普通煙草NtNAC072 基因的克隆、鑒定及表達模式分析
2019-04-09 01:30:00李曉旭郭永峰
煙草科技 2019年3期
李曉旭,郭 存,劉 成,李 虹,李 偉,蔣 勛,郭永峰*
1. 中國農(nóng)業(yè)科學院煙草研究所 煙草行業(yè)基因資源利用重點實驗室,山東省青島市嶗山區(qū)科苑經(jīng)四路11 號 266101
2. 鄭州大學生命科學院,鄭州高新技術產(chǎn)業(yè)開發(fā)區(qū)科學大道100 號 450001
轉(zhuǎn)錄因子(Transcription factor, TF)又稱為反式作用因子,能特異性地結合啟動子特定區(qū)域DNA序列(順式作用元件,Cis-acting element),激活或抑制下游基因的轉(zhuǎn)錄,從而調(diào)控相關的生物學過程。NAC 轉(zhuǎn)錄因子是一類植物特有的轉(zhuǎn)錄因子,其命名最初來源于矮牽牛NAM(No apical meristem)、擬南芥ATAF1-2 和擬南芥CUC1-2(Cup shaped cotyledon),以三者名稱的首字母組合為NAC[1]。NAC 轉(zhuǎn)錄因子家族成員在N 端具有高度保守的NAC 結構域,負責與特異性的順式作用元件結合,大約由150 個氨基酸組成,包含A、B、C、D、E 5 個亞結構域(Subdomain),其中,A、C 和D 在不同的物種間是高度保守的,B 和E 保守性相對較差,亞結構域D 中含有核定位信號(NLS),與NAC 轉(zhuǎn)錄因子的核定位相關。C 端為高度變異并具有轉(zhuǎn)錄激活功能的調(diào)控區(qū)[1-3]。
NAC 轉(zhuǎn)錄因子家族成員在植物生長發(fā)育中起重要作用[2]。最早報道的NAC 轉(zhuǎn)錄因子家族成員是矮牽牛中的NAM,矮牽牛nam 突變體在幼苗時期表現(xiàn)為頂端分生組織缺失、子葉融合,進而導致生長停滯[4]。在擬南芥中,NAC 轉(zhuǎn)錄因子CUC1和CUC2 高度同源,在頂端分生組織區(qū)域表達,cuc1/cuc2 雙突變體表型與矮牽牛nam 突變體類似,說明NAC 轉(zhuǎn)錄因子對于頂端分生組織的建立和維持具有重要作用[5-6]。在擬南芥中,NAC 轉(zhuǎn)錄因子家族成員編碼基因SND1 在莖木質(zhì)部纖維和束間纖維中特異性表達,參與次生細胞壁物質(zhì)合成的轉(zhuǎn)錄調(diào)控[7-8]。同時,NAC 轉(zhuǎn)錄因子家族成員廣泛參與生物、非生物逆境脅迫[9]。擬南芥中,由鹽脅迫、干旱脅迫和脫落酸ABA 誘導的基因ANAC072(RD26),過表達時能顯著提高轉(zhuǎn)基因擬南芥的抗旱能力[10-11]。在馬鈴薯中研究發(fā)現(xiàn),ANAC072 的馬鈴薯同源基因StNAC072 和StNAC101能夠被干旱脅迫和鹽脅迫所誘導,提示馬鈴薯StNAC072 和StNAC101 基因可能與非生物逆境響應相關[12]。擬南芥ANAC002(ATAF1)基因不但在響應干旱脅迫時發(fā)揮調(diào)控作用[13],同樣在非生物逆境脅迫中也發(fā)揮重要作用[14]。此外,擬南芥中NAC 轉(zhuǎn)錄因子家族基因ANAC029(NAP)在衰老時期表達量較高,誘導過表達該基因能夠促進葉片衰老,并且參與ABA 介導的信號通路[15-16]。最近的研究表明,參與非生物逆境應答的ANAC072,過表達該基因時能夠促進葉片衰老,暗示ANAC072基因可能同時調(diào)控非生物逆境應答和葉片衰老[17-18]。
