無瓣海桑人工混交林結構及幼苗天然更新能力
2019-04-10 03:15:36黃曉敏盧昌義
福建林業科技 2019年1期
關鍵詞:植物
黃曉敏,盧昌義
(1.廈門大學環境與生態學院,福建 廈門 361102;2.河口生態安全與環境健康福建省高校重點實驗室(廈門大學嘉庚學院),福建 漳州363105)
無瓣海桑(Sonneratiaapetala)是隸屬于海桑科(Sonneratiaceae)海桑屬(Sonneratia)的紅樹樹種,由于其能在惡劣的灘涂上存活,并能改善灘涂的立地條件,因此1985年從孟加拉國引種至海南省東寨港。目前已在海南、廣東、廣西和福建等地大面積種植,是華南沿海生態恢復應用中最重要的紅樹樹種[1-3]。為豐富下層植物種類,提高林分穩定性,人工紅樹林常被進行多樣性改造。無瓣海桑人工混交林是否比紅樹純林具有更好的生態效益,能否使外來種無瓣海桑演替成上層植物,鄉土種喬木演替成中層植物,鄉土種灌木演替成下層植物的復合健康林分體系是恢復生態學者關注的重要問題[3]。陳玉軍等[4]在海南省三江河兩岸的調查表明無瓣海桑群落的物種組成和物種多樣性指標稍高于秋茄(Kandeliaobovata)群落,并且在無瓣海桑群落中僅有高齡級個體存在,而秋茄和桐花樹(Aegicerascorniculatum)為旺盛增長種群。王旭等[5]調查發現海南省東寨港無瓣海桑與鄉土種海桑(Sonneratiacaseolaris)混交林出現了自然更新的秋茄和桐花樹等鄉土紅樹植物,認為“引進種+鄉土種”的人工恢復模式可以促進鄉土紅樹林的更新。唐以杰等[5]認為混交林和無瓣海桑純林之間大型底棲動物群落的優勢種存在差異,在無瓣海桑林下混交紅海欖(Rhizophorastylosa)或木欖這2種鄉土紅樹植物,可提高林地底棲動物的生物量和物種多樣性。
福建省廈門市海滄灣無瓣海桑人工混交林于2002年種植,林內紅樹植物有無瓣海桑、秋茄、紅海欖和白骨壤(Avicenniamarina)。目前,該林分已生長16 a,是廈門市海灣典型的無瓣海桑人工混交林。黃龍嬌[7]研究了該地無瓣海桑樹種的交配系統和基因流情況;文玉葉[8]對該地無瓣海桑的生長情況進行了調查,并著重研究了其繁殖體特性。對于該地無瓣海桑人工混交林的林分結構和幼苗自然更新擴散能力情況未見詳細報道。為探究無瓣海桑人工混交林的生態效應問題,本文對廈門市海滄灣16年生無瓣海桑人工混交林結構和幼苗更新擴散能力進行實地調查,以期為今后紅樹林群落恢復和保護工作提供參考,也為引種外來紅樹植物的監控提供階段性的資料。
