接種比例和接種物馴化對榨汁橙渣厭氧發(fā)酵的影響
2019-04-01 12:56:06張衍林
農業(yè)機械學報 2019年3期
席 江 艾 平 袁 蕭 龍 燕 張衍林
(1.華中農業(yè)大學工學院, 武漢 430070; 2.農業(yè)農村部沼氣科學研究所, 成都 610041)
0 引言
2016年全球橙種植面積超過3.96×106hm2,總產量超過7×107t,而中國橙產量位于全球第二,產量超過8×106t,占全球橙產量的11.5%[1]。全球30%~50%的柑橘被用于鮮食,剩余部分被用于加工,主要生產果汁、果醬、果膠和其他食品[2],柑橘加工后殘渣的質量達到了鮮果的50%~60%,主要包含皮、種子和果肉殘渣[3]。我國每年會產生超過5×106t柑橘廢渣,除去一小部分被用于果膠的提取和動物飼養(yǎng),絕大多數廢渣被直接丟棄或填埋[4]。榨汁橙渣富含果膠等有機質,適宜采用厭氧生物降解手段進行處理,既環(huán)保,又能回收能源,發(fā)酵之后的沼渣還可以作為一種土壤改良劑,替代部分化肥的使用,促進作物生長,緩解土壤板結[5]。
榨汁橙渣易酸化的特性和橙皮中含有對微生物有抑制作用的精油常會導致厭氧發(fā)酵不正常或產氣率較低。FAGBOHUNGBE等[6]采用3~3.26的接種比例(接種物與原料的揮發(fā)性固體質量比,Inoculum and substrate ratio, ISR)開展橙皮的厭氧發(fā)酵,盡管加入了生物炭,但得到最高甲烷累積產率僅154 mL/g,發(fā)酵過程中均出現了不同程度的停滯。SANJAYA等[7]研究了中溫條件下,橙的種子、果皮和果肉的甲烷累積產率,選擇的ISR為4,橙皮的甲烷累積產率只有48.23 mL/g,研究認為是因精油中檸檬烯等一些有芳香味的化合物對厭氧發(fā)酵產生了抑制,可能使用的接種物對橙皮適應性較差而同時添加量又不足,才造成厭氧發(fā)酵異常和產氣率較低。WIKANDARI等[8]在30 mL厭氧發(fā)酵體系中添加20 mL接種物,仍只得到131 mL/g的甲烷累積產率,說明以這類廢棄物為發(fā)酵原料可能需要更高的ISR。還有一些研究通過水蒸氣蒸餾[3]、溶劑萃取[8]、汽爆[9]和生物降解[4]等預處理手段以促進橙皮的厭氧發(fā)酵。FORGCS等[9]通過汽爆預處理柑橘榨汁渣,使甲烷累積產率提高了426%, 但預處理前的甲烷累積產率僅102 mL/g,可能是因原料添加量較高(接種物添加較少)而導致預處理前甲烷累積產率較低。一些其他的預處理如生物預處理之后甲烷累積產率也只有176.05 mL/g[4],且沒有通過優(yōu)化厭氧發(fā)酵體系來充分挖掘產甲烷潛力。對于榨汁橙渣厭氧發(fā)酵的研究,多集中于不同預處理的優(yōu)化,這些預處理均需要額外的能耗或化學試劑添加,而不同接種比例和接種物馴化對榨汁橙渣厭氧發(fā)酵影響的研究相對較少。