有研究表明,葉片衰老受許多分子機制調(diào)控[19],而煙葉成熟落黃對外觀品質(zhì)、化學成分以及香氣特征等起重要作用。深入研究煙草葉片衰老落黃的調(diào)控機制,對煙草育種具有理論和實踐意義[20-21]。同時,煙草響應各種生物、非生物脅迫的分子機制已成為相關領域的研究熱點[22-23]。為此,采用比較基因組學方法,在普通煙草中克隆NtNAC072 基因,并對其保守結構域、進化關系、亞細胞定位、轉(zhuǎn)錄激活以及基因表達模式進行分析,為揭示NtNAC072 基因在煙草葉片衰老落黃及響應非生物脅迫的作用機理提供依據(jù)。
1 材料與方法
1.1 材料
普通煙草品種紅花大金元(紅大)種植在中國農(nóng)業(yè)科學院煙草研究所煙草行業(yè)基因資源利用重點實驗室溫室。在煙株現(xiàn)蕾期,采集植株莖、莖尖、根、根尖、幼嫩葉片、衰老葉片等,液氮冷凍后放置于-80 ℃冰箱保存,備用;在垂直MS 固體培養(yǎng)基中萌發(fā)幼苗,待長至6 片真葉期時,將其置于MS 液體培養(yǎng)基中培養(yǎng)5 d,隨后轉(zhuǎn)移至含有50 mmol/L NaCl、10 μmol/L ABA 的MS 溶液以及4 ℃和37 ℃光照培養(yǎng)箱中進行脅迫和激素處理,并將幼苗置于吸水濾紙上進行干旱處理,以未經(jīng)處理的煙苗為對照。分別在處理1、3、6 h 共3 個時間點采樣,每組處理采集3 株長勢相近的整株煙苗,液氮冷凍后放置于-80 ℃冰箱保存,備用。
1.2 試劑和儀器
農(nóng)桿菌GV3101 菌株、酵母AH109 菌株以及pCHF3-cGFP、pBridge 載體由本實驗室保存。RNAiso、PrimeScriptTMRT reagent Kit 反轉(zhuǎn)錄試劑盒 、SYBR? Premix Ex TaqTM熒 光 定 量 試 劑 、Infusion 連接酶、酵母基本培養(yǎng)基(SD Base)、色氨酸缺失混合物(-Trp)、X-Gal 等購自寶生物工程(大連)有限公司。引物合成和DNA 測序由上海派森諾生物科技有限公司完成。KOD 酶購自日本東洋紡公司。限制性內(nèi)切酶EcoRI、SacI 購自英國NEB 公司。pEASY-Blunt 載體、T1 感受態(tài)細胞、質(zhì)粒提取試劑盒、膠回收試劑盒購自北京全式金生物技術有限公司。
熒光定量PCR 儀ABI 7500(美國ABI 公司),激光共聚焦顯微鏡TCS-SP8(德國Leica 公司)。
1.3 方法
1.3.1 RNA 提取及反轉(zhuǎn)錄PCR
將不同組織樣品在液氮下研磨成粉末,按照說明書利用RNAiso 提取總RNA,使用DNaseⅠ去除基因組DNA 后,參照反轉(zhuǎn)錄試劑盒進行cDNA第一條鏈的合成,合成后的cDNA 稀釋至50 ng/μL待用。
1.3.2 基因NtNAC072 的克隆
以擬南芥ANAC072 蛋白序列為query 序列,在茄科基因組數(shù)據(jù)庫(http://solgenomics.net/)中進行BLASTP 比對,數(shù)據(jù)庫(Database)選擇N.tabacum K326 protein sequences,獲得候選基因(mRNA_47257_cds),根據(jù)候選基因CDS 序列,設計特異性擴增引物。上游引物:5'-ATGGGTGTTCAAGAAA AAGA-3',下游引物:5'-CTACTGCCTGAATCCCAG-3',以普通煙草品種紅大的根組織cDNA 為模板,使用KOD 酶進行PCR 克隆目的基因。