三種不同需冷量桃品種休眠誘導(dǎo)期生理變化
2019-03-11 06:24:10文濱濱張新昊陳修德高東升朱翠英肖偉
文濱濱,張新昊,陳修德,2,高東升,2,朱翠英*,肖偉,2*
?
三種不同需冷量桃品種休眠誘導(dǎo)期生理變化
文濱濱1,張新昊1,陳修德1,2,高東升1,2,朱翠英1*,肖偉1,2*
1. 山東農(nóng)業(yè)大學(xué)園藝科學(xué)與工程學(xué)院, 山東 泰安 271018 2. 山東省果蔬優(yōu)質(zhì)高效生產(chǎn)協(xié)同創(chuàng)新中心, 山東 泰安 271018
以三個(gè)不同需冷量的桃品種‘常綠桃’(無(wú))、‘春捷’(102~120 C.U.)、‘青州蜜桃’(1200 C.U.)嫁接在同一桃樹(shù)砧木‘山桃’上為實(shí)驗(yàn)材料,研究不同需冷量桃葉片在自然休眠誘導(dǎo)期(8月20日-11月20日)的生理變化。結(jié)果表明:在自然休眠誘導(dǎo)期,桃葉片的抗氧化保護(hù)酶活性、光合速率、可溶性糖和淀粉含量發(fā)生顯著變化。隨著休眠誘導(dǎo)的發(fā)展,超氧化物歧化酶(SOD)活性一直升高,在休眠誘導(dǎo)期后期(10月10日)‘常綠桃’高于‘春捷’和‘青州蜜桃’,過(guò)氧化物酶(POD)活性下降,不同需冷量桃無(wú)顯著差異,過(guò)氧化氫酶(CAT)活性呈雙峰變化且常綠桃峰值低于‘春捷’和‘青州蜜桃’;凈光合速率呈下降趨勢(shì)但常綠桃高于‘春捷’和‘青州蜜桃’且峰值分別是‘春捷’和‘青州蜜桃’的1.96和3.13倍;可溶性糖含量降低、淀粉含量增加但‘常綠桃’可溶性糖含量高于‘春捷’和‘青州蜜桃’、淀粉含量低于‘春捷’和‘青州蜜桃’。以上結(jié)果表明,‘常綠桃’在休眠誘導(dǎo)期保護(hù)酶活性、光合速率和可溶性糖含量均高于‘春捷’和‘青州蜜桃’,淀粉含量低于‘春捷’和‘青州蜜桃’。
桃; 自然休眠誘導(dǎo); 生理變化; 需冷量; 葉片
落葉果樹(shù)休眠是多年生植物中非常重要的生命現(xiàn)象,是在自然進(jìn)化過(guò)程中對(duì)環(huán)境條件和季節(jié)性變化作出的一種有益的生物學(xué)適應(yīng)。自然休眠的發(fā)生與發(fā)展主要與溫度、光照等自然條件密切相關(guān),
Lang等根據(jù)植物的生長(zhǎng)特性將休眠定義為植物體內(nèi)包括任何分生組織的結(jié)構(gòu)有可見(jiàn)生長(zhǎng)的暫時(shí)停止[1]。根據(jù)休眠狀態(tài)的不同可以將休眠分為相對(duì)休眠,內(nèi)休眠和生態(tài)休眠。
在落葉果樹(shù)休眠期間,碳水化合物作為一種重要的能量來(lái)源,能夠增強(qiáng)植物的抗寒性、維持植物的正常生命活動(dòng)[2,3],其中可溶性糖含量的變化趨勢(shì)與果樹(shù)的休眠狀態(tài)密切相關(guān)[4]。葉片是植物進(jìn)行光合作用的主要部位,同時(shí)也是感應(yīng)外界環(huán)境的重要器官。李冬梅等研究發(fā)現(xiàn)葉片淀粉含量的增加和可溶性糖含量的減少標(biāo)志著果樹(shù)進(jìn)入休眠誘導(dǎo)期[5,6]。在自然休眠過(guò)程中,多個(gè)環(huán)境因子共同起作用,其中Gonzalez-Rossia的研究表明,低溫在誘導(dǎo)桃樹(shù)休眠的過(guò)程中,山梨醇和淀粉含量增加,而可溶性碳水化合物含量降低[7]。在一定溫度范圍內(nèi), 草莓葉片可溶性糖含量會(huì)隨溫度的降低與時(shí)間的延長(zhǎng)呈顯著增加趨勢(shì)[8]。所以北方落葉果樹(shù)休眠前的低溫誘導(dǎo)非常重要,將直接影響葉片光合作用同化物的積累以及養(yǎng)分回流并且直接影響翌年花芽分化的質(zhì)量以及果實(shí)的品質(zhì)[9-11]。目前,休眠過(guò)程中糖代謝的研究主要在果樹(shù)芽的自然休眠期[12]和生態(tài)休眠期[13],在休眠誘導(dǎo)期不同需冷量桃葉片糖含量的變化鮮見(jiàn)報(bào)道。
