光合-厭氧混合菌群生物共發酵產氫動力學研究
2019-04-01 12:54:12杜金宇任學勇青春耀荊艷艷李林澤
農業機械學報 2019年3期
杜金宇 任學勇 青春耀 荊艷艷 李林澤 王 毅
(1.河南牧業經濟學院能源與動力工程學院, 鄭州 450011; 2.北京林業大學材料科學與技術學院, 北京 100083;3.河南農業大學農業農村部農村可再生能源新材料與裝備重點實驗室, 鄭州 450002)
0 引言
氫能因其清潔、高效、可再生的特點,是一種理想的替代能源。生物制氫能將氫能生產與廢棄物利用、太陽能轉化和污染治理等相結合,具有廣闊的應用前景[1-2]。目前,主要有兩種研究較多的生物制氫方法,一是厭氧細菌暗發酵制氫,能夠利用廉價而大量的有機廢棄物為原料進行制氫,制氫成本低,產氫速率快,但是暗發酵產氫效率較低[3-5]。二是光合細菌光發酵制氫,可以利用暗發酵產氫的代謝產物進行產氫,產氫效率高,但其底物降解能力低,產氫速率慢[6]。為了克服厭氧細菌暗發酵制氫和光合細菌光發酵制氫各自的缺點,根據其代謝過程互補的性質,將暗發酵和光發酵聯合起來進行聯合制氫可以有效提高制氫效率[7-8]。
越來越多的研究者開始研究將光合細菌和厭氧細菌共同培養建立暗發酵和光發酵同時進行的共發酵產氫體系。FANG等[9]研究發現將厭氧細菌Clostridiumbutyricum和光合細菌Rhodobactersphaeroides共同培養利用葡萄糖為底物的產氫量比單獨的暗發酵或光發酵的產氫量大。MIYAKE等[10]將厭氧細菌C.butyricum和光合細菌突變株共同培養產氫,發現兩種細菌混合培養的產氫量從1.1 mol/mol上升到7.0 mol/mol(以葡萄糖為底物計)。 ASADA等[11]報道了將Lactobacillus和R.sphaeroidesRV固定化混合培養產氫時,產氫量可以達到7.1 mol/mol。發展兩種或多種細菌共存的共發酵產氫體系對于提高生物制氫效率、實現生物制氫規模化應用具有重要意義。
生物制氫最重要的目標就是實現制氫過程優化和控制,以實現規模化綠色生物氫能生產,而制氫過程數學模型的建立則是實現制氫過程優化控制的重要手段[12]。生物制氫動力學的研究以及數學模型的建立和優化能夠更好地認識產氫微生物制氫過程中菌體的生長和產物的形成機制,能夠有效分析和預測生物制氫的產量和過程控制,從而顯著提高生物制氫效率。LUIS等[13]建立了一個連續流生物制氫系統的數學模型,OBEID等[14]建立了一個沼澤紅甲單胞菌批式光發酵生物制氫的動力學模型,ZHANG等[15]采用GMOPERZ模型分析了光合細菌混合菌群的光發酵制氫。但是這些研究都是集中于利用現有的模型對生物制氫進行分析和預測,而且只針對產氫量進行數學模型的分析和動力學參數的計算,沒有將生物發酵制氫過程中菌體生長、底物消耗以及產氫量統一進行考慮,生物制氫過程的生長動力學、基質消耗動力學以及產氫動力學數學模型的研究還未展開[16]。