1 研究區自然概況
研究地點位于廈門市海滄區海滄大道未來海岸東側西海域中,與廈門島一水之隔,北鄰馬鑾灣,東鄰廈門西港,南面是九龍江入海口。地理坐標位于24°29′02″N、118°02′32″ E。試驗地紅樹林種植地土壤屬于沙質土,腳踩泥深度約8 cm,光灘地土壤屬于粘壤土,腳踩泥深度20 cm以上。潮汐屬正規半日潮,平均潮差3.99 m。高潮時海水平均鹽度為28.8‰。屬于南亞熱帶季風性氣候,年均氣溫21 ℃,年均日照時間達2233.5 h,年均降水量1036 mm,全年基本無霜。該地無瓣海桑人工混交林于2002年種植,秋茄的種植密度為50 cm×50 cm,無瓣海桑以2 m×2 m的種植密度間種于秋茄林中,紅海欖以2 m×2 m的種植密度間種于靠岸一側,4 m以內的秋茄林緣,少量白骨壤在秋茄林內混交。
2 調查方法
2.1 群落調查
參考方精云等[9]的經典樣方調查方法,于2018年6月對該群落進行樣方調查,根據樣地實際情況,隨機設置了3個5 m×10 m的樣方,對樣方內的喬木、灌叢、幼苗進行每木調查,分別記錄種名、高度、胸徑、地徑和冠幅。
2.2 重要值計算
根據高度及種群組成情況,將群落劃分為4層,高度≥7 m的為喬木一層,高度介于2.5~7 m的為喬木二層,高度介于1.3~2.5 m的為灌木-小樹層,高度<1.3 m的為幼苗層[10]。
喬木層和灌木-小樹層種群的重要值計算方法參照許會敏等[11]的計算方法。重要值=(相對密度+相對頻度+相對顯著度)/3。幼苗層種群的重要值,因沒有相對顯著度,則重要值=(相對密度+相對頻度)/2。
2.3 多樣性指數計算
參照伊賢貴等[12]計算多樣性指數的方法,根據樣方調查數據計算多樣性指數,包括Shannon-Wiener物種多樣性指數、Margalef豐富度指數、Simpson生態優勢度指數和Pielou均勻度指數。
2.4 幼苗天然更新和擴散調查
參考廖寶文等[13]調查幼苗更新和擴散情況的方法,于2018年6月分別在人工混交林內、林緣、光灘和互花米草(Spartinaalterniflora)地4種類型樣地中設置了52個2 m×2 m樣方,調查樣方內幼苗種類、數量、高度、地徑和冠幅。根據樣地實際情況,樣方設置具體為:在群落調查設置的3個大樣方內隨機設置4個樣方,共調查12個樣方;在無瓣海桑人工復合林東西兩邊林緣各從南到北隨機設置3個樣方,在北邊林緣按距林0~5、5~10、10~15、15~20 m 4個梯度,每個梯度設置2個樣方,在南邊林緣隨機設置2個樣方,共調查16個樣方;在光灘按距林緣0~2、2~4、4~6、6~8、8~10 m 5個梯度,每個梯度設置4個樣方,共調查20個樣方;在互花米草地隨機設置4個樣方。