北方落葉桃樹(shù)需冷量大多在800~1000 C.U.,而本實(shí)驗(yàn)室于湖北發(fā)現(xiàn)了可以周年生長(zhǎng)且新老葉持續(xù)更替的桃樹(shù)新品種,暫時(shí)定名為‘常綠桃’。本研究通過(guò)在同一砧木上嫁接三種不同需冷量的桃,測(cè)定休眠誘導(dǎo)期不同需冷量桃葉片抗氧化保護(hù)酶活性、光合性能、可溶性糖含量、淀粉含量初步探究不同需冷量桃在休眠誘導(dǎo)期的生理變化,為研究不同需冷量桃的休眠差異奠定理論基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料與處理
試驗(yàn)在山東農(nóng)業(yè)大學(xué)科技創(chuàng)新園進(jìn)行(36°18'N,117°13' E),8月平均自然溫度為20.7~30.4℃,9月為17.3~25.8℃,10月為10.5~22.1℃,11月為2.9~13.6 ℃。選取一年生直徑1cm的山桃實(shí)生苗,2016年3月初定植,35 cm截干,4月選留3個(gè)粗細(xì)一致的枝條,5月底分別在3個(gè)枝上嫁接‘常綠桃’[(L.)Evergrowing peach]、‘春捷’[(L.)Mill]和‘青州蜜桃’[( L.)Mill]三種需冷量不同的桃品種,6月20日剪砧,相互對(duì)照,統(tǒng)一正常管理。
1.2 測(cè)定項(xiàng)目與方法
1.2.1 枝條生長(zhǎng)狀況和休眠狀態(tài)的測(cè)定8月20日起,每隔10 d選取生長(zhǎng)均勻一致且方位相同的的一年生枝條,測(cè)量從生長(zhǎng)點(diǎn)到枝條基部的長(zhǎng)度,3次生物學(xué)重復(fù)。取樹(shù)冠外圍(相同方位和高度)10根生長(zhǎng)健壯的1年生枝條,然后去掉葉片,剪去頂端2~3 cm并涂上凡士林,將枝條基部黑暗處理并放在含有3~4 cm清水的玻璃瓶中培養(yǎng)[14]。休眠狀態(tài)的測(cè)定參照J(rèn)ian和王海波的研究方法[15,16]。
1.2.2 抗氧化保護(hù)酶活性的測(cè)定粗酶的提取參照王學(xué)奎的方法[17]。然后分別采用氮藍(lán)四唑光化還原法測(cè)定SOD活性,愈創(chuàng)木酚法測(cè)定POD活性,雙氧水法測(cè)定CAT活性。
1.2.3 光合速率的測(cè)定選取植株一年生枝倒數(shù)第5~8片完全展開(kāi)、未受傷的功能葉片,采用CIRAS-2 型(PP-Systems)便攜式光合儀測(cè)定葉片凈光合速率,重復(fù)3次。
1.2.4 可溶性糖和淀粉含量的測(cè)定可溶性糖和淀粉含量的測(cè)定采用蒽酮顯色法[18]。
1.2.5 數(shù)據(jù)處理應(yīng)用Microsoft Excel 2003以及GraphPad進(jìn)行圖表繪制,試驗(yàn)數(shù)據(jù)均采用DPS7.05軟件進(jìn)行單因素方差法分析,LSD法進(jìn)行差異顯著性檢驗(yàn)。
2 實(shí)驗(yàn)結(jié)果
2.1 新梢生長(zhǎng)情況和休眠狀態(tài)的測(cè)定

圖 1 不同需冷量桃樹(shù)休眠進(jìn)程的界定
由圖1A可知,‘常綠桃’在休眠誘導(dǎo)期一直保持生長(zhǎng)趨勢(shì),但是從10月10日起生長(zhǎng)緩慢。在9月10日,‘春捷’的一年生枝達(dá)到81.5 cm后不再增加。在8月20日,‘青州蜜桃’一年生枝平均長(zhǎng)度達(dá)到57.6 cm后不再增加。圖1B表明,清水插枝的‘常綠桃’一年生枝條在10 d內(nèi)都可以萌發(fā)而‘春捷’第一個(gè)芽萌發(fā)時(shí)間為13 d,此時(shí)表明‘春捷’桃芽開(kāi)始進(jìn)入休眠誘導(dǎo)期,10月30日枝條萌發(fā)天數(shù)為40 d,說(shuō)明‘春捷’桃芽進(jìn)入了自然休眠期。在8月20日,青州蜜桃第一芽萌發(fā)需要的時(shí)間為14 d,表明‘青州蜜桃’桃芽進(jìn)入休眠誘導(dǎo)期,11月10日之后萌發(fā)天數(shù)超過(guò)42 d,表明芽體已進(jìn)入自然休眠期。說(shuō)明在休眠誘導(dǎo)期,‘常綠桃’相比于‘春捷’和‘青州蜜桃’無(wú)停長(zhǎng)現(xiàn)象并一直保持生長(zhǎng)的趨勢(shì)。