PCR反應總體系為50 μL,其中PCR buffer 5 μL,dNTP 5 μL,鎂離子3 μL,引物各1.5 μL,KOD酶1 μL,cDNA 模板1 μL,用ddH2O 補充至50 μL。PCR 反應程序:94 ℃預 變性3 min,98 ℃ 變 性10 s,52 ℃ 退火30 s,68 ℃延伸1 min;32 個循環(huán),最終68 ℃延伸7 min。擴增得到目的片段,經(jīng)膠回收試劑盒回收。取4 μL 回收片段與1 μL pEASY-Blunt 混合,25 ℃反應15 min 進行連接,轉(zhuǎn)化到大腸桿菌感受態(tài)T1中,在含有100 μg/mL 卡那霉素的LB 平板上進行篩選,將PCR 鑒定為陽性的克隆送上海派森諾生物科技有限公司測序,將測序正確的亞克隆載體進行保菌處理,并命名為Blunt-NtNAC072。
1.3.3 生物信息學分析
候選基因所編碼的蛋白氨基酸序列由ProtParam 程 序(https://web.expasy.org/protparam/)預測其分子量、等電點等基本理化性質(zhì);分別在擬南芥數(shù)據(jù)庫TAIR(www.arabidopsis.org)和茄科基因組數(shù)據(jù)庫(http://solgenomics.net/)中下載擬南芥ANAC072、 ANAC019、 ANAC055、 ANAC002、ANAC081、ANAC029 和 馬 鈴 薯 StNAC072、StNAC101 等衰老及脅迫相關的NAC 轉(zhuǎn)錄因子蛋白序列[12],參照文獻[24]、利用MAFFT 在默認參數(shù)下進行多序列比對,使用Texshade 程序?qū)⒔Y果可視化,分析其保守結構域;在NCBI 中下載水稻等其他物種中衰老、脅迫相關NAC 轉(zhuǎn)錄因子蛋白序 列 ,包 括 水 稻SNAC1(AAT02360)、OsNAC4(BAA89798),玉米ZmNAC1(ADK25055),小麥TaNAC2(AAU08786),棉花GhNAC1(ACI15341)、GhNAC2(ACI15342),鷹嘴豆CarNAC4(ACS94037)、CarNAC5(ACS94038),番茄SlNAC1(AAR88435),利用MEME(http://meme-suite.org/)在默認參數(shù)下進行保守基序(motif)分析,并使用MAFFT 在默認參數(shù)下進行多序列比對,使用MGEA6.06 構建鄰接樹(Neighbor-Joining, NJ tree)。參數(shù)設置為:Bootstrap 檢驗次數(shù)設為1 000 次,并選擇Poisson Model 和Pairwise Deletion 進行分析。
1.3.4 亞細胞定位分析
參考文獻[25]進行亞細胞定位分析。將含有Blunt-NtNAC072 亞克隆載體的菌株擴大培養(yǎng)并提取質(zhì)粒,以該質(zhì)粒為模板進行PCR 擴增。使用Infusion 重組酶將不含終止密碼子的NtNAC072 基因CDS 序列,連接到pCHF3-cGFP 亞細胞定位載體上,構建35S 啟動子驅(qū)動下的C 端GFP 融合表達載體,轉(zhuǎn)化到大腸桿菌感受態(tài)T1后,在含有100 μg/mL壯觀霉素的LB 平板上進行篩選,將PCR 鑒定為陽性的克隆進行測序,將測序正確的重組載體轉(zhuǎn)化農(nóng)桿菌GV3101,在含有100 μg/mL 壯觀霉素和50μg/mL 利福平的YEP 平板上篩選,通過PCR 進行陽性鑒定,以含有35::GFP 質(zhì)粒的農(nóng)桿菌菌株為對照,將兩者分別在YEP 液體培養(yǎng)基中震蕩培養(yǎng)至OD600約為0.8,5 000 r/min 離心5 min 集菌,用重懸液(150 μmol/L 乙酰丁香酮,10 mmol/L MES,10 mmol/L MS,pH 5.6)重懸并注射到煙草葉片背部,培養(yǎng)3 d 后取適量注射葉片材料在激光共聚焦顯微鏡下觀察GFP 信號,激發(fā)光波長488 nm。
