鹽單胞菌S62 β-半乳糖苷酶合成低聚半乳糖的研究
2019-04-09 12:30:48朱五二繆明永顧正華李由然丁重陽石貴陽
生物加工過程 2019年2期
朱五二,繆明永,顧正華,李由然,丁重陽,石貴陽
(1. 江南大學生物工程學院糖化學與生物技術教育部重點實驗室,江蘇無錫214122; 2. 江南大學糧食發(fā)酵工藝與技術國家工程實驗室,江蘇無錫214122; 3. 第二軍醫(yī)大學生物化學與分子生物學教研室,上海200433)
β-半乳糖苷酶(EC 3.2.1.23)俗稱乳糖酶(lactase),屬于糖基水解酶類(glycosyl hydrolases),可以水解β-1,4-糖苷鍵,同時還具有半乳糖苷轉移酶活性[1],用于功能性食品低聚半乳糖(GOS)的合成。β-半乳糖苷酶廣泛存在于微生物(細菌、真菌和酵母)、植物(尤其是杏仁、桃、杏和蘋果)和動物器官中[2-4]。微生物酶具有較高的生產力,能降低生產成本。目前已有古細菌、細菌、真菌和酵母的β-半乳糖苷酶研究報道[5],從真菌(米曲霉和黑曲霉)和酵母(乳酸克魯維酵母和脆弱克魯維酵母)獲得的酶顯示出巨大的商業(yè)潛力,來自乳酸克魯維酵母的β-半乳糖苷酶是使用最廣泛的酶之一[6-9]。
低聚半乳糖是以乳糖作為供體和受體底物,利用β-半乳糖苷酶的轉糖基化活性產生的具有高達9聚合度(DP)的寡糖混合物[9],主要結構由葡萄糖末端連接可變數目的半乳糖單元組成,半乳糖單元的數目通常為2~5[10],半乳糖單元通過β(1→3),β(1→6)和β(1→4)鍵連接,而半乳糖-葡萄糖鍵在大多數情況下是β(1→4)[11]。不同來源的酶合成低聚半乳糖是有差異的,其中低聚半乳糖主要包含三糖、四糖、五糖,以及異乳糖和半乳二糖[12]。
低聚半乳糖是一種調節(jié)結腸微生物菌群的益生元,具有改善礦物質吸收和預防結腸癌的功效[13]。研究表明,低聚半乳糖大大增加了雙歧桿菌的數量及其在腸道中的代謝活性[14-16],可降低過敏發(fā)生率[17-18]和病原體的黏附[15,18-19],并且能介導腸道免疫系統(tǒng)。此外,低聚半乳糖有效治療代謝性疾病[20]。因此,低聚半乳糖廣泛應用在飲料和嬰兒奶粉的配方中[21]。