3 結果與分析
3.1 林分結構特征
調查結果(表1)表明,該研究地共有紅樹植物3科4屬,包括海桑科的無瓣海桑,紅樹科的秋茄和紅海欖以及馬鞭草科的白骨壤。該群落外貌簡單,包括喬木層、灌木-小樹層和幼苗層。整個群落林相整齊,位于喬木層的無瓣海桑密度為0.05株·m-2,平均高度為7.13 m。其中位于喬木一層的是無瓣海桑,密度為0.03株·m-2,平均高度為8.5 m;位于喬木二層的有無瓣海桑、秋茄和紅海欖,密度分別為0.03、0.95、0.03株·m-2,平均高度分別為5.75、4.08、3.88 m。位于灌木-小樹層的有秋茄、紅海欖、白骨壤,密度分別為0.04、0.02、0.09株·m-2,平均高度分別為2.25、2.03、1.48 m。幼苗層只有秋茄幼苗,密渡為0.14株·m-2,平均高度為17 cm。
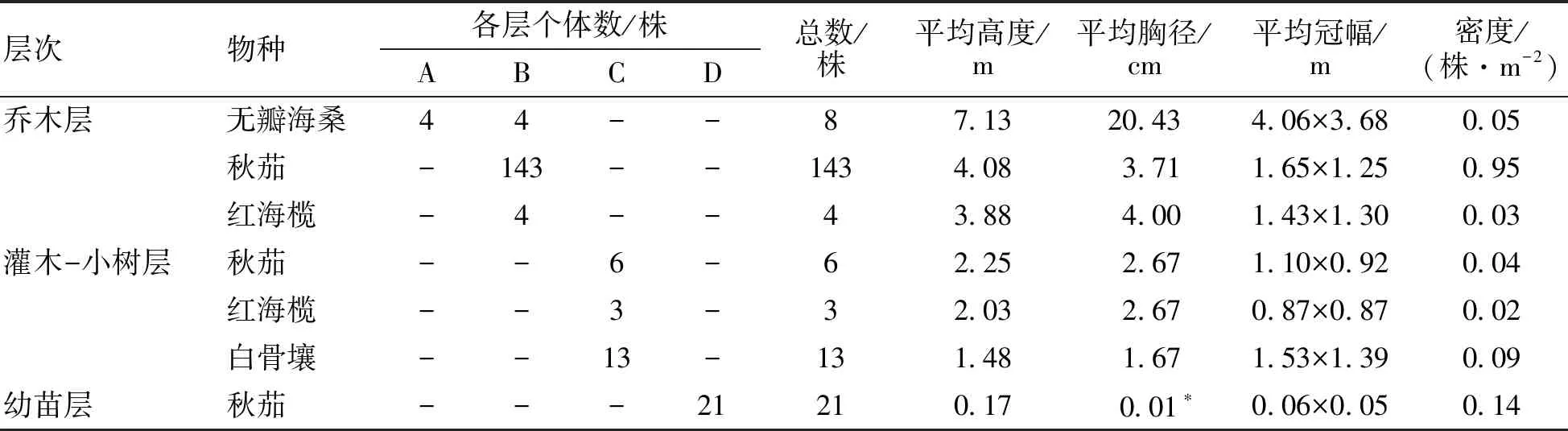
表1 無瓣海桑人工混交林林分結構特征
*:-為沒有調查;*為平均地徑(由于植株高度<1.3 m沒有胸徑);A為喬木一層;B為喬木二層;C為灌木-小樹層;D為幼苗層;下同。
3.2 重要值
重要值是評價某一種植物在濕地群落中作用的綜合性數量指標,是研究群落結構的重要指標之一,也是表征優勢種的重要指標[14]。由表2可以看出,在喬木層中無瓣海桑和秋茄的相對頻度都為40.0%,但由于秋茄的種群數量大于無瓣海桑的種群數量,所以秋茄的重要值最大,為57.1%,無瓣海桑的重要值為35.1%,而紅海欖的重要值最小。在灌木-小樹層,秋茄有小樹生長,并且重要值最大;白骨壤相對頻度雖較小,但其相對多度最大,所以其重要值也較大。紅海欖在灌木-小樹層的重要值比在喬木層的大。幼苗層只有秋茄生長。
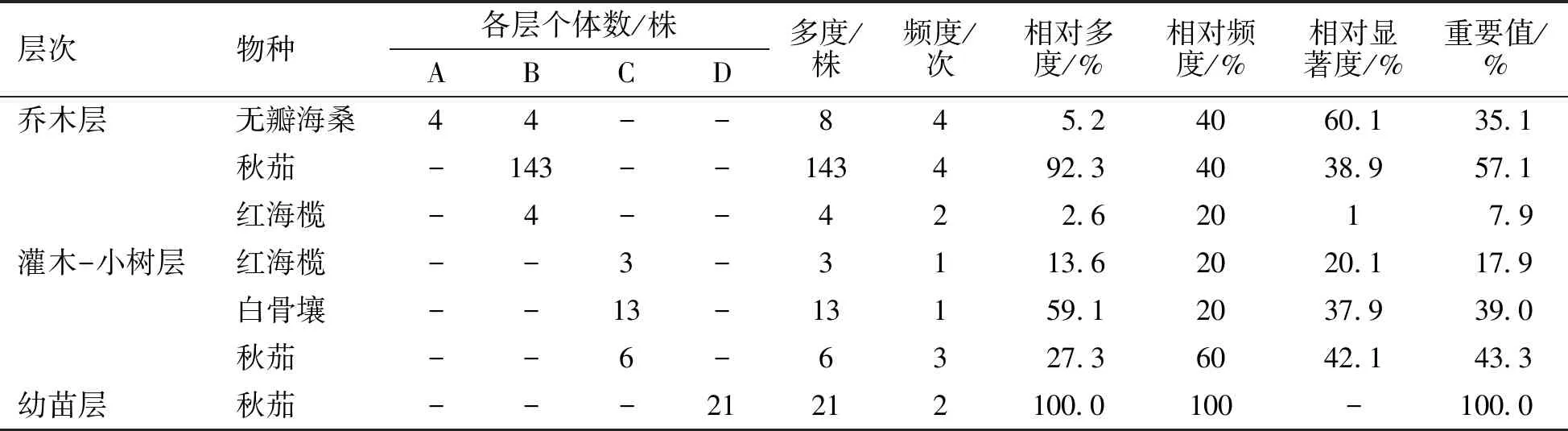
表2 無瓣海桑人工混交林不同層次物種重要值
3.3 物種多樣性
物種多樣性在某種程度上和生態系統之間的關聯性能夠反映生態系統的穩定[15]。由表3可以看出,該群落的多樣性指數值分別為:Shannon-Wiener指數值為0.56,Margalef指數值為0.57,Simpson指數值為0.26,Pielou指數值為0.41,物種多樣性較低。喬木層、灌木-小樹層和幼苗層3個層次的物種多樣性指數、豐富度指數、生態優勢度指數和均勻度指數變化趨勢較為一致,表現為灌木-小樹層>喬木層>幼苗層。
