小RNA病毒3C蛋白功能的研究進(jìn)展
2019-02-28 07:02:17曹圣慧黃孝天
中國人獸共患病學(xué)報(bào) 2019年12期
關(guān)鍵詞:結(jié)構(gòu)研究
曹圣慧,黃孝天
1 小RNA病毒科的基因組結(jié)構(gòu)與致病性
1.1小RNA病毒基因組結(jié)構(gòu) 小RNA病毒科基因組為單正鏈RNA,大小為6.7~10.1 kb,病毒顆粒呈球形,無被膜包被,病毒衣殼為二十面體結(jié)構(gòu)[1]。小RNA病毒基因組結(jié)構(gòu)高度保守,包括5′非編碼區(qū)(5′-noncoding region,5′-NCR)、開放閱讀框、3′非編碼區(qū)(3′-NCR)和3′端的polyA尾。5′-NCR包含多個(gè)RNA二級(jí)結(jié)構(gòu)元件,其中一個(gè)重要的二級(jí)結(jié)構(gòu)元件是內(nèi)部核糖體進(jìn)入位點(diǎn)(Internal ribosome entry site,IRES),IRES三葉草結(jié)構(gòu)與細(xì)胞蛋白質(zhì)相互作用后招募核糖體,啟動(dòng)病毒基因組的翻譯。病毒基因組開放閱讀框包括3個(gè)部分,分別為P1區(qū)、P2區(qū)和P3區(qū)。P1區(qū)經(jīng)翻譯加工后形成結(jié)構(gòu)蛋白VP1、VP2、VP3和VP4,組成病毒的衣殼結(jié)構(gòu)。P2區(qū)和P3區(qū)分別翻譯加工形成非結(jié)構(gòu)蛋白2A、2B、2C以及3A、3B、3C、3D。其中,2A蛋白和3C蛋白分別參與病毒蛋白的剪切。3′-NCR和病毒的復(fù)制效率相關(guān),3′端的polyA尾參與病毒的復(fù)制和翻譯[2]。
1.2常見小RNA病毒的致病性 小RNA病毒科包括35個(gè)屬,80個(gè)種[1]。許多小RNA病毒可引起人和動(dòng)物的腦部、心臟、肝臟、皮膚、胃腸道或上呼吸道疾病,且目前無有效的疫苗用于治療。常見的小RNA病毒包括腸道病毒71型(Enterovirus 71,EV-71)、腸道病毒D68型(Enterovirus D68,EV-D68)、柯薩奇病毒(Coxsackievirus,CV)、脊髓灰質(zhì)炎病毒(Poliovirus,PV)、甲型肝炎病毒(Hepatitis A virus,HAV)、鼻病毒(Human rhinovirus,HRV)、口蹄疫病毒(Foot-and-mouth disease virus,F(xiàn)MDV)、腦心肌炎病毒(Encephalomyocarditis virus,EMCV)等。EV-71、CV-A2、CV-A16、CV-A6和CV-A10可導(dǎo)致每年亞太國家數(shù)百萬兒童患手足口病,甚至引起更嚴(yán)重的臨床癥狀,如無菌性腦膜炎、急性遲緩性麻痹和神經(jīng)呼吸系統(tǒng)綜合征[3]。CV-B4可引起新生兒心肌炎、Ⅰ型糖尿病以及神經(jīng)性疾病[4]。EV-D68可引起世界范圍內(nèi)人類呼吸道疾病的流行[5]。EMCV可引起動(dòng)物腦炎、心肌炎為主要特征的急性傳染病,F(xiàn)MDV可導(dǎo)致牛羊等動(dòng)物口蹄疫的暴發(fā)流行,對(duì)農(nóng)業(yè)生產(chǎn)造成重大的經(jīng)濟(jì)損失[6-7]。
2 小RNA病毒3C蛋白的結(jié)構(gòu)
