簡析表觀遺傳學
2019-03-08 14:33:09陶艾艾
中學生物學 2019年11期
關鍵詞:機制
陶艾艾
表觀遺傳學是一門由現象到領域的學科,是近年來現代遺傳學發展最快的一個分支領域。隨著各國科研人員的不斷深入研究,對表觀遺傳學及其機制的認識和理解有了突破性進展,這意味著表觀遺傳學的內容必然要納入高中生物新課程中。對于大多數中學生物教師和中學生而言,這是一個全新的概念和全新的領域。現就表觀遺傳學的概念、調控機制、特征以及研究進展進行綜述。
1表觀遺傳學的誕生
“表觀遺傳學”這個詞于1942年由英格蘭生物學家沃丁頓在Endeavour雜志中第一次正式提出,且沃丁頓指出“表觀遺傳學”與“遺傳學”是一對相對應的概念,但是當時沒有引起大多數人的重視。“表觀遺傳學”經過40多年的沉寂,又被重新提出,并開始迅速發展。經典遺傳學是指基因型的遺傳,生物體細胞中遺傳物質發生改變即遺傳信息改變而導致可遺傳現象。現代生物學的中心法則即“DNA<=>RNA=>蛋白質”涵蓋了生命所必需的遺傳信息模板的維持和表達過程,卻否認了蛋白質到DNA的反饋過程;但是,有些稀有的蛋白質,例如朊粒通過改變其同源基因產物,從而導致可遺傳的表型改變,即可以在沒有DNA或者RNA模板的情況下被繼承,這看起來似乎有悖于中心法則。但其實是這些特殊的自我聚集的蛋白質具有一些和DNA類似的性質,包括復制和信息的儲存機制。生命是一個復雜的過程,經典遺傳學已經不能解釋所有的生命現象,表觀遺傳學就是在研究與經典孟德爾遺傳定律不相符合的許多遺傳現象(并非指細胞質遺傳)過程中逐步發展起來的。
表觀遺傳是指DNA序列不改變的前提下,在基因組染色質水平上調控基因表達的細胞親子代間的遺傳行為,由此改變了人們對基因組的單一認識,即基因組序列不僅包含有傳統意義上的遺傳信息,而且還包含由相關修飾而產生的表觀遺傳信息。表觀遺傳學則是對表觀遺傳及其機制進行研究的一門學科,是闡明基因組功能及基因表達調控的關鍵研究領域,也是一門從現象到領域的遺傳學分支學科。基因組中的遺傳信息可以為蛋白質的合成提供模板,而表觀遺傳信息卻會影響基因的功能和活性,如基因能否表達、什么時候表達、表達強度如何,以及如何表達。
2表觀遺傳學的主要調控機制
2.1DNA甲基化
DNA甲基化是已知最早被發現的表觀遺傳學機制,也是基因表達調控的主要表觀遺傳修飾方式,可以調節基因的功能,在酵母以外的所有真核生物中都有不同程度的存在。DNA甲基化是指在DNA序列不改變的前提下,在DNA甲基化轉移酶的作用下,主要將特定位點的胞嘧啶脫氧核苷酸5′-端的胞嘧啶進行甲基化,使之轉化為5′-甲基胞嘧啶。而且這種DNA的甲基化是可逆的,在DNA去甲基化酶的作用下,甲基化的DNA可以完成去甲基化過程。在脊椎動物中,DNA的甲基化或者去甲基化主要發生于啟動子區域的胞嘧啶與鳥嘌呤含量豐富的區域,即所謂的CpG島[CpG即胞嘧啶(C)-磷酸(p)-鳥嘌呤(G)的縮寫],而植物中除了CpG島甲基化外,還有CHH和CHG的甲基化(H可以為A或C或T)。
DNA的甲基化狀態有三種:
1DNA的高度甲基化狀態,如哺乳動物雌性個體體細胞內的兩條X染色體會失活其中一條;
2DNA的低甲基化狀態,如始終保持低甲基化狀態的管家基因,可以一直處于活性轉錄狀態;