1.3.5 轉(zhuǎn)錄激活實驗
參考文獻[26]進行轉(zhuǎn)錄激活分析。以Blunt-NtNAC072 質(zhì)粒為模板,進行PCR 擴增,使用Infusion 重組酶將NtNAC072 基因CDS 全長序列,連接到pBridge 載體EcoRI 位點中,在含有100 μg/mL氨芐青霉素的LB 平板上進行篩選,PCR 篩選陽性克隆并測序驗證,構建NtNAC072 與GAL4BD 融合表達載體。將重組質(zhì)粒與pBridge 空白載體分別轉(zhuǎn)入酵母AH109 菌株感受態(tài)中,在-Trp 營養(yǎng)缺陷培養(yǎng)基(SD/-Trp)上進行篩選,隨后將所篩選的陽性克隆轉(zhuǎn)移至顯色培養(yǎng)基(SD/-Trp/X-gal)上倒置避光培養(yǎng),直至顯色。顯藍色的酵母菌斑,表明其轉(zhuǎn)入片段能夠啟動報告基因表達,具有轉(zhuǎn)錄激活活性。
1.3.6 NtNAC072 基因表達模式分析
利用熒光定量PCR 反應來檢測NtNAC072 基因的表達模式。根據(jù)該基因的CDS 序列設計特異性qRT-PCR 引物,上游引物:5'-GAATATCCGAGG TAATGCCAAGGTAG-3',下游引物:5'-GTCTGGTT AGGACACTGAATCCCAAA-3',采用煙草26S rRNA基因作為管家基因[27]。培養(yǎng)基中培育的煙苗經(jīng)脅迫、激素處理分別平行收集3 組樣品為3 次生物學重復,每組樣品分別提取RNA 并反轉(zhuǎn)錄為cDNA,定量分析時設計3 次技術重復,計算其平均值及標準差并根據(jù)2-ΔΔCt法計算基因的相對表達量。
2 結果與分析
2.1 NtNAC072 基因全長CDS 序列的克隆及序列分析
基于茄科基因組數(shù)據(jù)庫BLASTP 比對信息,設計候選基因特異性引物,以紅大葉片cDNA 為模板進行PCR 擴增,目的片段大小在1 000 bp 左右,泳道1 為不含cDNA 模板的陰性對照(圖1)。將目的片段回收后連接至pEASY-Blunt 載體,挑取單克隆進行測序。測序結果表明,NtNAC072 基因的CDS 序列全長1 065 bp,編碼354 個氨基酸。ProtParam 工具分析該蛋白的相對分子質(zhì)量為40 266.28 Da,理論等電點(Theoretical pI)為8.99。
基于新鑒定的NtNAC072 蛋白序列,將其與擬南芥、馬鈴薯和水稻等物種中衰老、逆境脅迫相關的NAC 轉(zhuǎn)錄因子ANAC072 等蛋白序列進行多序列比對(圖2),完全保守的氨基酸殘基用紫色標注,其余的相對保守的氨基酸殘基用藍色、紅色標注,發(fā)現(xiàn)NtNAC072 在N 端具有保守的典型NAC結構域。該結構域可以進一步分為A~E 共5 個亞結構域,在其亞結構域D 中發(fā)現(xiàn)保守的核定位信號,C 端序列則呈現(xiàn)高度多樣性,其可能與轉(zhuǎn)錄激活功能相關。
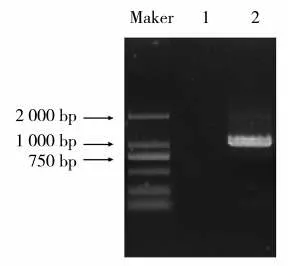
圖1 NtNAC072 基因PCR 擴增產(chǎn)物電泳圖Fig.1 Electrophoretogram of NtNAC072 PCR amplification