因此,本文以榨汁橙渣為厭氧發(fā)酵原料,通過監(jiān)測不同ISR條件下榨汁橙渣厭氧發(fā)酵產甲烷量、pH值和揮發(fā)酸含量,比較接種物和發(fā)酵結束后厭氧發(fā)酵體系中細菌和古菌群落組成的變化,系統分析榨汁橙渣厭氧發(fā)酵的規(guī)律,同時,通過監(jiān)測馴化后接種物的厭氧發(fā)酵甲烷產氣量,分析通過馴化接種物促進榨汁橙渣厭氧發(fā)酵的有效性和可行性,為榨汁橙渣厭氧發(fā)酵及其預處理優(yōu)化研究提供參考。
1 材料與方法
1.1 實驗材料與接種物
實驗用榨汁橙渣(Orange pressing waste, OPR)取自“天使之橙”鮮榨橙汁自助販賣機,手動分選出橙皮渣(Orange peel waste, OPW)和橙肉渣(Orange pulp waste, OPU),OPW與OPU鮮質量比為2.39∶1,經粉碎和過10目篩后,抽真空-20℃分別保存?zhèn)溆谩=臃N物取自四川省德陽市正常運行的集中供氣工程,該工程以豬糞為發(fā)酵原料。接種物取回后裝入5 L厭氧發(fā)酵瓶中,35℃水浴放置一段時間直到不再產氣為止。OPW、OPU和接種物的總固體(Total solid, TS)、揮發(fā)性固體(Volatile solid, VS)和檸檬烯含量見表1。
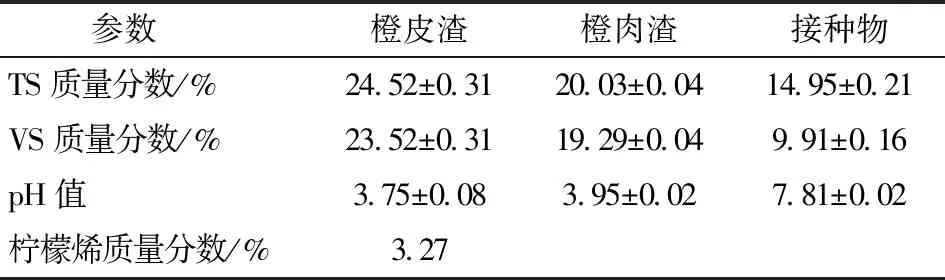
表1 榨汁橙皮渣、橙肉渣和接種物理化特性Tab.1 Characteristics of OPW, OPU and inoculum
驗證實驗的發(fā)酵原料為夏橙榨汁渣,該夏橙產自湖北省宜昌市秭歸縣,手動剝下橙皮,橙肉渣為橙肉經飛利浦HR871/00型榨汁機榨汁后的殘渣,夏橙橙皮渣和橙肉渣VS質量分數分別為19.85%和13.94%,OPW與OPU鮮質量比為1.97∶1,經粉碎和10目篩過篩后,抽真空-20℃分別保存?zhèn)溆谩?/p>
1.2 厭氧發(fā)酵實驗
厭氧發(fā)酵裝置采用全自動甲烷潛力測試系統(Bioprocess AMPTSII),發(fā)酵瓶容積為400 mL,沼氣通過3 mol/L NaOH溶液吸收后進入流量自動記錄裝置,裝置將甲烷產氣量自動換算成標準狀態(tài)并記錄,厭氧發(fā)酵裝置示意圖見圖1。每隔1 d取4 mL發(fā)酵液用于pH值和揮發(fā)酸(Volatile fatty acids, VFAs)測量。厭氧發(fā)酵溫度為35℃,ISR設置見表2,每個處理設置2個重復。采用修正的Gompertz 模型描述厭氧發(fā)酵過程[10],模型公式為
式中p——t時刻的單位揮發(fā)性固體累積甲烷產量,mL/g
p0——單位揮發(fā)性固體最大產甲烷潛力,mL/g
Rmax——單位揮發(fā)性固體最大產甲烷速率,mL/(g·d)
λ——停滯期,d
t——實驗持續(xù)時間,d
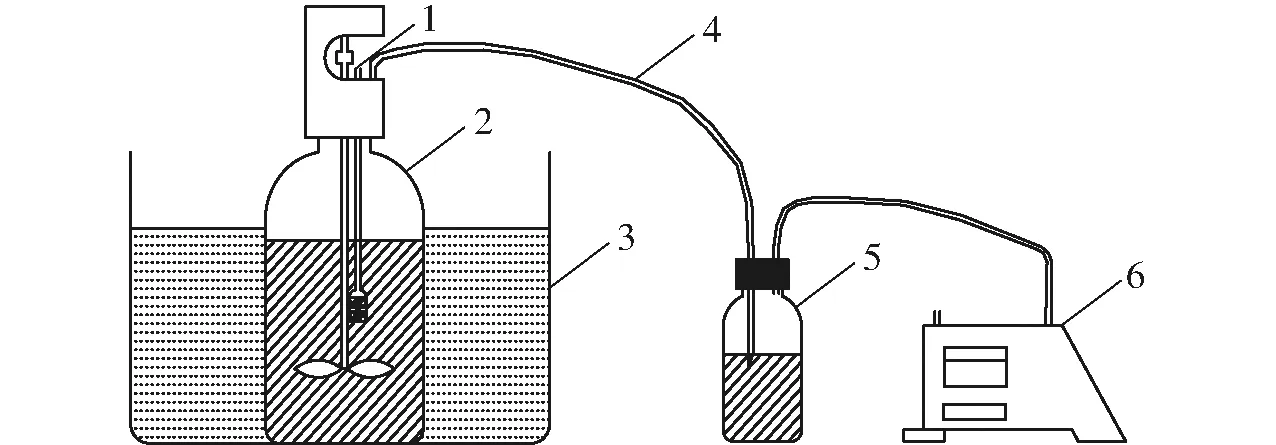
圖1 厭氧發(fā)酵裝置示意圖Fig.1 Experimental sketch of anerobic digestion device1.液樣取樣口 2.厭氧發(fā)酵瓶 3.水浴鍋 4.導氣管 5.CO2吸收瓶 6.流量自動記錄裝置