我國目前仍存在β-半乳糖苷酶產量低的問題,從而限制了β-半乳糖苷酶在工業(yè)中的應用。如何尋找一種高產、高酶活的β-半乳糖苷酶仍是國內研究的難點。繆明永課題組的Wang等[22]對中國東海海水和海泥來源的海洋微生物進行篩選,獲得了1株產S62β-半乳糖苷酶的鹽單胞菌S62(Halomonassp. S62),并對其低溫β-半乳糖苷酶基因(GenBank登錄號JQ337961)進行了重組表達研究。
在以上基礎上,筆者對S62β-半乳糖苷酶合成低聚半乳糖的反應條件進行探索,以獲得最大低聚半乳糖產率,以期為利用低溫β-半乳糖苷酶進行工業(yè)生產低聚半乳糖提供重要的理論依據。
1 材料與方法
1.1 試劑與儀器
一水合乳糖、葡萄糖、半乳糖,國藥集團化學試劑有限公司;乙腈,TEDIA公司。
電熱恒溫水浴鍋,上海醫(yī)用恒溫設備廠;超高效液相色譜串聯(lián)四級桿飛行時間質譜儀(UPLC-Q-TOF-MS)、高效液相色譜儀(HPLC),美國Waters公司。
1.2 低聚半乳糖的檢測
1.2.1 低聚半乳糖種類檢測
采用 Waters Acquity BEH Amide柱(2.1 mm × 100 mm,1.7 μm)分離,質譜檢測器,時間為20 min。流動相A相:80%乙腈加0.1%氨水;B相:30%乙腈加0.1%氨水。柱溫45 ℃,流速0.3 mL/min,進樣量1 μL。線性梯度洗脫程序見表1。
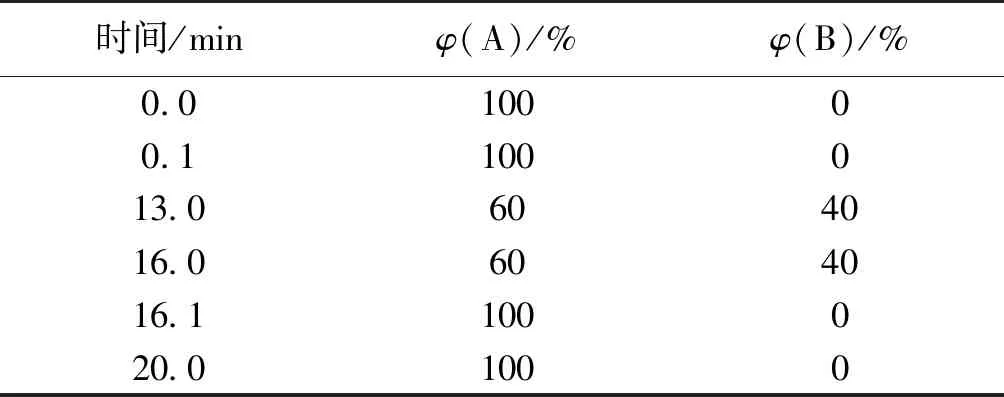
表1 LC-MS洗脫條件
注:質譜離子化方式為負離子(ESI-),毛細管電壓為3.5 kV,錐孔電壓為20 V,脫溶劑氣溫度為400 ℃,脫溶劑氣流速為700 L/h,碰撞能量為6 eV,質量范圍為10~1 000,檢測電壓為1 800 V。二級質譜碰撞能量為6 eV,錐孔電壓為50 V,選擇離子質荷比為341、503、665。
1.2.2 低聚半乳糖含量的檢測
實驗采用雙柱法檢測[23-24],并計算低聚半乳糖含量。
葡萄糖和半乳糖檢測:用伯樂公司的Aminex HPX-87C柱(300 mm×7.8 mm,9 μm)測出葡萄糖和半乳糖含量,流動相為純水,檢測器為示差檢測器,流速為0.6 mL/min,柱溫為50 ℃,檢測器靈敏度為8,檢測器溫度為30 ℃,進樣量為20 μL。
乳糖檢測:使用Waters公司的Amide柱(4.6 mm × 250 mm,5 μm)測出乳糖含量,流動相為70%乙腈,檢測器為示差檢測器,流速為1.0 mL/min,柱溫為40 ℃,檢測器靈敏度為8,檢測器溫度為30 ℃,進樣量為20 μL。