圖2 NtNAC072 轉(zhuǎn)錄因子NAC 結構域分析Fig.2 NAC conserved domain analysis of NtNAC072 transcription factor
2.2 NtNAC072 轉(zhuǎn)錄因子的進化分析和保守基序分析
以擬南芥、馬鈴薯和水稻等物種中已報道的衰老、脅迫NAC 轉(zhuǎn)錄因子為參考進行多序列比對,基于比對結果利用MEGA 構建了鄰接樹。基于重建的鄰接樹(圖3),參與進化分析的18 個NAC 轉(zhuǎn)錄因子家族成員被分為NAP、RD26 和ATAF 共3 個亞家族(Subgroup)。煙草NtNAC072與馬鈴薯StNAC072、擬南芥ANAC072 聚在一起,說明三者直系同源。利用序列保守基序識別工具MEME 對蛋白序列的保守基序進行分析,結果表明,同一亞家族內(nèi)的NAC 轉(zhuǎn)錄因子家族成員的保守基序種類和組織形式具有較強的一致性,從側(cè)面反映了系統(tǒng)進化分析的可靠性。同時,NtNAC072 與StNAC101、StNAC072 和ANAC072保守基序的種類和組織形式高度一致,暗示NtNAC072 與StNAC101、StNAC072 和ANAC072可能具有相似的功能。
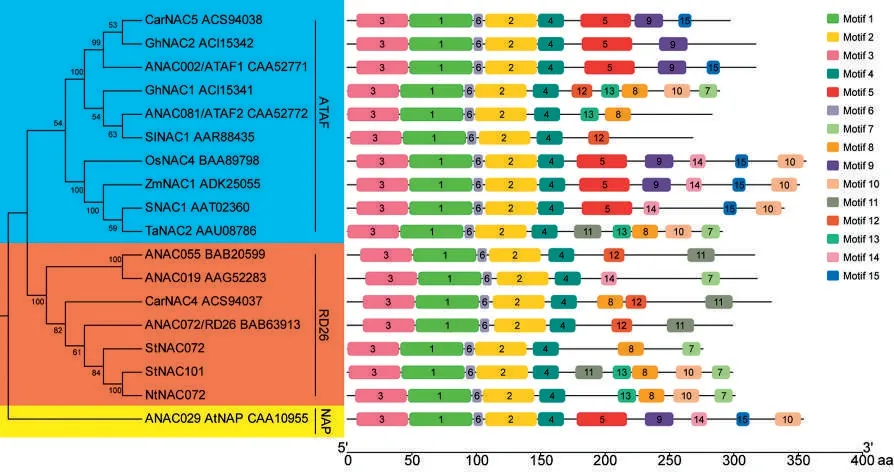
圖3 NtNAC072 轉(zhuǎn)錄因子系統(tǒng)進化及保守基序分析Fig.3 Phylogenetic and conserved motifs analysis of NtNAC072 transcription factor
2.3 NtNAC072 轉(zhuǎn)錄因子的亞細胞定位分析
通過農(nóng)桿菌侵染,讓NtNAC072 基因在煙草葉片中瞬時表達,確定NtNAC072 轉(zhuǎn)錄因子的亞細胞定位(圖4)。其中,DAPI 指示細胞核,BF 為明場。在35::GFP 瞬時轉(zhuǎn)化對照中,GFP 信號遍布整個細胞,而融合NtNAC072 的GFP 蛋白信號只能在細胞核中觀察到,并與細胞核特異性染料DAPI 的信號重疊,這說明NtNAC072 是一個核定位蛋白,符合其轉(zhuǎn)錄因子的特性。
2.4 NtNAC072 轉(zhuǎn)錄因子的轉(zhuǎn)錄激活分析

圖4 NtNAC072 的亞細胞定位Fig.4 Subcellular localization of NtNAC072
pBridge 中含有酵母轉(zhuǎn)錄因子GAL4 的BD(binding domain)序列,其編碼的蛋白能和對應酵母菌株AH109 中的UAS 序列特異性結合,如果GAL4-BD 融合的目的蛋白具有轉(zhuǎn)錄激活活性,則能夠啟動UAS 下游的報告基因,因此能夠利用報告基因表達與否,判斷融合片段是否具有轉(zhuǎn)錄激活活性。將pBridge 空載對照和NtNAC072-pBridge載體分別轉(zhuǎn)化AH109 酵母菌株,在SD/-Trp 平板上,30℃條件下培養(yǎng)3 d 篩選,隨后將篩選的陽性酵母克隆轉(zhuǎn)移至顯色培養(yǎng)基(SD/-Trp/X-gal)上倒置避光培養(yǎng)。從圖5 中可以看出,含有NtNAC072的全長CDS 的酵母表達菌株在X-gal 存在的情況下,能夠啟動報告基因表達進而顯示藍色。而只含有pBridge 的對照酵母菌株不能激活報告基因表達,說明NtNAC072 具有轉(zhuǎn)錄激活活性。