2.2 ‘常綠桃’和休眠桃葉片在休眠誘導(dǎo)期抗氧化酶活性的變化
由圖2A可知,在休眠誘導(dǎo)后期,SOD活性發(fā)生顯著變化,隨休眠誘導(dǎo)的加深,不同需冷量桃葉片的SOD活性呈緩慢上升趨勢(shì)。9月10日后,‘常綠桃’葉片的SOD活性迅速增加,10月30日左右出現(xiàn)峰值,為450.47 U·g-1(FW),之后下降。‘春捷’和‘青州蜜桃’葉片的SOD活性變化趨勢(shì)一致,在10月20日,‘春捷’葉片的SOD活性上升至峰值365.73 U·g-1(FW)后緩慢下降,變化趨勢(shì)不明顯。‘青州蜜桃’葉片的SOD活性在11月10日出現(xiàn)峰值,之后快速下降,其峰值為‘春捷’的1.06倍,差異不顯著。‘常綠桃’葉片SOD活性的峰值是‘春捷’的1.23倍、‘青州蜜桃’的1.15倍,差異顯著。
由圖2B可知,進(jìn)入休眠誘導(dǎo)期后,‘常綠桃’和‘春捷’葉片的POD活性呈降低的變化趨勢(shì),9月10日以后,‘常綠桃’葉片的POD活性高于‘春捷’和‘青州蜜桃’,且在9月20日出現(xiàn)峰值5.73 U·g-1(FW)·min-1,之后下降。‘春捷’在8月30日上升至峰值5.34 U·g-1(FW)·min-1以后,迅速下降。‘青州蜜桃’在10月20日左右回升出現(xiàn)峰值4.82 U·g-1(FW)·min-1,之后迅速下降。在休眠誘導(dǎo)期內(nèi),‘常綠桃’峰值分別比‘春捷’和‘青州蜜桃’高19.1%和7.4%,差異顯著,但在整個(gè)休眠誘導(dǎo)期期間‘常綠桃’葉片的POD活性差異不大,‘春捷’和‘青州蜜桃’卻顯著降低,說(shuō)明POD主要在生長(zhǎng)期向休眠期轉(zhuǎn)換的過(guò)程中發(fā)揮作用。
在休眠誘導(dǎo)期,不同蓄冷量桃葉片CAT活性呈先增加后降低的變化趨勢(shì)。在10月30日,‘常綠桃’葉片CAT活性上升至峰值95.43 U·g-1(FW)·min-1,其峰值為‘春捷’的86.5%、‘青州蜜桃’的82.5%,差異顯著。在10月20日,‘青州蜜桃’葉片CAT活性迅速下降,顯著低于‘春捷’70.32 U·g-1(FW)·min-1和‘常綠桃’80.32 U·g-1(FW)·min-1。
從抗氧化酶活性的變化可以看出,在休眠誘導(dǎo)期不同需冷量桃表現(xiàn)出顯著的差異,‘常綠桃’的抗氧化保護(hù)酶活性高于‘春捷’和‘青州蜜桃’。
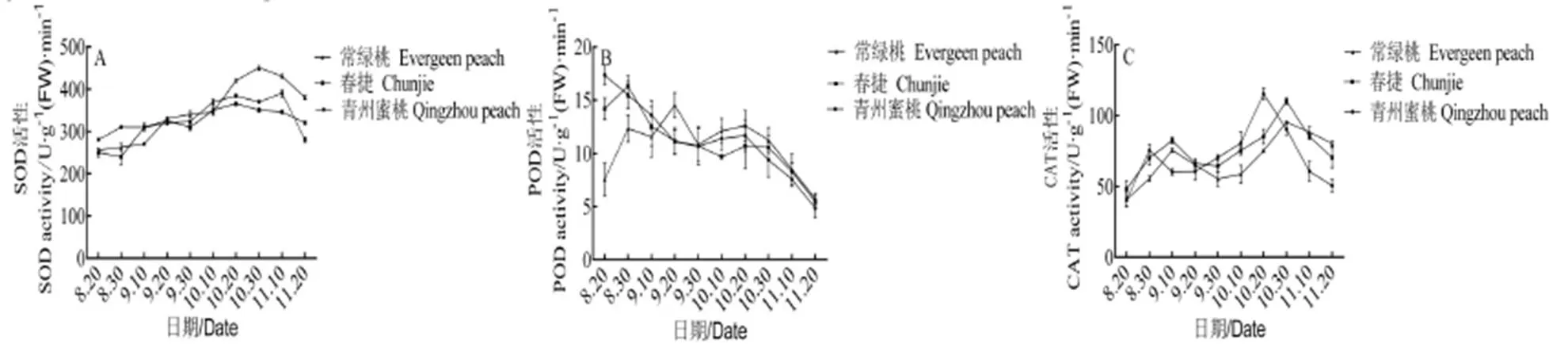
圖 2 不同需冷量桃在休眠誘導(dǎo)期保護(hù)酶活性的變化
2.3 ‘常綠桃’和休眠桃在休眠誘導(dǎo)期葉片光合特性的差異
從圖3A可以看出,進(jìn)入休眠誘導(dǎo)期后,不同需冷量桃的凈光合速率均呈下降趨勢(shì)。在9月20日,‘常綠桃’的凈光合速率上升至峰值,為18.1mmol·m-2·s-1,隨后迅速下降,10月10日后,下降速率減緩。‘春捷’在8月30日凈光合速率達(dá)到最大值,為18.2mmol·m-2·s-1,之后下降。而‘青州蜜桃’峰值最低,為16.7mmol·m-2·s-1。在11月20日,‘常綠桃’凈光合速率仍然可以維持在10.09mmol·m-2·s-1,而‘春捷’和‘青州蜜桃’在休眠誘導(dǎo)期下降趨勢(shì)明顯,并最終維持在5.15mmol·m-2·s-1和3.24mmol·m-2·s-1。
氣孔導(dǎo)度是植物葉片中氣體和水分交換的影響因子,其值的大小對(duì)植物的蒸騰速率以及O2、CO2、水蒸汽的交換速率有直接的影響。在生長(zhǎng)期向休眠期轉(zhuǎn)換的過(guò)程中,不同需冷量桃的Gs均呈先增加后降低的變化趨勢(shì),‘常綠桃’的氣孔導(dǎo)度于9 月10日左右達(dá)到最大值,為194.01 mmol·m-2·s-1。‘春捷’和‘青州蜜桃’氣孔導(dǎo)度的最大值分別出現(xiàn)在8月30日之前和8月20日之前,進(jìn)入休眠誘導(dǎo)期后顯著下降。在11月20日,‘春捷’的氣孔導(dǎo)度約為70.54 mmol·m-2·s-1,‘青州蜜桃’約為45.71 mmol·m-2·s-1,‘常綠桃’的氣孔導(dǎo)度峰值分別是‘春捷’和‘青州蜜桃’的1.51和2.31倍。