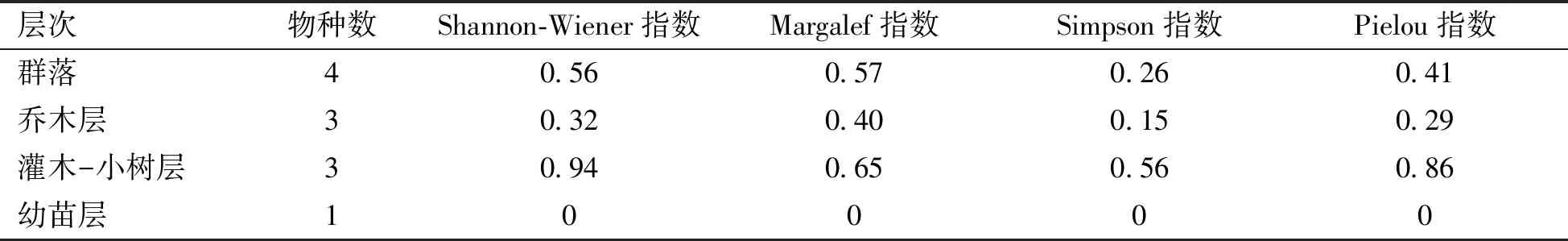
表3 無瓣海桑人工混交林整個群落和不同層次的多樣性指數
3.4 幼苗天然更新和擴散
在無瓣海桑人工混交林內、林緣、光灘和互花米草地4種類型樣地設置的52個2 m×2 m的樣方內,均未發現有無瓣海桑幼苗;而本地種秋茄和白骨壤都能進行天然更新和擴散。從表4可以看出秋茄在4種類型樣地中都進行了天然更新和擴散,其中秋茄幼苗在光灘分布最多,達1.11株·m-2,其次是互花米草地,為0.44株·m-2,最少的是在林緣,僅為0.28株·m-2。白骨壤幼苗只有在互花米草地中生長,密度為0.06株·m-2。
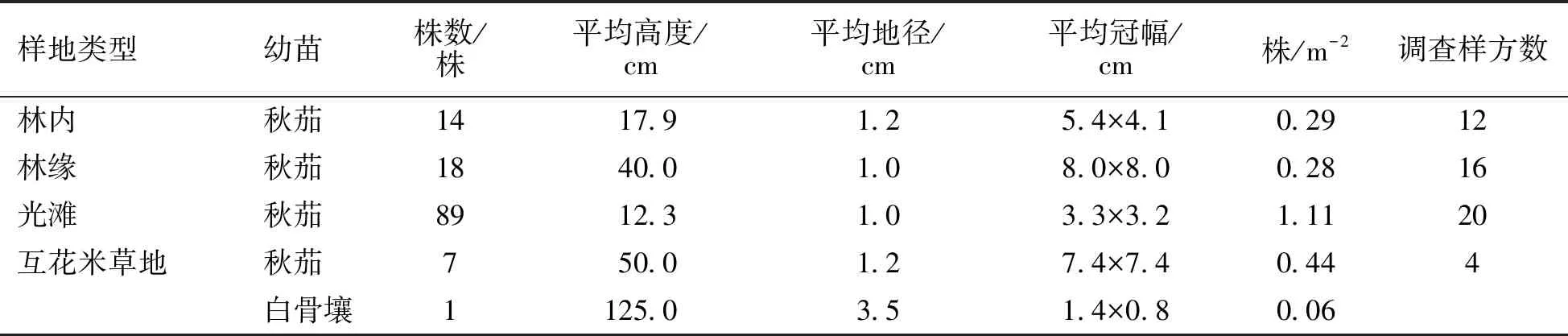
表4 調查樣地內鄉土紅樹植物幼苗情況
4 結論與討論
研究結果表明,該研究地群落空間層次分層明顯,各物種占據不同生態位,充分利用生境資源正常生長,在景觀搭配和防護效用上起著不同的協調互補作用。鄉土紅樹植物秋茄在該群落中占有絕對優勢,對當地的環境適應能力比外來物種無瓣海桑更強。采用“引進種+鄉土種”的人工種植恢復模式可增加群落的物種多樣性,使群落層次更加豐富,群落結構更加穩定,促進鄉土紅樹植物的天然更新和擴散。調查樣地未發現無瓣海桑幼苗,說明其在該地天然更新和擴散極其困難。無瓣海桑對鄉土紅樹植物生長和幼苗天然更新、擴散無抑制作用,并且能為鄉土紅樹植物的天然定植和生長創造有利條件。在人工林恢復建設中可考慮以鄉土樹種為主,引進種與鄉土種相結合的方式。但引進種植外來植物仍然有必要不斷跟蹤監控,以獲取更多的現狀和動態資料,全面掌握外來生物的引進和合理利用。
廈門市海滄灣無瓣海桑人工混交林于2002年人工種植,群落已有16 a。由于不同紅樹植物種類自身特性和對生境的要求不同[16],因此按照無瓣海桑、秋茄、紅海欖和白骨壤各自的生長特性,將其按一定位置人工種植在海灘上排列成帶。無瓣海桑和秋茄為這個群落的建群種,無瓣海桑相間種植在秋茄林中,白骨壤和紅海欖數量較少,分散在群落的較高潮位。群落空間層次分層明顯,位于喬木一層的只有無瓣海桑,密度為0.03株·m-2,平均高度為8.5 m;位于喬木二層的紅樹植物按平均高度從高到底排列依次是無瓣海桑、秋茄、紅海欖;位于灌木-小樹層的紅樹植物按平均高度從高到底排列依次是秋茄、紅海欖、白骨壤。無瓣海桑雖然位于喬木一層,但現存密度小,對其下層紅樹植物不造成遮光影響。