低聚半乳糖質量分數計算見式(1)。
w(GOS)=w(Lac)-w1(Gal)-w1(Glu)-
w(H+)-w1(Lac)
(1)
式中:w1(Gal)=w(Gal)×(180-17)/180;
w1(Glu)=w(Glu)×(180-1)/180;
w(H+)=[n(Glu)-n(Gal)]×1。
w(Gal)、w(Glu)、w(Lac)、w1(Lac)分別為水解半乳糖質量分數、水解葡萄糖質量分數、初始乳糖質量分數、反應后乳糖質量分數;n(Glu)、n(Gal)分別為葡萄糖和半乳糖的摩爾數。
乳糖消耗率和低聚半乳糖產率的計算見式(2)和(3)。
乳糖消耗率=w1(Lac)/w(Lac)×100%
(2)
低聚半乳糖產率=w(GOS)/w(Lac)×100%
(3)
1.3 單因素條件優(yōu)化
1.3.1 pH對低聚半乳糖合成的研究
用磷酸緩沖液配制pH為6.0~8.0的乳糖溶液,質量濃度為250 g/L,用純水配制酶液600 U/mL,取1.9 mL乳糖溶液加入0.1 mL酶,在30 ℃下反應6 h。計算出低聚半乳糖產率及乳糖消耗率。
1.3.2 底物濃度對低聚半乳糖合成的研究
采用單一變量法控制反應,底物質量濃度為150~350 g/L,pH為7.0,其他條件如1.3.1節(jié)所述。計算出低聚半乳糖產率及乳糖消耗率。
1.3.3 溫度對低聚半乳糖合成的研究
采用單一變量法控制反應,反應溫度為25~45 ℃,pH為7.0,其他條件如1.3.1節(jié)所述。計算出低聚半乳糖產率及乳糖消耗率。
1.3.4 反應時間對低聚半乳糖合成的研究
采用單一變量法控制反應,反應時間為4~12 h,pH為7.0,其他條件如1.3.1節(jié)所述。計算出低聚半乳糖產率及乳糖消耗率。
1.3.5 加酶量對低聚半乳糖合成的研究
采用單一變量法控制反應,加酶量為15~75 U/mL,pH為7.0,其他條件如1.3.1節(jié)所述,計算出低聚半乳糖產率及乳糖消耗率。
1.4 低聚半乳糖合成正交試驗
在前期的基礎上,控制反應時間為4 h,以溫度、底物濃度、pH、加酶量為考察因素,以低聚半乳糖含量為指標,進行L9(34)的正交試驗。
1.5 最適條件低聚半乳糖合成及酶穩(wěn)定性
在正交最適條件下進行反應8 h,定時取樣,使用HPLC測糖組分,并測定40 ℃下酶的穩(wěn)定性。