胞間二氧化碳濃度是確定光合速率變化的主要原因,其大小主要取決于葉片周?chē)諝獾腃O2濃度、氣孔導(dǎo)度、葉肉導(dǎo)度以及葉肉細(xì)胞的光合活性。圖3C顯示,8月20日以后,不同需冷量桃葉片的胞間二氧化碳濃度均呈上升趨勢(shì),在10月10日~11月10日之間,‘常綠桃’葉片胞間二氧化碳濃度低于‘春捷’和‘青州蜜桃’,其它時(shí)期高于‘春捷’和‘青州蜜桃’。在10月30日,‘春捷’葉片的胞間二氧化碳濃度達(dá)到峰值,為349.11 μmol·mol-1。‘青州蜜桃’的胞間二氧化碳濃度在整個(gè)休眠誘導(dǎo)期都低于‘常綠桃’和‘春捷’。
從光合參數(shù)的變化情況可以看出,在整個(gè)休眠誘導(dǎo)期,不同需冷量桃的光合性能存在差異,‘常綠桃’的凈光合速率高于‘春捷’和‘青州蜜桃’。
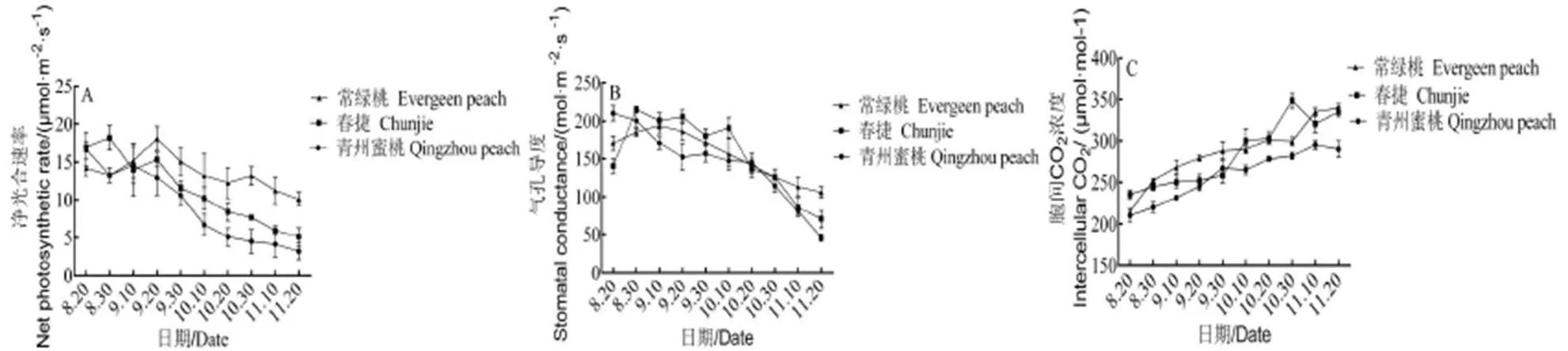
圖 3 不同需冷量桃樹(shù)在自然休眠誘導(dǎo)期光合參數(shù)的變化
2.4 ‘常綠桃’和休眠桃葉片在休眠誘導(dǎo)期可溶性糖和淀粉含量的變化
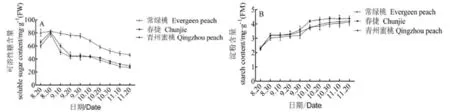
圖 4 不同需冷量的桃樹(shù)品種葉片在休眠誘導(dǎo)期可溶性糖和淀粉含量的變化
由圖4A可以看出,在生長(zhǎng)期向休眠期轉(zhuǎn)換的過(guò)程中葉片可溶性糖含量降低,‘常綠桃’葉片的可溶性糖含量在8月30日達(dá)到峰值82.07 mg·g-1后呈緩慢下降趨勢(shì),在10月10日降幅最大,為14.71%;‘春捷’和‘青州蜜桃’葉片在休眠誘導(dǎo)期可溶性糖含量差異不顯著,但是在8月30日達(dá)到峰值以后,迅速下降,‘春捷’最大降幅為25.9%,‘青州蜜桃’為35.9%,之后變化平緩。‘常綠桃’葉片可溶性糖含量在整個(gè)休眠誘導(dǎo)期都顯著高于‘春捷’和‘青州蜜桃’,最高時(shí)分別為‘春捷’和‘青州蜜桃’的1.69和1.76倍,其峰值無(wú)顯著差異。
在休眠誘導(dǎo)期,‘常綠桃’和休眠桃葉片淀粉含量增加。由圖4B可以看出,在8月20日,‘常綠桃’葉片的淀粉含量突然增加,上升幅度為29.31%,之后緩慢上升,但是變化不明顯,持續(xù)一個(gè)月以后,含量又突然增加,由3.2 mg·g-1升至3.8 mg·g-1,上升幅度為18.75%,此后葉片淀粉含量又保持相對(duì)穩(wěn)定但是低于‘春捷’和‘青州蜜桃’。在整個(gè)休眠誘導(dǎo)期‘春捷’葉片淀粉含量高于‘常綠桃’和‘青州蜜桃’。在8月20日,‘青州蜜桃’葉片淀粉含量突然增加以后,呈緩慢上升的變化趨勢(shì)。
從不同需冷量桃葉片的碳水化合物含量變化可以看出,‘常綠桃’葉片的可溶性糖含量顯著高于‘春捷’和‘青州蜜桃’但是淀粉含量卻低于‘春捷’和‘青州蜜桃’,說(shuō)明‘常綠桃’葉片碳水化合物具有較強(qiáng)的流動(dòng)性。
3 討論
3.1 ‘常綠桃’和休眠桃葉片在休眠誘導(dǎo)期抗氧化保護(hù)酶活性分析
落葉果樹(shù)的休眠是植物在生長(zhǎng)進(jìn)化過(guò)程中一個(gè)重要的生物學(xué)特性,通過(guò)休眠使植物生長(zhǎng)停止而度過(guò)不良的環(huán)境[19],植物在休眠的過(guò)程中主要通過(guò)葉片來(lái)感知一系列的生長(zhǎng)變化,而活性氧可攻擊葉片中生命大分子物質(zhì)和細(xì)胞壁,造成機(jī)體的損傷和病變,加速機(jī)體的衰老進(jìn)程[20]。SOD可以清除新陳代謝過(guò)程中產(chǎn)生的超氧陰離子自由基O2-,POD和SOD主要將H2O2水解形成水和氧氣避免產(chǎn)生對(duì)植物有害的OH-,并且可以有效地抑制膜脂過(guò)氧化[21]。本研究中,‘常綠桃’和休眠桃在休眠誘導(dǎo)前期無(wú)明顯差異,但是隨著休眠誘導(dǎo)的加深,‘常綠桃’葉片的酶活性高于‘春捷’和‘青州蜜桃’。其主要原因可能是落葉桃樹(shù)受到外界不良環(huán)境后表現(xiàn)出休眠特性,而‘常綠桃’為了適應(yīng)外界不良環(huán)境,通過(guò)抗氧化酶活性的增加來(lái)清除體內(nèi)產(chǎn)生的活性氧等有害物質(zhì),使‘常綠桃’保持明顯的生長(zhǎng)勢(shì)。