驗證實驗采用相同厭氧發(fā)酵裝置,ISR分別為8、6、4和2,每個處理設置2個重復,固定發(fā)酵原料總VS質量為2 g。
1.3 參數測量方法
TS和VS含量采用差重法測量,取10 g樣品,TS測量在(105±5)℃干燥24 h,VS含量測量在550~600℃馬弗爐中灼燒4 h; pH值采用梅特勒 FE28 pH計進行測量; VFAs含量的測量使用島津GC-2030型氣相色譜儀(日本),檢測器為火焰離子化檢測器,色譜柱為IntertCap WAX型毛細管柱(30 m×0.25 mm×0.25 μm),載氣為氦氣,進樣口溫度200℃,柱箱溫度95℃保持2 min,以10℃/min升溫至160℃,繼續(xù)以400℃/min升溫至240℃,保持5 min。檸檬烯含量測量使用島津GCMS-TQ8050型三重四極桿氣相色譜質譜聯用儀,模式選擇Q3Scan,色譜柱使用Rtx-5MS型毛細管柱(30 m×0.25 mm×0.25 μm),進樣口溫度250℃,柱溫箱溫度50℃保持1 min,以10℃/min的速率上升到280℃,保持5 min,離子源溫度200℃。
1.4 細菌和古菌多樣性分析
取樣品10 mL,于-80℃下保存。使用E.Z.N.A.soil試劑盒(美國Omega Bio-tek公司)進行總DNA抽提,DNA濃度和純度利用NanoDrop2000型紫外可見光分光光度計進行檢測,利用1%瓊脂糖凝膠電泳檢測DNA提取質,檢測合格后用引物Arch344F(5′-ACGGGGYGCAGCAGGCGCGA-3′)和Arch915R(5′-GTGCTCCCCCGCCAATTCCT-3′),338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)分別進行PCR擴增,利用Illumina公司的Miseq PE300平臺進行測序,數據進行OUT抽平后進行多樣性分析。
2 結果與討論
2.1 不同ISR對厭氧發(fā)酵產甲烷規(guī)律的影響
2.1.1甲烷日產氣率
ISR為8、6、4的處理表現出相似的產甲烷規(guī)律,見圖2。第1天達到第1個產氣高峰之后產氣量下降,隨后上升達到第2個產氣高峰,且第2個產氣高峰的日產氣率高于第1個產氣高峰。但在第1個產氣高峰之后的產氣下降幅度和時長有所區(qū)別,ISR為8、6、4分別在第3、3、4天達到甲烷日產氣率相對低點,分別為28.15、21.05、4.58 mL/g,接種比例高的先達到相對低點,同時甲烷日產氣率也較高,隨后分別在第5、6、9天分別達到第2個產甲烷高峰,分別為52.9、49.8、41.9 mL/g,接種比例高的先達到第2個產氣高峰,且甲烷日產氣率也較高。ISR為2的處理發(fā)生了可逆的酸化,在第1天達到第1個產氣高峰之后,產氣迅速停滯,直到第8天才恢復產氣,并在第10天達到第2個產氣高峰,甲烷日產氣率達到23.48 mL/g。這種現象反映了產酸和產甲烷速率的失衡,產酸產生的VFAs積累又導致pH值的下降,進一步抑制產甲烷過程[11],在這個ISR下厭氧發(fā)酵不能正常進行。
2.1.2甲烷累積產率