3DNA去甲基化狀態,如細胞分化的某種狀態下或者生物個體發育中的某一階段中,原來甲基化的DNA,也可以被誘導去除甲基化,從而使原本沉默的基因表現出轉錄活性。
總而言之,DNA甲基化一般會抑制基因表達或者說DNA甲基化可以使基因沉默,而DNA去甲基化后則可以使基因活化從而能正常表達。
2.2組蛋白修飾
組蛋白是組成真核生物染色質的基本結構,因富含精氨酸和賴氨酸等堿性氨基酸而呈堿性,可以與酸性的DNA緊密結合。組蛋白修飾是指在相關酶系統的催化下,組蛋白的某些氨基酸(主要是精氨酸或賴氨酸)上會發生乙酰化、甲基化、磷酸化、腺苷酸化、泛素化、類泛素化、ADP核糖基化等的修飾過程,也可在其對應的逆向修飾酶系統的作用下可發生相反的過程,如組蛋白去乙酰化酶可以去除這些乙酰化基團標簽。組蛋白的修飾會影響組蛋白和DNA的結合,從而改變染色質的結構狀態,使之疏松或者螺旋化;同時會影響某些蛋白質因子與染色質的結合,還可影響轉錄因子與特定DNA序列的結合,從而間接地調控基因的表達過程,引起細胞或個體的表型改變。組蛋白修飾并不是獨立發生的,而是在特定的組蛋白尾部的鄰近殘基發生多個共價修飾,即組合成所謂的修飾盒,或者是不同組蛋白尾巴或者核小體間的共價修飾組合即反式組蛋白通路,它們協同或者拮抗起作用。
2.3染色質重塑染色質重塑即染色質結構的改變,是指在需要??? 耗能的情況下核小體的發生解離和重配或者組蛋白共價修飾,使得染色質的結構疏松或者凝集。染色質結構的改變主要包括兩種類型:
1依賴共價結合反應的化學修飾,即在組蛋白的末端進行共價修飾,如乙酰化、甲基化、磷酸化等。
2依賴ATP的物理修飾,在ATP的供能下,能使核小體發生滑動或者重新置配。
當染色質結構發生改變,也就是染色體重塑,就會影響附近基因的功能和活性。
2.4非編碼RNA調控
非編碼RNA是細胞內不編碼蛋白質的RNA的統稱,包括兩種類型:
1管家型非編碼RNA,包含rRNA、tRNA,此外還有核內小RNA(snRNA)、核仁小RNA(snoRNA)。
2調節型(或者說功能型)非編碼RNA,包括短鏈非編碼RNA[sncRNA,如小干擾RNA(siRNA)、微小RNA(miRNA)及與Piwi蛋白相互作用的RNA(piRNA)等]和長鏈非編碼RNA(lncRNA)。sncRNA主要參與mRNA的降解和誘導染色質結構的改變,從而調控基因表達過程,決定著細胞的分化命運,還起著保護自身的基因組的作用,參與降解外源DNA。lncRNA可以通過與DNA、RNA和蛋白質這些生物大分子發生相互作用來調控基因表達過程。具體來說,lncRNA可通過調節組蛋白修飾、染色質重塑、一些蛋白質的功能和RNA代謝等,從基因轉錄水平和轉錄后水平對基因表達調控。有實驗證據表明lncRNA表達失調將導致廣泛的細胞信號通路紊亂。
2.5其他機制
除了上面提及的4種機制外,還有基因印記、X染色體失活等。而基因印記和X染色體失活的本質仍然是DNA甲基化、組蛋白修飾和染色質重塑。這些機制往往都是相互關聯的,而且具有一定的層次性,它們之間的具體關系以及它們如何共同調控表觀遺傳,還需要進一步研究。
3表觀遺傳學的特征
3.1DNA序列不變