圖5 NtNAC072 酵母轉(zhuǎn)錄激活實驗Fig.5 Transactivation analysis of NtNAC072 in yeast
2.5 NtNAC072 基因的表達模式分析
實時熒光定量PCR 結果表明,NtNAC072 基因在莖(ST)、莖尖(SAM)、根(RT)、根尖(RAM)、幼葉(YL)和衰老葉片(SL)所有測試組織中均有表達,主要在煙草衰老葉片和根等部位表達(圖6)。當煙草葉片衰老時NtNAC072 基因表達量顯著上升,這說明該基因可能參與煙草葉片衰老落黃。
對NtNAC072 基因在不同脅迫及激素處理下的表達分析結果可見,NtNAC072 基因的表達受多種脅迫的誘導(圖6),如:鹽脅迫(NaCl)、干旱脅迫等,其中干旱脅迫誘導作用最為明顯。在干旱脅迫下,NtNAC072 基因的表達量隨著干旱處理時間的延長而顯著增高,在6 h 時達到對照(CK)的12.6倍。在鹽脅迫下,NtNAC072 基因的轉(zhuǎn)錄水平也隨著鹽脅迫處理時間的延長而增高,并且在處理3 h內(nèi)急劇上升,為對照的5.7 倍左右,隨后略微下降。此外,ABA 能夠顯著誘導NtNAC072 基因的表達,在處理6 h 后,達到對照的11.1 倍。值得注意的是,冷脅迫和熱脅迫處理時,NtNAC072 基因的表達情況變化不明顯,說明NtNAC072 基因主要受到鹽脅迫和干旱脅迫的誘導,可能在煙草鹽脅迫和干旱脅迫等非生物逆境應答方面發(fā)揮重要調(diào)控作用。

圖6 NtNAC072 基因的表達模式分析Fig.6 Expression pattern analysis of NtNAC072
3 結論
利用比較基因組學方法,從普通煙草中分析并克隆得到擬南芥ANAC072 的同源基因NtNAC072。生物信息學分析發(fā)現(xiàn),NtNAC072 蛋白具有保守的典型NAC 結構域,在其亞結構D 中發(fā)現(xiàn)保守的核定位信號。進化分析發(fā)現(xiàn),NtNAC072 與擬南芥ANAC072 直系同源,并且保守基序具有很強的一致性。亞細胞定位實驗與轉(zhuǎn)錄激活實驗顯示,NtNAC072 定位在細胞核內(nèi)且具有轉(zhuǎn)錄激活活性。表達模式分析發(fā)現(xiàn),NtNAC072 基因主要是在根部和衰老葉片中表達,并且衰老葉片中的表達量明顯上升。在鹽脅迫處理下,NtNAC072 基因的表達峰值約為對照的5.7 倍。在干旱脅迫下,NtNAC072 基因的表達量隨著處理時間的延長而逐漸升高,表達峰值約為對照的12.6 倍。ABA 作為一個重要的信號分子參與植物的脅迫響應,ABA處理能夠誘導NtNAC072 的表達,且表達量顯著增加。因此,NtNAC072 基因為擬南芥ANAC072 的同源基因,編碼典型NAC 型的轉(zhuǎn)錄因子,在衰老落黃的煙草葉片中高量表達,并受到鹽脅迫、干旱脅迫和ABA 的誘導。NtNAC072 基因可能在煙草葉片衰老落黃以及鹽脅迫、干旱脅迫等非生物逆境應答方面發(fā)揮重要調(diào)控作用。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
浙江中西醫(yī)結合雜志(2017年2期)2017-01-12 18:23:59
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
聲屏世界(2014年6期)2014-02-28 15:18:09
終身教育研究(2014年5期)2014-02-28 01:23:06
中國煙草學報(2012年5期)2012-04-12 06:21:18