本文在研究光合、厭氧混合菌群共發酵產氫特性的基礎上,建立混合菌群生物共發酵產氫過程的菌體生長動力學、底物降解動力學和產氫力學模型,分析數學模型中動力學參數變化對共發酵產氫的影響,對數學模型動力學參數的敏感性進行分析。目的在于建立光合、厭氧混合菌群生物共發酵產氫的動力學模型,為生物制氫反應器的運行和優化及生物氫能的實際應用提供參考和依據。
1 實驗
1.1 實驗材料
1.1.1混合菌群
實驗使用的光合、厭氧混合產氫菌群由河南農業大學農業農村部農村可再生能源新材料與裝備重點實驗室提供,混合菌群篩選過程參見文獻[12,17]。該菌群包含5種光合細菌、11種厭氧細菌。
1.1.2培養基
(1)生長培養基:NH4Cl質量濃度0.1 g/L、NaHCO3質量濃度0.2 g/L、K2HPO4質量濃度0.02 g/L、CH3COONa質量濃度0.3 g/L、MgSO4·7H2O質量濃度0.02 g/L、NaCl質量濃度0.2 g/L、酵母膏質量濃度0.1 g/L。培養基在0.1 MPa下滅菌20 min。
(2)產氫培養基:NH4Cl質量濃度0.4 g/L;MgCl2質量濃度0.2 g/L;酵母膏質量濃度0.1 g/L;K2HPO4質量濃度0.5 g/L;NaCl質量濃度2 g/L;谷氨酸鈉質量濃度3.5 g/L,葡萄糖質量濃度30 g/L。
1.2 實驗方法
1.2.1產氫實驗
產氫反應器采用300 mL廣口瓶,加入240 mL產氫培養基,在121℃條件下滅菌處理15 min,冷卻后接種60 mL(接種量20%)菌種,頂部空間充入氬氣,用膠塞密封,石蠟封口。培養溫度30℃,利用60 W白熾燈光照,反應器外表面的光照強度為150 W/m2,發酵產生的氣體用排水集氣法收集,記錄產氣量[18]。
1.2.2測試方法
(1)氫氣濃度
氫氣濃度采用安捷倫6820GC-14B型氣相色譜儀進行測定。色譜條件:進樣口溫度100℃,柱溫80℃,TCD檢測器150℃,進樣量500 μL,保留時間2 min,填充有5A分子篩的1 m柱,載氣為氬氣[19]。
(2)菌體生物量
菌體細胞濃度采用光電比濁法測定,使用721型分光光度計測量菌體細胞在660 nm處的吸光度,對照吸光度與菌體濃度的標準曲線獲得菌體濃度。
(3)底物濃度
產氫底物葡萄糖濃度測試采用DNS法測定,利用分光光度計在540 nm波長下測定其OD(吸光度)值,對照標準曲線獲得葡萄糖濃度[20]。
2 數學模型
2.1 模型建立
在批式發酵產氫實驗中,菌體生物量和產氫底物質量濃度與時間的關系表示[21-22]為
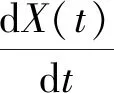
(1)
(2)
其中
式中X——菌體質量濃度
S——底物質量濃度
μ(S)——比生長速率
μmax——最大比生長速率
KS——飽和常數
YX/S——底物利用率系數
t——時間
產氫速率既與產氫菌體質量濃度直接相關,又與光照強度相關。依賴于光照強度的產氫動力學模型可以表示為
(3)
式中VH2——產氫量
YX/P——氫得率系數
β——非生長性產物得率系數
φ(I)——光照強度
當t=0時,令X(0)=X0,S(0)=S0,VH2(0)=VH20。
通過式(1)和式(2)可以得到
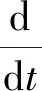