從圖2可以看出,ISR為8和6的甲烷累積產率相近,分別為320.0 mL/g和304.9 mL/g,說明不能通過進一步增加接種物量的方式來提高原料產氣率。這個數據與CALABRO等[12]、RUIZ等[5]和NEGRO等[13]分別報道的361、357.3、285 mL/g相近,結果略低于KAPRAJU等[14]報道的490 mL/g,這種差異可能是原料理化特性差異造成的。但實驗結果高于FORGCS等[9]報道的102 mL/g和FAGBOHUNGBE等[6]報道的165.9 mL/g,這可能是由于他們添加的接種物量不足而導致原料沒有被充分利用。
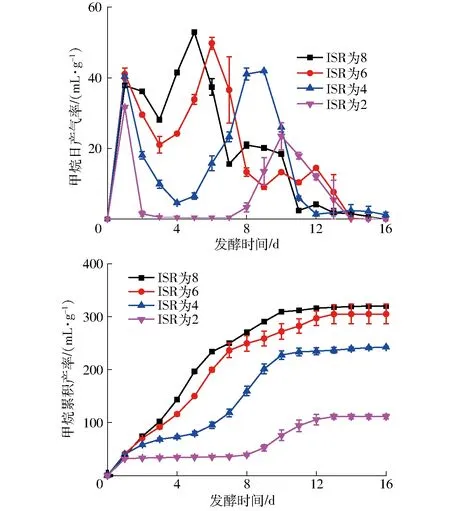
圖2 ISR為2~8的甲烷日產氣率和累積產率Fig.2 Daily and cumulative methane production of ISR 2~8
ISR為4和ISR為2的甲烷累積產率低于ISR為8和ISR為6時的數值。ISR為4的甲烷累積產率為242.6 mL/g,是ISR為8和6的甲烷累積產率的75.8%和79.6%。而ISR為2的甲烷累積產率僅為111.4 mL/g,是ISR為8和6的甲烷累積產率的34.8%和36.5%。從累積產甲烷的曲線可以看出,第1天之后厭氧發(fā)酵進入停滯階段,尤其是ISR為2時的停滯期長達7 d。
根據表1和表2可以計算出添加到厭氧發(fā)酵體系中檸檬烯的質量濃度為490 mg/L,這個濃度在中溫條件下并不會對厭氧發(fā)酵體系產生顯著抑制[12], ISR為4和2的處理的累積產甲烷量低,主要是因為接種物量較少,不能充分利用底物。
2.1.3pH值和VFAs含量
厭氧發(fā)酵系統pH值的變化與VFAs(乙酸、丙酸、丁酸和異丁酸含量之和)積累呈負相關(圖3)。ISR為8、6、4的處理在第2天揮發(fā)酸質量濃度達到峰值,分別為1 445.89、1 806.05、2 067.37 mg/L,隨接種比例增加,揮發(fā)酸質量濃度峰值降低,在第10天已經檢測不出揮發(fā)酸。ISR為2的處理是在第4天揮發(fā)酸質量濃度才達到峰值2 201.92 mg/L,呈先上升再下降的規(guī)律,在第10天揮發(fā)酸質量濃度仍達到279.24 mg/L,直到第12天才檢測不出揮發(fā)酸。同時,隨著VFAs含量的減少,pH值在逐漸上升。產甲烷的最適宜pH值是6.8~8.5[15], ISR為8和6的處理pH值一直高于7。ISR為4的處理在第2天pH值低于6.8,此后逐步恢復到7以上。而ISR為2 的處理在第6天之前pH值也一直維持在6.8以下,直到第8天 pH值才恢復到6.8以上,同時產甲烷也恢復。由于榨汁橙渣pH值在3~4[3],其低pH值和易酸化特性是影響厭氧發(fā)酵效果的一個重要因素,通過增加接種量可以有效提高發(fā)酵系統的緩沖能力,防止發(fā)酵過程中pH值大幅下降。
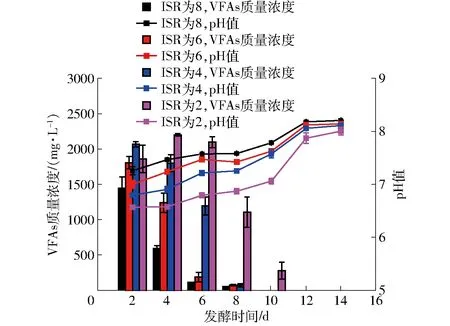
圖3 ISR為2~8 時pH值和揮發(fā)酸含量變化圖Fig.3 Changes of pH and VFAs of ISR 2~8