2 結果與討論
2.1 低聚半乳糖種類鑒定
通過液相色譜-質譜(LC-MS)鑒定S62β-半乳糖苷酶合成低聚糖的種類,結果如圖1所示。
由圖1可知:出峰時間8.71、9.17和9.72 min為3種二糖,其質譜圖依次為b、c、d,分別為乳糖、異乳糖和半乳二糖;出峰時間11.30和12.25 min為2種低聚半乳三糖,質譜圖為e、f;出峰時間13.51和14.23 min為2種低聚半乳四糖,質譜圖為g、h。此前,Rodriguez-Colinas等[25]鑒定了來自乳酸克魯維酵母的β-半乳糖苷酶產生的低聚半乳糖混合物中的5種低聚半乳糖。Urrutia等[26]從米曲霉的β-半乳糖苷酶產生的低聚半乳糖混合物中發(fā)現了9個種類。Yanahira等[27]從環(huán)狀芽孢桿菌的β-半乳糖苷酶產物中分離出11種低聚半乳糖。S62β-半乳糖苷酶以乳糖為底物合成了6種低聚半乳糖,可以看出S62β-半乳糖苷酶同樣具有較高的轉糖基酶活。
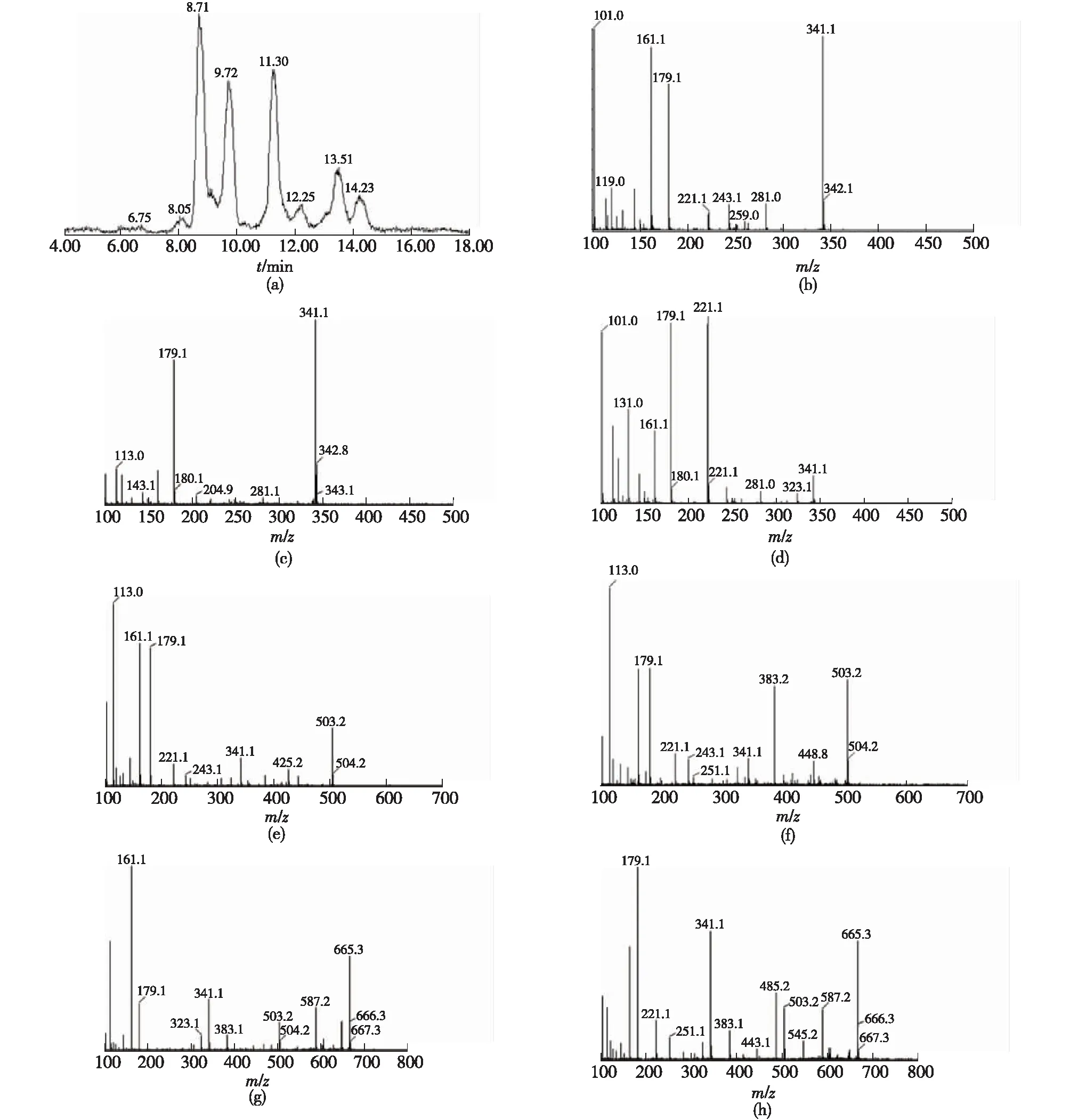
(a)—選擇離子色譜圖;(b)、(c)、(d)—二糖質譜圖;(e)、(f)—三糖質譜圖;(g)、(h)—四糖質譜圖圖1 低聚半乳糖種類分析Fig.1 Analysis of types of GOS
2.2 pH對低聚半乳糖合成的影響
Torres等[5]總結了細菌β-半乳糖苷酶轉糖基反應最適pH為6.0~8.0。以磷酸鹽緩沖液控制反應體系pH,考察pH對低聚半乳糖合成的影響,結果如圖2所示。
由圖2可知:S62β-半乳糖苷酶在pH 6.0~8.0穩(wěn)定性較好,表現出較好的轉糖基作用,低聚半乳糖產率為26%~30%,乳糖轉化率為69%~75%,其中在pH為7.0時,有最大低聚半乳糖產率(29.46±0.66)%,乳糖消耗率為(73.40±0.85)%。因此,在此反應體系中,最適pH為7.0。
2.3 底物濃度對低聚半乳糖合成的影響
在乳糖存在時,β-半乳糖苷酶存在著水解活性與轉糖基化活性相互競爭現象,即當反應開始時,乳糖濃度較高,轉糖基作用占主導作用,但是隨著反應進行,水解活性逐漸增強,最終生成葡萄糖和半乳糖[28]。由此可見高濃度的乳糖有利于低聚半乳糖的合成。因此,選擇150~350 g/L的乳糖,研究其對低聚半乳糖合成的影響,結果如圖3所示。