3.2 ‘常綠桃’和休眠桃葉葉片在休眠誘導(dǎo)期糖含量變化差異分析
糖不僅可以提供能源而且是一種信號(hào)物質(zhì),在信號(hào)轉(zhuǎn)導(dǎo)過(guò)程中起到信號(hào)調(diào)節(jié)的作用。碳水化合物作為植物體內(nèi)的貯藏物質(zhì)具有一定的流動(dòng)性[22],并且在低溫條件下,是一種非常重要的滲透調(diào)節(jié)物質(zhì),可以提高滲透作用,增強(qiáng)植株抵御外界不良環(huán)境的能力[23,24]。在休眠誘導(dǎo)期,溫度開(kāi)始降低,光合作用也相應(yīng)減弱,植株出現(xiàn)限碳的情況,為了維持正常的生命活動(dòng)和防止冷誘導(dǎo)損傷,植株會(huì)調(diào)節(jié)淀粉與可溶性糖之間的碳水化合物代謝平衡[25,26]。在這一時(shí)期,不同需冷量桃葉片的可溶性糖含量降低,其中‘常綠桃’葉片可溶性糖含量高于‘春捷’和‘青州蜜桃’,但是淀粉含量無(wú)顯著差異,這可能與‘常綠桃’在休眠誘導(dǎo)期仍保持較高的光合速率有關(guān)。進(jìn)入休眠誘導(dǎo)期以后,‘春捷’和‘青州蜜桃’葉片可溶性糖含量突然降低,而‘常綠桃’葉片可溶性糖含量降幅較緩,這可能與不同需冷量桃適應(yīng)不良環(huán)境能力的不同有關(guān),當(dāng)進(jìn)入休眠誘導(dǎo)期以后,休眠桃的碳水化合物從葉片中快速轉(zhuǎn)移,通過(guò)提高滲透勢(shì)來(lái)抵御外界不良環(huán)境,而‘常綠桃’則一直保持生長(zhǎng)優(yōu)勢(shì),碳水化合物的轉(zhuǎn)移沒(méi)有明顯的變化。
4 結(jié)論
不同需冷量桃在休眠誘導(dǎo)期表現(xiàn)出不同的生理變化,‘常綠桃’能夠保持較高的抗氧化保護(hù)酶活性和光合效率,且可溶性糖含量高于‘春捷’和‘青州蜜桃’,淀粉含量低于‘春捷’和‘青州蜜桃’,在春捷’和‘青州蜜桃’停長(zhǎng)時(shí)仍能保持良好的生長(zhǎng)勢(shì)。
[1] Lang GA, Early JD,Martin GC,Endo-, para-, and ecodormancy: physiological terminology and classification for dormancy research[J]. Hortscience, 1987,22(3):371-377
[2] Sherson SM, Alford HL, Forbes SM,. Roles of cell wall invertases and monosaccharide transporters in the growth and development of Arabidopsis[J]. Journal of Experimental Botany, 2003,54(382):525-531
[3] 王海波,王孝娣,高東升,等.不同需冷量桃品種芽休眠誘導(dǎo)期間的生理變化[J].果樹(shù)學(xué)報(bào),2009,26(4):445-449
[4] 代漢萍,薛志杰,郭修武.樹(shù)莓自然越冬枝條生理生化特性研究[J].吉林農(nóng)業(yè)大學(xué)學(xué)報(bào),2009,31(5):573-577
[5] 李冬梅,譚秋平,高東升,等.光周期對(duì)休眠誘導(dǎo)期桃樹(shù)光合及PSⅡ光系統(tǒng)性能的影響[J].應(yīng)用生態(tài)學(xué)報(bào),2014,25(7):1933-1939
[6] 李冬梅.光周期誘導(dǎo)設(shè)施桃樹(shù)休眠的生理生化變化及蛋白質(zhì)組學(xué)初探[D].泰安:山東農(nóng)業(yè)大學(xué),2012:12-15