2.2 不同ISR對微生物多樣性的影響
2.2.1細菌群落組成
由圖4可知,接種物中主要是硬壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)和變形菌門(Proteobacteria),相對豐度分別達到86.96%、4.39%和4.48%,放線菌門(Actinobacteria)、Atribacteria和綠彎菌門(Chloroflexi)相對豐度較低,分別為2.26%、0.52%和0.32%。其他的一些研究也發(fā)現厭氧發(fā)酵系統中硬壁菌門和擬桿菌門細菌為優(yōu)勢菌群[16],放線菌門和綠彎菌門也常見于產甲烷的厭氧發(fā)酵裝置和廢水處理工程,但它們在產甲烷過程中的作用并不清楚[15]。
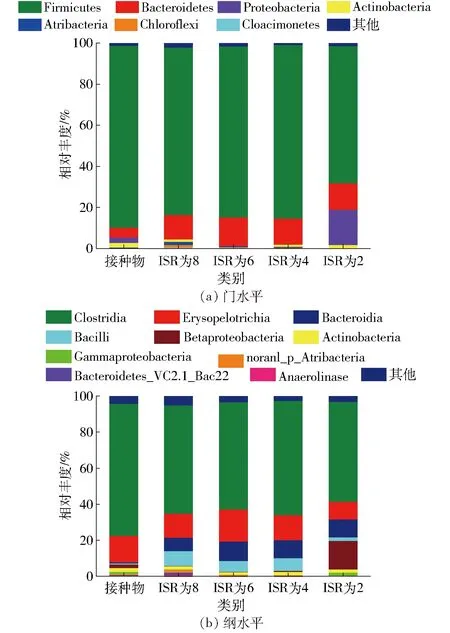
圖4 不同ISR發(fā)酵前后細菌群落組成Fig.4 Composition of bacterial community before and after anaerobic digestion
在門水平,ISR 為4、6和8的沼渣中硬壁菌門和擬桿菌門相對豐度最高,分別為79.90%~82.86%和3.33%~11.66%,LI等[17]發(fā)現這兩個門的細菌在纖維素的降解中發(fā)揮重要作用;Atribacteria和綠彎菌門相對豐度均高于接種物,但相對豐度隨ISR 下降而降低;變形菌門和放線菌門相對豐度低于接種物;Cloacimonetes門則只存在于榨汁橙渣發(fā)酵后的沼渣中,這類細菌被認為與氨基酸、丙酸、丁酸以及纖維素的降解有關[18],且相對豐度隨接種比例降低而下降。ISR為2沼渣的細菌群落組成與其他處理相比,則表現較大差異,硬壁菌門相對豐度下降到65.10%,變形菌門相對豐度上升到19.09%,綠彎菌門和Cloacimonetes門未檢測到。
在綱水平,ISR為4、6、8的沼渣中細菌群落組成相較接種物存在明顯差異,表現為梭菌綱(Clostridia)相對豐度下降到57.74%~61.7%,而同屬于硬壁菌門的芽孢桿菌綱(Bacilli)相對豐度增加到6.06%~8.20%,擬桿菌綱(Bacteroidia)相對豐度則增加到9.65%~13.00%。梭菌綱和擬桿菌綱的細菌被認為主要參與水解和產酸過程[19]。同樣地,ISR 為2沼渣中芽孢桿菌綱相對豐度低于其他處理,僅1.88%,這可能導致底物水解產酸過程受影響,但β-變形菌綱(Betaproteobacteria)豐度高于其他處理,達到15.91%。
在門水平和綱水平,ISR為4、6和8與接種物的細菌群落均有明顯差異,可能因為接種物取自以豬糞為發(fā)酵原料的沼氣工程,豬糞和榨汁橙渣的理化特性差異導致其最適宜的細菌群落組成存在差異,而經過一輪發(fā)酵后的群落組成可能更適應榨汁橙渣的厭氧發(fā)酵。ISR 為2的沼渣在門水平和綱水平與其他處理的差異可能是因為起始接種物數量不足,發(fā)酵后期菌群已經失調。