由圖3可知:當乳糖質量濃度大于200 g/L時,乳糖轉化率隨著底物濃度的增加逐漸降低,其原因主要是在一定的反應時間內,底物過飽和,酶結合底物能力有限,進而轉化率下降;而底物質量濃度為150 g/L時,底物濃度降低,酶水解活性增強,導致乳糖和低聚半乳糖被降解為葡萄糖和半乳糖。所以當酶量為30 U/mL,反應時間為6 h,底物質量濃度為200 g/L,其低聚半乳糖產率為(26.57±0.36)%,乳糖消耗率最高為(67.40±0.55)%。
2.4 溫度對低聚半乳糖合成的影響
低溫酶在低溫條件下通常保留著較高的酶活。Fan等[29]等研究了來自Rahnellasp. R3的低溫β-半乳糖苷酶,以乳糖為底物,在4~35 ℃的寬溫度范圍內顯示高活性;Coker[30]報道了AntarcticArthrobacterβ-半乳糖苷酶以乳糖為底物時最適為15~20 ℃。筆者研究了溫度對低聚半乳糖合成的影響,結果如圖4所示。
由圖4可知:乳糖存在時,S62β-半乳糖苷酶同樣在25~45 ℃時,表現出較高的酶活,最適轉糖基化溫度為40 ℃,此時低聚半乳糖產率為(36.15±0.01)%,乳糖消耗率為(82.45±0.01)%。溫度過高時,S62β-半乳糖苷酶在較短的時間內失活,導致溫度為45 ℃時的酶活比40 ℃時的酶活下降明顯。
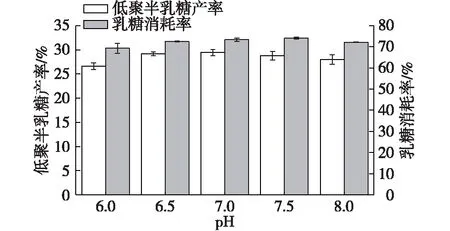
圖2 pH對低聚半乳糖合成的影響Fig.2 Effects of pH on GOS synthesis
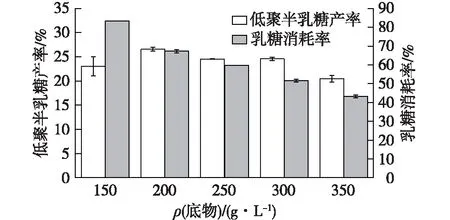
圖3 乳糖濃度對低聚半乳糖合成的影響Fig.3 Effects of lactose concentration on GOS synthesis
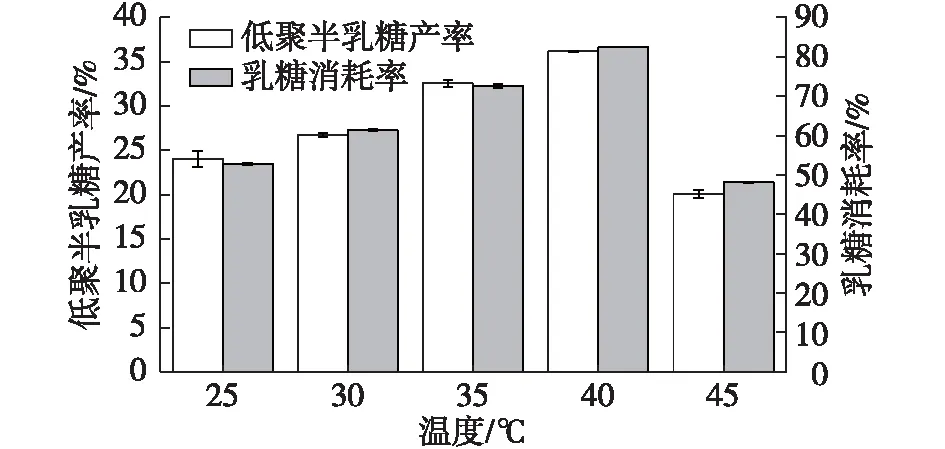
圖4 溫度對低聚半乳糖合成的影響Fig.4 Effects of temperature on GOS synthesis
2.5 加酶量對低聚半乳糖合成的影響
在酶催化過程中為了保證底物高效利用以及產物高得率,酶用量適當是關鍵因素。S62β-半乳糖苷酶加酶量的實驗結果如圖5所示。