(4)
對式(4)求解可得
(5)

2.2 模型求解
近年來,非線性方程的求解已經有非常深入的研究,研究較多的有同倫攝動法(Homotopy perturbation method)、同倫分析法(Homotopy analysis method)、變分迭代法(Variational iteration method)等[24-25]。同倫攝動法(HPM)因其在零次迭代時能夠獲得簡單的近似解,因此受到廣泛應用,本文采用同倫攝動方法對式(1)~(3)非線性微分方程進行求解[26]。菌體質量濃度表示為
(6)
其中
XSS=X0+S0YX/S
將式(6)代入式(4),底物質量濃度表示為
(7)
(8)
式中VH2SS——穩定期產氫量
VH20——起始產氫量
產氫量VH2(t)代入式(1)、(3)、(6)得到
(9)
式(6)、(7)、(9)分別為生物量、底物質量濃度和產氫量的數學表達式,將式(6)、(7)代入式(9)中,可以得到產氫量與底物質量濃度的關系
(10)
3 結果與分析
3.1 動力學參數
動力學參數由實驗數據獲得。其中μmax=0.3 g/L,KS=10 g/L,YX/S=0.7 g/L,YX/P=0.7 g/L,β=12 g/L,φ(I)=3 000 lx,X0=0.2 g/L,S0=4.2 g/L,VH20=0 g/L。
式(6)~(9)方程的解析解通過同倫攝動法(HPM)進行求解,圖1對實驗數據獲得的菌體質量濃度、底物質量濃度、產氫量和動力學模型進行了對比,模型和實驗數據能夠很好地保持一致,能夠很好地反映產氫過程中菌體質量濃度、底物質量濃度及產氫量的變化情況。

圖1 動力學模型和實驗數據的對比Fig.1 Comparison of experimental results and kinetics models
3.2 敏感性分析
為了進一步優化動力學模型,有效預測產氫過程中參數的動態變化,對模型的敏感性進行了分析,菌體質量濃度X隨μmax、KS和YX/S的變化量分別為79%、-2%和23%。底物濃度S隨μmax、KS和YX/S的變化量分別為118%、-15%和-3%。產氫量VH2隨μmax、φ(I)和YX/P的變化量分別為98.4%、2%和-0.6%。本研究中最大比生長速率μmax的變化最為重要,因為μmax在菌體質量濃度、底物質量濃度以及產氫量的模型中都直接相關。
圖2a~2c分別給出了菌體質量濃度X(t)關于最大比生長速率μmax、底物利用率系數YX/S和飽和常數KS隨時間t的變化趨勢。從圖2a可以看出,最大比生長速率μmax越大,菌體質量濃度X(t)到達穩定期的時間越短。當μmax非常小時,菌體質量濃度達到穩定期的時間會變得很大。圖2b給出了底物利用率系數的變化對菌體質量濃度的影響,可以看出,菌體質量濃度X(t)隨著底物利用率系數YX/S的增大而增大,這是因為菌體穩定期濃度XSS取決于底物利用率系數YX/S(XSS=X0+S0YX/S)。在各種YX/S情況下,菌體質量濃度在t=80 h時達到最大值。圖2c表明,飽和常數KS的變化對菌體質量濃度X(t)的影響相對于μmax和YX/S并不顯著。