WJD-0.75電動鏟運機是武山銅礦井下主要采掘設備之一,承擔南、北礦帶大部分礦石及廢石鏟裝任務。電纜托輥是電動鏟運機運行時實現電纜順利收放的裝置[1],但在使用過程中,由于電纜托輥磨損、軸承損壞或受到外部撞擊,則需更換托輥。在實踐更換時,維修難度大、維修時間長,特別是更換水平托輥時,還需多人配合才能完成,影響井下作業(yè)點正常生產組織。因此,通過對電動鏟運機電纜托輥安裝結構進行分析,提出了托輥支架的結構改進措施。
2.2.2古菌群落組成
由圖5可知,接種物中主要存在4個屬的產甲烷菌,分別是Methanosaeta、Methanosarcina、Methanospirillum和Methanobacterium,其相對豐度分別為72.58%、5.15%、16.06%和1.73%。其中,Methanosaeta和Methanosarcina同屬于Methanosarcinales目,前者為專性乙酸營養(yǎng)性產甲烷古菌,后者則可以通過CO2還原、甲基裂解和乙酸發(fā)酵途徑產甲烷[20],而2/3甲烷的產量是來自乙酸的裂解[21];Methanospirillum和Methanobacterium分屬Methanomicrobiales和Methanobacteriales目,均為氫營養(yǎng)型產甲烷古菌[20]。發(fā)酵后古菌群落組成與接種物相比已經有所變化,在接種物中Methanospirillum的相對豐度較高,而Methanosarcina相對豐度較低,可能與發(fā)酵原料以及發(fā)酵系統中pH值有關,因為接種物取自以豬糞為發(fā)酵原料的沼氣工程,豬糞的pH值較橙榨汁渣高,而變化后的古菌群落組成可能更適應榨汁橙渣的厭氧發(fā)酵。

圖5 不同接種物量發(fā)酵前后古菌群落組成Fig.5 Composition of archaea community before and after anaerobic digestion
總體上看,Methanosaeta和Methanosarcina乙酸營養(yǎng)性產甲烷古菌的累積相對豐度隨ISR的降低而提高,ISR為2條件下,這兩個屬的產甲烷古菌相對豐度達到90.17%。這與厭氧發(fā)酵系統中揮發(fā)酸的積累隨ISR降低而提高的趨勢一致,而Methanospirillum和Methanobacterium氫營養(yǎng)型產甲烷古菌的相對豐度隨ISR降低而下降,ISR為2條件下,這兩個屬的產甲烷古菌相對豐度合計僅為6.92%。這可能與底物被大量轉化為揮發(fā)酸,而這類古菌可利用底物量下降有關。
單獨來看,乙酸營養(yǎng)性產甲烷古菌中Methanosaeta相對豐度隨ISR降低而降低,Methanosarcina相對豐度隨ISR降低而提高,在ISR為2條件下,相對豐度分別為24.45%和66.72%。這可能是因為前者最適生長的pH值范圍(7.0~7.3)較后者(6.5~7.8)更窄[22],而ISR為2的最低pH值達到6.57,不適宜Methanosaeta生長,而Methanosarcina仍能適應。同樣地,氫營養(yǎng)型產甲烷古菌中Methanospirillum相對豐度隨ISR降低而降低,Methanobacterium相對豐度隨ISR降低而提高,在ISR為2條件下,相對豐度分別為1.71%和5.21%。這可能是因為前者的最適生長的pH值范圍(7.0~7.8)較后者(5.6~8.6)更窄[22]。