[7] González-rossia D, Reig C, Dovis V,. Changes on carbohydrates and nitrogen content in the bark tissues induced by artificial and its relationship with dormancy bud break insp.[J]. Sciential Horticulturae, 2008,118(4):275-281
[8] Wang K, Shao XF, Gong YF,. The metabolism of soluble carbohydrates related to chilling injury in peach fruit exposed to cold stress[J]. Postharvest Biology & Technology, 2013,86(3):53-61
[9] Koehler G, Wilson RC, Goodpaster JV,. Proteomic study of low-temperature responses in strawberry cultivars (Fragaria×ananassa) that differ in cold tolerance[J]. Plant Physiology, 2012,159(4):1787-1805
[10] Ghrab M, Mimoun MB, Masmoudi MM,. Chilling trends in a warm production area and their impact on flowering and fruiting of peach trees[J]. Scientia Horticulturae, 2014,178(8):87-94
[11] Horvath DP, Chao WS, Anderson JV. Molecular analysis of signals controlling dormancy and growth in underground adventitious buds of leafy spurge[J]. Plant Physiology, 2002,128(4):1439-1446
[12] 孫凌俊,呂春晶,馬麗,等.‘巨峰’葡萄休眠及解除過(guò)程糖類(lèi)物質(zhì)變化研究[J].中國(guó)農(nóng)學(xué)通報(bào),2017,33(28):93-98
[13] Mohamed HB, Vadel AM, Geuns JMC,. Biochemical changes in dormant grapevine shoot tissues in response to chilling: possible role in dormancy release[J]. Scientia Horticulturae, 2010,124(4):440-447
[14] 李玲.桃花芽休眠解除SSH文庫(kù)構(gòu)建及相關(guān)基因的功能分析[D].泰安:山東農(nóng)業(yè)大學(xué),2011:34-35
[15] Jian LC, Li PH, Sun LH,. Alteration in ultrastructure and subcellular localization of Ca2+in poplar apical bud cells during the induction of dormancy[J]. Journal of Experimental Botany, 1997,48(6):1195-1207
[16] 王海波,王孝娣,程存剛,等.桃芽休眠的自然誘導(dǎo)因子及鈣在休眠誘導(dǎo)中的作用[J].應(yīng)用生態(tài)學(xué)報(bào),2008,19(11):2333-2338
[17] 王學(xué)奎.植物生理生化實(shí)驗(yàn)原理和技術(shù)[M].北京:高等教育出版社,2006:211-232