由圖5可知:當加酶量為15 U/mL時,加酶量低,轉化效率不高,乳糖消耗率和低聚半乳糖產率都最低;當加酶量為75 U/mL時,加酶量高,在6 h內反應速度過快,乳糖消耗率最高,但低聚半乳糖產率降低,此時低聚半乳糖更多的被降解。所以在此反應體系中,S62β-半乳糖苷酶最適加酶量為60 U/mL,此時乳糖消耗率為(82.51±0.79)%,GOS轉化率為(33.88±0.01)%。
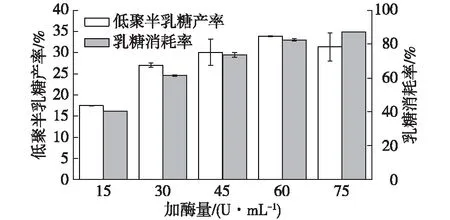
圖5 加酶量對低聚半乳糖合成的影響Fig.5 Effects of enzyme amount on GOS synthesis
2.6 反應時間對低聚半乳糖合成的影響
β-半乳糖苷酶反應工藝中要求的是高效高產低耗,所以反應時間的控制有利于提高工藝效率。S62β-半乳糖苷酶的反應時間對合成的影響結果如圖6所示。
由圖6可知:乳糖消耗率隨著反應時間的增加而增加,但低聚半乳糖產率先增加后降低,在反應時間為10 h時,低聚半乳糖產率達到最大值,為(33.1±0.32)%,乳糖消耗率為(77.87±0.28)%。
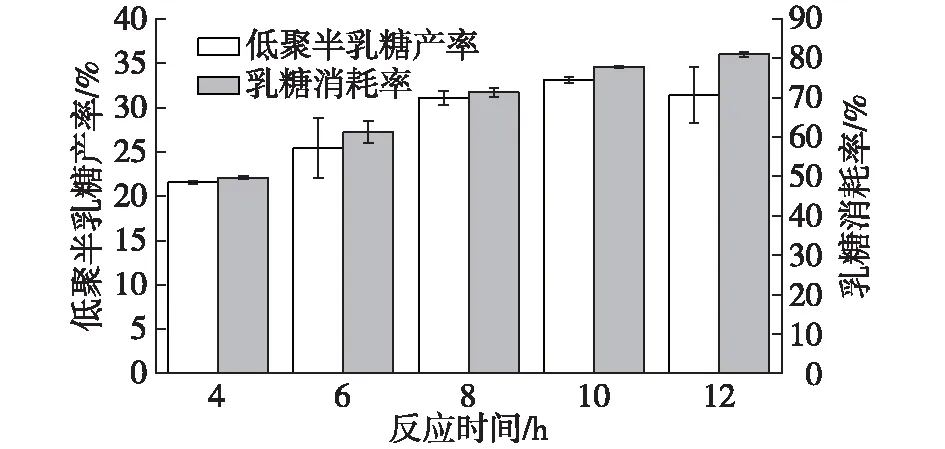
圖6 反應時間對低聚糖合成的影響Fig.6 Effects of time on the GOS synthesis
2.7 低聚半乳糖合成正交試驗
在以上的單因素條件基礎上,設定反應時間為4 h,以底物濃度、反應溫度、加酶量、pH為考察對象,設計正交試驗表如表2所示,試驗結果和結果分析如表3和表4所示。

表2 正交試驗設計表
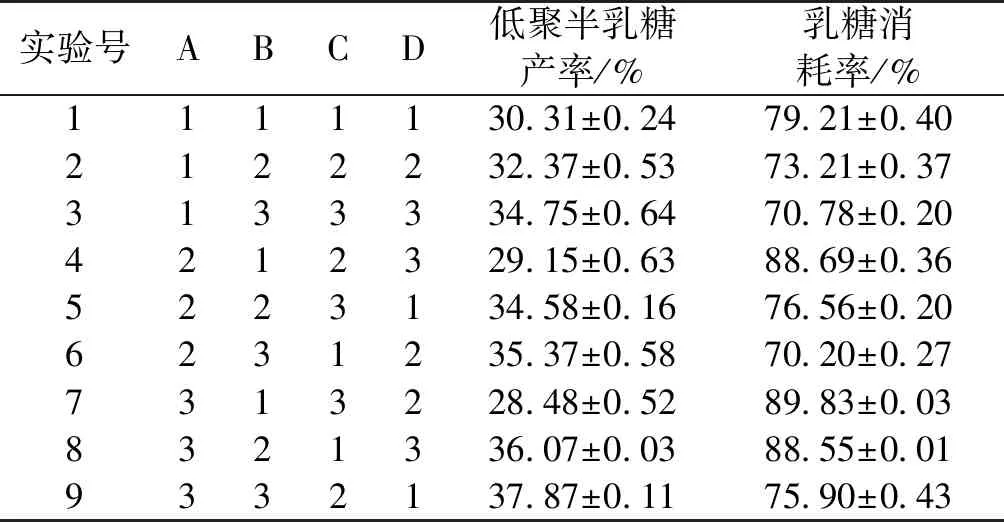
表3 正交試驗結果表
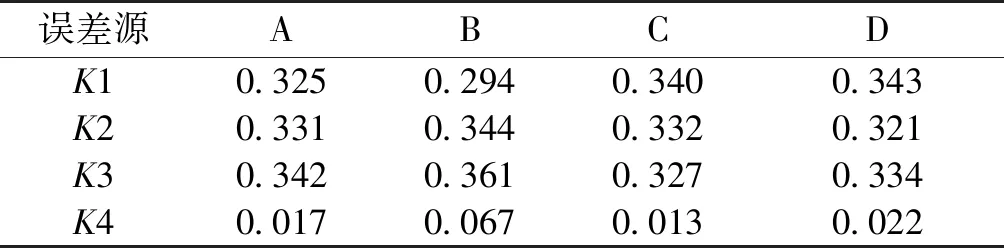
表4 結果分析表
由表2~4可知:最適反應條件為反應溫度40 ℃、底物質量濃度300 g/L、反應pH 7.0、加酶量50 U/mL,得到最高低聚半乳糖產率為(37.87±0.11)%,此時乳糖消耗率為(75.90±0.43)%。不同因素對低聚半乳糖產率的影響順序從大到小依次為底物濃度、加酶量、pH、溫度。
2.8 低聚半乳糖合成最適條件
在上述正交試驗結果的最優(yōu)反應體系下,每隔1 h取樣1次,結果如圖7所示。
由圖7可知:低聚半乳糖產率和乳糖降解率隨著反應時間的延長而增加,在4~8 h內,低聚半乳糖產率都維持在40%,在反應6 h時可獲得最大低聚半乳糖產率(41.91±0.27)%,乳糖消耗率為(82.47±0.38)%。