2.3 接種物馴化對厭氧發(fā)酵的影響
厭氧發(fā)酵體系中微生物群落組成經過一輪厭氧發(fā)酵,已經得到了優(yōu)化,這種改變的驅動力是發(fā)酵原料特性及厭氧發(fā)酵的條件[5]。為了進一步研究接種物馴化后對榨汁橙渣厭氧發(fā)酵效率的提升,待第1輪厭氧發(fā)酵停止后,在發(fā)酵裝置中加入與第1輪相同的原料量進行第2輪發(fā)酵,在第2輪厭氧發(fā)酵停止后再加入相同原料量進行第3輪發(fā)酵,監(jiān)測厭氧發(fā)酵的甲烷產量。
ISR為8和ISR為6的3輪厭氧發(fā)酵的甲烷累積產率見圖6a。接種物馴化后甲烷累積產率沒有得到進一步提高,說明馴化前的微生物數量是足夠的,原料在發(fā)酵過程中被充分降解和利用。隨著微生物對底物適應性的提高,產氣效率在逐步提高,說明經過馴化,接種物對橙榨汁渣的適應性增強。ISR為8時第3、2、1輪甲烷累積產率分別為326.6、347.0、320.0 mL/g。從累積產甲烷量來看,雖然甲烷累積產率并沒有表現出顯著差異,但第3、2、1輪在前4 d產甲烷量占整個發(fā)酵周期產氣量的95.75%、 83.78%和44.89%,通過接種物馴化,有效縮短了產氣周期,提高了產氣效率。ISR為6的第3、2、1輪甲烷累積產率分別為326.0、311.3、304.9 mL/g。雖然甲烷累積產率并沒有表現出顯著差異,但同樣地,第3、2、1輪在前4 d產甲烷量占整個發(fā)酵周期產氣量的93.40%、86.72%和38.03%,有效縮短了產氣周期,提高了產氣效率。ISR為8和6條件下,通過接種物馴化對厭氧發(fā)酵促進的效果是一致的,這也說明ISR為6下微生物數量相對榨汁橙渣量是充裕的,通過接種物馴化可以進一步提高厭氧發(fā)酵的效率,而ISR為6較ISR為8時添加較少的接種物,更具有應用價值。
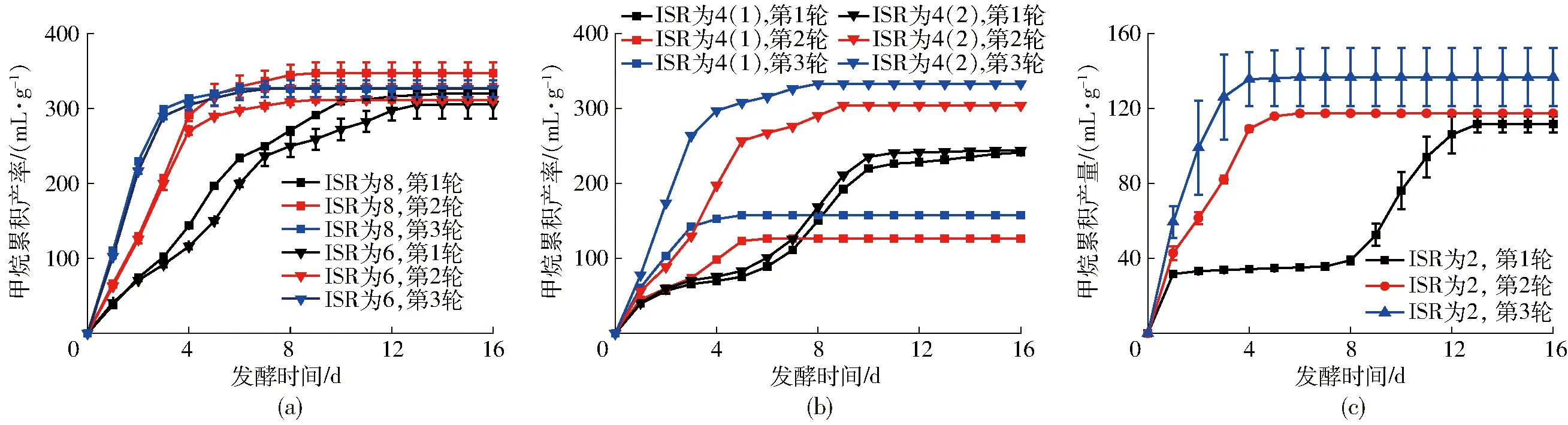
圖6 ISR為 8、6、4和2時3輪厭氧發(fā)酵甲烷累積產率Fig.6 Cumulative methane production of ISR 8, 6, 4 and 2 from the first to the third round
ISR為2的3輪厭氧發(fā)酵的甲烷累積產率見圖6c。ISR為2的處理與ISR為4的第2個重復較為相似的是對厭氧發(fā)酵的提升也不顯著,第3、2、1輪甲烷累積產率分別為136.6、117.2、111.4 mL/g。這說明在ISR為2條件下,第1輪發(fā)酵結束后微生物群落組成可能就失衡了,且為不可逆的,僅通過接種物的馴化不能促進榨汁橙渣的厭氧發(fā)酵。
