研究中發現,白樺TCP7啟動子中含有鹽、旱及ABA響應元件,通過NaCl及PEG處理后TCP7啟動子表現出一定的響應。本研究對轉TCP7白樺進行不同濃度的NaHCO3脅迫處理,發現低濃度的NaHCO3處理后轉基因及對照株系差別不明顯,但中、高濃度的NaHCO3處理后,對照株系的細胞膜損傷程度明顯高于轉基因株系,而滲透調節物及活性氧清除酶類的積累明顯低于轉基因株系,說明BpTCP7基因提高了白樺的抗鹽堿能力。
綜上所述,轉TCP7白樺株系在NaHCO3脅迫下,能有效誘導體內的保護酶系統,清除體內的活性氧,降低膜脂氧化損傷,一定程度上維持了植物正常的生理代謝活動。同時,通過體內滲透調節物質的積累,起到了滲透調節的作用,使得轉TCP7白樺對NaHCO3等逆境適應性增強。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