表觀遺傳學不涉及細胞核中DNA序列即遺傳物質的變化,只是對DNA和蛋白質進行修飾或是利用非編碼RNA或是影響染色質的結構來調控基因的功能。例如,同卵雙胞胎擁有完全相同的DNA序列,但是外觀卻存在微小的差異,而且對疾病易感成度也存在著一定的差異。這些顯然不能通過DNA序列相同來解釋,而是表觀遺傳調控的結果。
3.2可繼承性即可遺傳
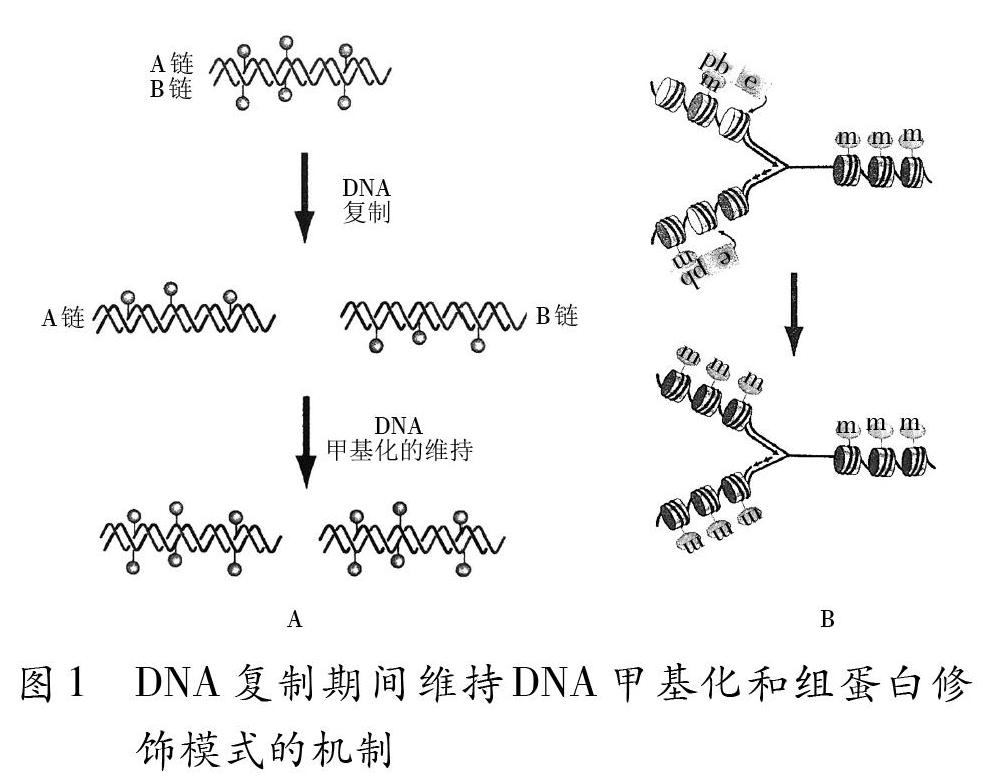
基因組的遺傳信息可以通過有絲分裂和減數分裂遺傳給子代細胞或子代個體。那么,這些表觀遺傳學修飾是如何通過細胞分裂遺傳給子代細胞的呢?目前具體機制仍然不是很清楚,通常認為DNA甲基化的修飾作用在配子形成過程中被清除并重新設定,由此阻斷了這種修飾通過減數分裂在世代間傳遞。但是,至少哺乳動物某些基因的DNA甲基化標記在減數分裂過程中沒有完全被清除從而能使之傳遞給子代。DNA甲基化、組蛋白修飾有類似DNA半保留復制的延伸機制如圖1所示。
圖1A為DNA復制期間維持DNA甲基化模式的機制。DNA在半保留復制期間,一條DNA鏈的CpG或者CpXpG殘基上存在特異的甲基化模式,開始配對合成另一條新的未甲基化的DNA鏈。每條DNA鏈上的CpG會有相應的CpG在另一條鏈上存在。此時DNA甲基轉移酶會識別半甲基化的位點并甲基化新鏈上的胞嘧啶,以維持甲基化的模式不被破壞。圖1B所示為復制期間維持組蛋白修飾的基本機制。修飾的組蛋白尾部(m)與含有特異的修飾結合位點的蛋白結合子(pb)結合;反之,pb也含有與特異的進行組蛋白修飾的酶(e)的結合位點。酶(e)進而能修飾相鄰的核小體。復制期間新插入的組蛋白與親本組蛋白是散在分布的,因此獲得了親本的修飾。在細胞周期的任何階段相似的機制使得組蛋白修飾從一個修飾的區城向未修飾的區域延伸。另外,還有一些是通過非依賴復制的方式來繼續保持其在子代中原有的狀態。大部分表觀遺傳信息是如何被繼承并傳播的,細胞記憶的分子機制是什么?......這些機制還需要國內外參與表觀遺傳學研究的科研人員不斷地去探索去破譯。
3.3具有可逆性
DNA的甲基化修飾可以通過去甲基化酶的作用去除,同樣,其他的修飾也具有類似的可逆機制。因此表觀遺傳的修飾是可逆的,這一點對人類相關疾病的防治研究具有十分重要意義。例如,異常的DNA甲基化修飾常會伴有復雜的突變和表型缺陷的多種人類疾病,這為人類疾病的防治提供了新的思路。
3.4受環境因素影響
一些環境因素可以通過改變生物的遺傳物質來改變遺傳信息從而影響細胞和個體的表現型。同樣,很多環境因素也可以引起表觀遺傳信息的改變,
如飲食習慣、壓力因素、吸煙等會改變表觀基因組(尤其是DNA甲基化)。例如,以基因型為aa的母鼠及其孕育的基因型為AVYa的仔鼠作實驗對象。孕鼠分為兩組,實驗組孕鼠相比對照組孕鼠除了飼喂標準飼料外,另外還從受孕前兩周起在飼料中添加富含甲基的葉酸和乙酰膽堿等。結果對照組孕鼠產下的仔鼠的毛色大多數是黃色,而實驗組孕鼠產下的仔鼠大多數出現了大小不一的棕色斑塊,甚至出現了以棕褐色毛色為主的仔鼠。分析表明喂以富甲基飼料的孕鼠所產仔鼠的IAP(IAP是鼠毛色控制基因的第一外顯子前插入的一段逆轉座子,鼠毛色控制基因間接地受到了IAP啟動子的影響)所含CpG島的甲基化平均水平遠高于對照組,這就意味著轉錄調控區DNA的高甲基化狀態使得原來應該表達的基因功能喪失,趨于沉默,毛色也出現了棕褐色。