采用修正Gompertz模型擬合的模型參數見表3。通過該模型可以很好地描述ISR為8和6的3輪厭氧發(fā)酵過程(R2>0.99)。從第1到第3輪,最大產甲烷速率Rmax逐步提高,反應停滯期λ在逐步縮小,其中第3輪ISR為8和ISR為6的Rmax分別達到了145.07 mL/(g·d)和139.32 mL/(g·d),而λ縮短到0.27 d和0.26 d。模型的參數也體現出ISR為8和ISR為6并沒有明顯差異,而ISR為6更具有實際應用價值。對于ISR為4和ISR為2時,修正Gompertz模型只能很好地描述部分厭氧發(fā)酵實驗(R2>0.99),見表3,其中ISR為4 的第3輪比第2輪的Rmax進一步提高,λ進一步縮小,分別達到106.91 mL/(g·d)和0.32 d,表現出與ISR為8和ISR為6時一致的規(guī)律。

表3 修正Gompertz方程預測的榨汁橙渣厭氧發(fā)酵產甲烷動力學參數Tab.3 Model estimation results of OPR by modified Gompertz models
驗證實驗以夏橙榨汁渣為發(fā)酵原料,接種物經過一輪馴化,厭氧發(fā)酵結果見表4。ISR為8和6的甲烷累積產率分別為291.0 mL/g和298.5 mL/g,高于ISR為4和2的269.7 mL/g和208.1 mL/g,且ISR為8和6的產氣周期也較ISR為4和ISR為2時短。這進一步驗證了以橙榨汁廢渣為原料開展厭氧發(fā)酵實驗,宜采用ISR為6并將接種物馴化一輪。
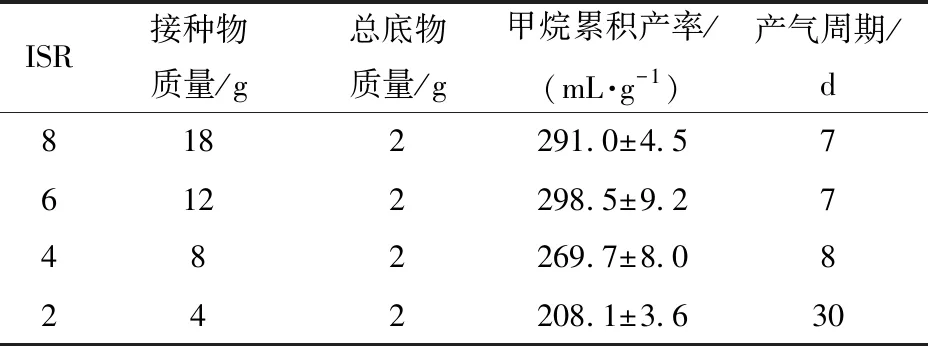
表4 夏橙榨汁渣厭氧發(fā)酵結果Tab.4 Results of anerobic digestion of XC
3 結論
(1)ISR為8和6條件下,榨汁橙渣厭氧發(fā)酵可以正常進行,甲烷累積產率分別達到320.0 mL/g和304.9 mL/g,產氣結束后發(fā)酵系統中細菌和古菌的群落組成也相似。同時,通過馴化優(yōu)化微生物群落組成雖然不能提高甲烷累積產率,但可以有效縮短發(fā)酵周期,其中,ISR為6條件下第3、2、1輪發(fā)酵前4 d產甲烷量占總產量的93.40%、86.72%和38.03%,有效提高了發(fā)酵效率。開展榨汁橙渣中抑制成分在厭氧發(fā)酵中的代謝規(guī)律以及厭氧發(fā)酵的預處理優(yōu)化等相關實驗時,推薦選擇ISR為6,并進行至少一輪接種物的馴化以避免因接種物數量和來源的差異對實驗結果帶來干擾。
(2)ISR為4條件下,接種物中微生物數量相對不足,不能充分利用底物,厭氧發(fā)酵效率低于ISR為8和6,累積甲烷產率也只有ISR為8和6的75.8%和79.6%。在該ISR下通過馴化接種物來促進厭氧發(fā)酵存在偶然性和不確定性,接種物中本身的微生物菌群落組成可能會更多地影響馴化的效果和質量。
(3)ISR為2條件下,接種物中微生物數量嚴重不足,厭氧發(fā)酵出現停滯,累積甲烷產率只有ISR為8和ISR為6的34.8%和36.5%,芽孢桿菌綱相對豐度下降到1.88%,Methanosaeta和Methanospirillum的相對豐度也分別下降到24.45%和1.71%,影響了甲烷的產生,且通過馴化接種物也無法改變微生物群落組成失調的狀況。