[18] 李合生.植物生理生化試驗(yàn)原理和技術(shù)[M].北京:高等教育出版社,2000:250-251
[19] Kalberer SR, Wisniewski M, Arora R. Deacclimation and reacclimation of cold-hardy plants: Current understanding and emerging concepts[J]. Plant Sciece, 2006,171(1):3-16
[20] Martinez-reyes I, Cuezva JM. The H(+)-ATP synthase: Agate to ROS-mediated cell death or cell survival[J]. Biochimica Biophysica Acta, 2014,1837(7):1099-1112
[21] 李春燕,陳思思,徐雯,等.苗期低溫脅迫對(duì)揚(yáng)麥葉片抗氧化酶和滲透調(diào)節(jié)物質(zhì)的影響[J].作物學(xué)報(bào),2011,37(12):2293-2298
[22] Ley S, Dolger K, Appenroth KJ. Carbohydrate metabolism as a possible physiological modulator of dormancy in turions of(L.) Schleiden[J]. Plant Science, 1997,129(1):5-7
[23] Tabaeiaghdaei SR, Pearce RS, Harrison P. Sugars regulate cold-induced gene expression and freezing-tolerance in barley cell cultures[J]. Journal of Experimental Botany, 2003,54(387):1565-1575
[24] 施征,白登忠,張維誠(chéng),等.青海云杉休眠前后非結(jié)構(gòu)性碳水化合物含量隨海拔變化[J].林業(yè)科學(xué)研究,2017,30(6):908-915
[25] Ito A, Sugiura T, Sakamoto D,. Effects of dormancy progression and low-temperature response on changes in the sorbitol concentration in xylem sap of Japanese pear during winter season[J]. Tree Physiology 2013,33(4):398-408
[26] Anderson JV, Gesch RW, Jia Y,. Seasonal shifts in dormancy status, carbohydrate metabolism, and related gene expression in crown buds of leafy spurge[J]. Plant, Cell & Environment, 2005,28(12):1567-1578
Physiological Changes of Peach Cultivars with Three Different Chilling Requirement during Dormancy Induction
WEN Bin-bin1, ZhANG Xin-hao1, CHEN Xiu-de1,2, GAO Dong-sheng1,2, ZHU Cui-ying1*, XIAO Wei1,2*
1.271018,2.271018,