目前,大部分β-半乳糖苷酶用于合成低聚半乳糖時,乳糖的高效利用率和低聚半乳糖高產率無法兼得。Aburto等[11]對米曲霉β-半乳糖苷酶進行了研究,在其最優(yōu)反應體系中,低聚半乳糖產率為40%,40.3%的乳糖發(fā)生轉化,但其乳糖利用率不高。Geiger等[31]報道了嗜熱鏈球菌β-半乳糖苷酶在37 ℃、80%乳糖轉化時,低聚半乳糖產率達到總糖含量的34.2%;在94%~95%的乳糖轉化率下,低聚半乳糖產率降低至30%,乳糖利用率高,但低聚半乳糖產率低。Vasiljevic等[32]報道了保加利亞乳桿菌的β-半乳糖苷酶以質量分數30%的乳糖為底物,在50 ℃下獲得最佳的低聚半乳糖產量,為20%。而本研究中的S62β-半乳糖苷酶在乳糖轉化率接近90%時,反應4~6 h時,低聚半乳糖產率始終可保持40%以上,所以S62β-半乳糖苷酶可以在較長的時間內維持高低聚半乳糖產量。
對40 ℃下S62β-半乳糖苷酶剩余酶活進行檢測,8 h時酶活僅剩60%左右。由于S62β-半乳糖苷酶屬于低溫酶,溫度高時,酶的半衰期短,后期酶活降低,減慢了低聚半乳糖的水解,這也可能是低聚半乳糖產率在長時間內維持在40%以上的原因之一。
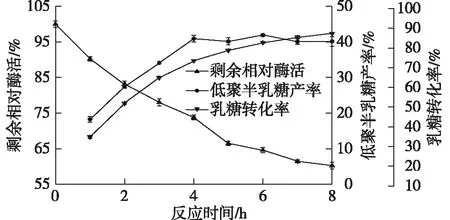
圖7 最適條件反應結果及剩余酶活變化Fig.7 Optimum reaction conditions and the remaining changes of enzyme activity
3 結論
筆者對S62β-半乳糖苷酶產低聚半乳糖工藝進行了研究,反應體系為1.9 mL反應液,0.1 mL酶液,得到最適產低聚半乳糖條件如下:反應溫度40 ℃、底物質量濃度300 g/L、反應pH 7.0、加酶量50 U/mL,反應6 h時可獲得最大低聚半乳糖產率(41.91±0.27)%,此時乳糖消耗率為(82.47±0.38)%。反應4~8 h內,低聚半乳糖產率均維持在40%以上。而在本實驗中,當乳糖轉化率接近90%時,低聚半乳糖產率仍可維持在40%左右,即乳糖轉化率與低聚半乳糖產率均可維持在較高水平,不僅獲得了高附加值的產物,也在一定程度上提高了原料的利用率,這可為后續(xù)的S62β-半乳糖苷酶工藝應用提供技術參考,同時也有利于低聚半乳糖的工業(yè)化生產。