4表觀遺傳學的研究進展
隨著對表觀遺傳學及其機制的不斷深入研究,越來越多的人們關注到表觀遺傳學。1999年,英德法三國成立了人類表觀基因組協會(HEC)。2003年,人類基因組測序工作在全球科學家多年的共同努力下,終于完成了。同年10月,HEC正式宣布開始投資和實施人類表觀基因組計劃(HEP)。2004年,“表觀遺傳學”第一次作為主題呈現在第69屆冷泉港定量生物學會議上,表明表觀遺傳學已然成為具有鮮明特色的生物學研究領域。我國科技部也于2005年啟動了“腫瘤和神經系統疾病的表觀遺傳機制”的“973”重大專項研究。2006年,中日韓以及新加坡的科研人員召開了第一屆亞洲表觀遺傳組學聯盟年會,成為亞洲表觀遺傳學研究發展的重要交流和合作平臺。2010年1月,由多個國家參與的國際人類表觀遺傳學合作組織(IHEC)在巴黎成立,并計劃在第一階段10年內繪制1000個人類表觀基因組圖譜。德國于2012年9月正式啟動了《德國人類表觀遺傳學研究計劃》(DEP),其發展目標是標記測量健康細胞和疾病細胞的表觀遺傳基因開關。近幾年,表觀遺傳學的研究已經成為人類基因組測序后的又一重大研究方向之一,是生命科學最前沿的研究領域之一。表觀遺傳學與人類健康密切相關,人類表觀基因組計劃的開展以及對表觀遺傳學機制的不斷深入研究,將對人類疾病的防治、研發新藥物以及對基因何時何地以何方式表達的研究有著深遠影響。
參考文獻:
[1]C.D.Allis,T.Jenuwein,D.Reinberg,等編著.朱冰,孫方霖主譯.表觀遺傳學[M].北京:科學出版社,2009:29,46,21-22
[2]程曉剛.表觀遺傳學教學實踐與體會[J].基礎醫學教育,2011,13(4):306-307.
[3]李文藝.表觀遺傳學及其研究進展[J].安徽農業科學,2009,37(14):6358-6360.
[4]薛京倫.表觀遺傳學——原理、技術與實踐[M].上海:上海科學技術出版社,2006:4-5.
[5] Long HK,Blackledge NP,Klose RJ. ZF -CxxC domain-con? taining proteins,CpG islands and the chromatin connection[J]. BiochemSocTrans,2013,41(3):727-740.
[6]王瑞嫻,徐建紅.基因組DNA甲基化及組蛋白甲基化[J].遺傳,2014,36(3):191-199.
[7]高原,周劍濤,夏德宜.表觀遺傳學與人類表觀基因組計劃[J].Progress in Modern Biomedicine,2009,9(2):365-367.
[8]王麗娟,劉家熙.表觀遺傳及表觀遺傳學概述[J].生物學教學,2017,42(1):2-4.
[9]李靈,宋旭.長鏈非編碼RNA在生物體中的調控作用[J].遺傳,2014,36(3):228-236.
[10]孫亮,唐愛發,桂耀庭,等.DNA甲基化的世代傳遞[J].生命的化學,2010,30(5):685-687.
[11]王麗娟,劉家熙.表觀遺傳及表觀遺傳學概述[J].生物學教學,2017,42(1):2-4.
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