Physiological changes of peach leaves in dormancy induction period (20thAugust-20thNovember) were studied with three different chilling requirement peach cultivars ‘evergreen peach’ (no), ‘Chunjie’ (102~120 C.U.), ‘Qingzhou’ Peach (1200 C.U.) was grafted on the same peach rootstock. The result indicated that the activities of antioxidant protective enzymes, photosynthetic rate, soluble sugar and starch content in peach leaves all significantly changed during the natural dormancy induction. Along with dormancy developing, the activity of superoxide dismutase (SOD) increased and in the late dormancy induction period (10thOctober), the evergreen peach was higher than that of ‘Chun Jie’ and ‘Qingzhou peach’, the activity of peroxidase (POD) decreased and there was no significant difference between peach cultivars with different chilling requirement, the activity of catalase (CAT) changed as double Peak and the peak value of ‘evergreen peach’ were lower than dormant peach; the net photosynthetic rate decreased but the value of everygreen peach was higher than ‘Chun Jie’ and ‘Qingzhou peach’ and the peak was 1.96 and 3.13 times of ‘chunjie’ and ‘qingzhou’ peach respectively. The content of soluble sugar decreased and the starch content increased but the content of soluble sugar of evergreen peach was higher than ‘Chun Jie’ and ‘Qingzhou peach’, starch content was lower than them. The above results showed that the protective enzymes activity, photosynthesis rate and soluble sugar content was higher than ‘Chun Jie’ and ‘Qingzhou peach’, the starch content was lower than ‘Chun Jie’ and ‘Qingzhou peach’.
Peach; dormancy induction; Physiological changes; chilling requirement; peach leaf
S662.1
A
1000-2324(2019)01-0001-04
10.3969/j.issn.1000-2324.2019.01.001
2018-07-15
2018-08-12
山東省現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系果品創(chuàng)新團(tuán)隊(duì)項(xiàng)目(SDAIT-06-01)
文濱濱(1993-),男,碩士,研究方向:設(shè)施果樹(shù)休眠生理與分子. E-mail:wbbsdau@163.com
Author for correspondence. E-mail:chunying196217@163.com; gulight986918@163.com