圖2 菌體質量濃度隨不同動力學參數的變化曲線Fig.2 Changing curves of concentration of biomass with time under different kinetic parameters
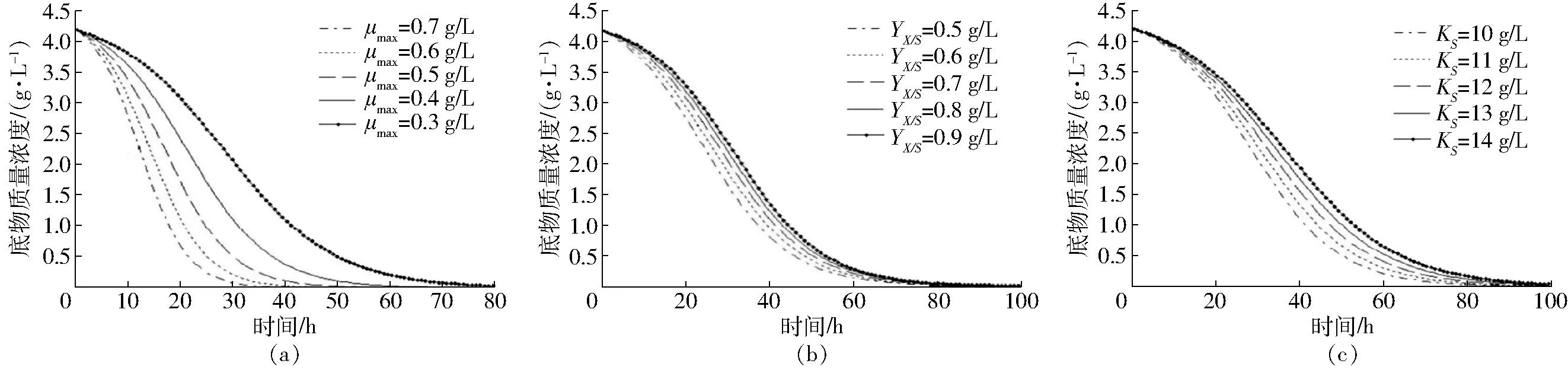
圖3 底物質量濃度隨不同動力學參數的變化曲線Fig.3 Changing curves of concentration of substrate with time under different kinetic parameters
圖3給出了動力學參數的變化對底物質量濃度的影響,從圖3a可以看出,底物質量濃度S(t)隨著最大比生長速率μmax的增加而迅速降低,很快達到穩定期底物質量濃度SSS=0。從圖3b和圖3c可以看出,隨著底物利用率系數YX/S和飽和常數KS的增大,底物質量濃度會增大。
圖4給出了最大比生長率μmax、光照強度φ(I)和非生長性產物得率系數β的變化對產氫量VH2(t)的影響。從圖4a可以看出,隨著最大生長速率μmax的增高,產氫量VH2(t)也在提高,從圖4b可以看出,產氫量VH2(t)和非生長性產物得率系數β也直接相關,VH2(t)隨著β的增大而提高。圖4c給出了光照強度φ(I)的變化對產氫量VH2(t)的影響,可以看出在其他動力學參數一定的情況下,改變光照強度φ(I)并不會引起產氫量VH2(t)的變化,這是因為光照強度對光合細菌的影響主要體現在光照強度較小的范圍內,即使當光照強度超過8 000 lx(圖中未列出)時,增加光照強度也不能提高產氫量,該結果與張全國等[27]的研究一致。在其他動力學參數一定的情況下,光照強度對產氫量的影響并不顯著,混合菌群生物共發酵產氫過程中,產氫量主要取決于菌體的最大比生長速率,與菌體的生長直接相關,光照強度的影響并不大。
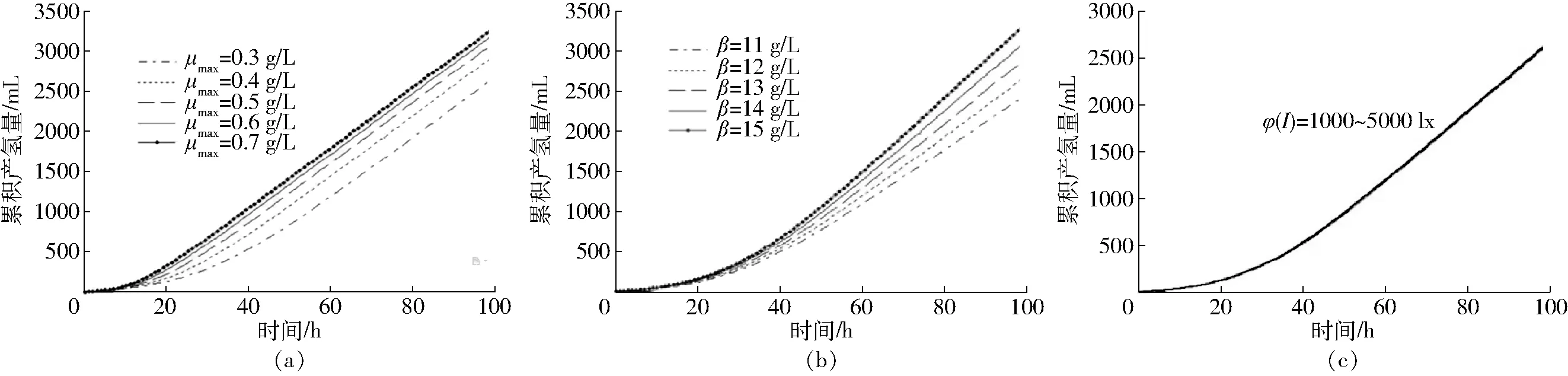
圖4 產氫量隨不同動力學參數的變化曲線Fig.4 Changing curves of hydrogen production of substrate with time under different kinetic parameters
4 結論
(1)采用同倫攝動法(HPM)對非線性動力學模型進行求解,得到了混合菌群共發酵產氫過程中菌體濃度、底物利用及產氫量的動力學模型,以及共發酵產氫過程中產氫量對于菌體質量濃度、底物質量濃度的動力學模型,建立了產氫量和菌體生長及底物消耗的關系。
(2)通過與實驗數據對比,建立的動力學模型與實驗數據基本一致,能夠很好地反映出共發酵產氫過程中菌體質量濃度、底物質量濃度、產氫量等的變化趨勢。
(3)動力學參數中最大比生長速率對模型結果的影響最大,最大比生長速率對菌體質量濃度影響的變化量達到79%,對底物質量濃度影響的變化量達到118%,對產氫量影響的變化量達到98.4%。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