各紅樹樹種在群落中占據不同生態位,充分利用生境資源正常生長,并且各自樹形有著明顯差異,在景觀搭配和防護效用上也起著不同的協調互補作用。幼苗層有鄉土紅樹植物秋茄幼苗的更新,而并未發現無瓣海桑幼苗和歷年各級小樹,可見無瓣海桑在林內自然更新成林困難,對鄉土紅樹植物的更新也沒有抑制作用,而且還為鄉土紅樹植物幼苗定植提供了有利條件,促進整個群落的自然更新。鄉土紅樹植物秋茄在喬木層、灌木-小樹層和幼苗層中均有分布,并且在各層次秋茄的重要值都為最大,可見鄉土紅樹植物秋茄在群落中占有絕對優勢,對當地的環境適應性較強。
該研究地各多樣性指數與珠海無瓣海桑人工林多樣性指數[17]比較可以發現,其數值略小于珠海的無瓣海桑人工林多樣性指數值,并且兩地物種多樣性指數均比陸地植被群落的物種多樣性低,主要因為海岸潮間帶環境惡劣,能夠生長的植物不多,并且也與灘涂生境、人工種植方式、種植年限以及演替程度有關。喬木層多樣性指數值比灌木-小樹層要低,由于喬木層各樹種數量分布不均勻,而灌木-小樹層的物種數量分布均勻,所以灌木-小樹層多樣性指數值大,群落結構較為穩定,優勢度大,是生長旺盛的層次。
在4種類型樣地內進行幼苗天然更新和擴散情況的調查發現,4種類型樣地中均未發現無瓣海桑幼苗。彭友貴等[1]研究發現鹽度高于5‰時,無瓣海桑種子發芽開始受到抑制;高于8‰時,幼苗生長開始受到抑制;鹽度高于15‰以上時,種子不能出土萌發。該研究地高潮時海水平均鹽度為28.8‰,廈門其他海域海水的鹽度更高,因此鹽度很可能是通過抑制無瓣海桑種子萌發、幼苗生長而限制其自然更新和擴散的主要原因。該研究樣地中鄉土紅樹植物秋茄和白骨壤都能進行天然更新和擴散,無瓣海桑對鄉土紅樹植物的天然更新成林和擴散沒有產生影響。林內和林緣秋茄幼苗密度較小,在光灘和互花米草地秋茄幼苗密度較大,并且在互花米草地出現了白骨壤幼苗,張喬民等[18]研究發現紅樹林可以生長在各種底質的海岸上,以淤泥質潮灘最普遍和生長最好。何琴飛等[19]研究也發現紅樹林生長較好的立地類型為土壤養分較高的內、中灘泥質或沙泥質土。而該研究地林內和林緣都為沙質土,光灘和互花米草地為淤泥質土,因此土壤質地很可能是影響鄉土紅樹植物幼苗擴散分布的原因。
*:感謝廈門大學環境與生態學院諸姮老師協助數據分析,感謝楊志偉老師提供研究區紅樹林的種植資料,感謝參與本次調查工作的廈門大學環境與生態學院王秀麗、宋瑞瑞、彭建同學。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