小RNA病毒的3C蛋白包含175~253個(gè)氨基酸,大小為19~28 kD。3C蛋白集絲氨酸蛋白酶和半胱氨酸蛋白酶特性為一體。作為絲氨酸蛋白酶,3C蛋白具有催化三聯(lián)體Cys-His-Glu/Asp[8]。作為半胱氨酸蛋白酶,3C蛋白具有特征催化基序Gly-X-Cys-Gly[9]。近年來,多種小RNA病毒如HRV、PV、HAV等的3C蛋白晶體結(jié)構(gòu)顯示,3C蛋白都具有二個(gè)相同的、6條反向平行鏈構(gòu)成的、呈約90°的β-桶狀(β-barrel)結(jié)構(gòu)域,二個(gè)結(jié)構(gòu)域之間延伸出一個(gè)可結(jié)合底物的淺槽。其中,由十幾個(gè)氨基酸殘基構(gòu)成的環(huán)狀結(jié)構(gòu)位于淺槽的上方,稱為β-折疊(β-ribbon),它對(duì)底物的特異性識(shí)別具有重要作用[9](圖1)。研究顯示HRV、PV、HAV、FMDV中3C蛋白的β-折疊為閉合構(gòu)象,而EV-71中3C蛋白的β-折疊是一種開放構(gòu)象,位于β-折疊基底的Gly-123和 His-133控制著β-折疊的靈活性[10]。
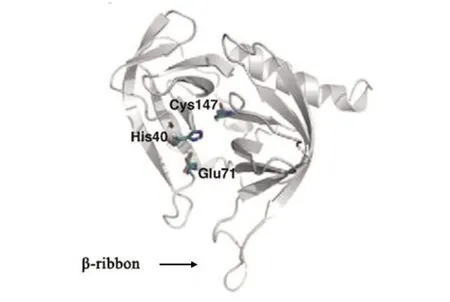
圖1 EV-71的3C蛋白結(jié)構(gòu)[10]Fig.1 Structure of EV-71 3Cpro[10]
3 小RNA病毒3C蛋白的功能
近年來對(duì)于小RNA病毒3C蛋白的研究表明,該蛋白參與病毒前體蛋白的剪切,與促進(jìn)病毒復(fù)制、調(diào)控細(xì)胞凋亡以及逃避免疫應(yīng)答等密切相關(guān)。
3.1促進(jìn)病毒復(fù)制 由于自身基因組的限制,許多病毒依賴于宿主細(xì)胞來完成自身的復(fù)制。研究發(fā)現(xiàn)3C蛋白可裂解多種蛋白如poly-A結(jié)合蛋白(poly(A)-binding protein,PABP)、剪接因子脯氨酸和谷氨酰胺(Splicing factor proline and glutamine rich,SFPQ)、蛋白激酶R (protein kinase R,PKR)等。這些蛋白對(duì)病毒復(fù)制的影響是不同的。Sun等研究表明鴨甲肝病毒(duck hepatitis A virus,DHAV)的3C蛋白可裂解PABP蛋白中 Q367和G368之間的特異性位點(diǎn),PABP蛋白被裂解為N端片段和C端片段,N端片段的存在利于DHAV的復(fù)制,而C端片段則截然相反。C端片段的功能被細(xì)胞所抑制的機(jī)制尚不清楚[11]。Kobayashi等研究結(jié)果顯示EMCV的3C蛋白可裂解PABP蛋白中的Q437和G438之間的特異性位點(diǎn),導(dǎo)致一個(gè)N端45 kD的蛋白在病毒感染的細(xì)胞中累積,促進(jìn)病毒的復(fù)制。而當(dāng)PABP的裂解受到干擾時(shí),病毒的復(fù)制明顯受到抑制。因此,EMCV的3C蛋白可通過裂解PABP而促進(jìn)病毒復(fù)制[12]。此外,Dylan等研究顯示HRV的3C/3CD 可裂解SFPQ,而SFPQ水解片段的產(chǎn)生導(dǎo)致病毒復(fù)制以及病毒顆粒的增多[13]。Chang等人發(fā)現(xiàn)EV-71感染宿主時(shí),3C通過裂解PKR增強(qiáng)了病毒的復(fù)制[14]。Yao等發(fā)現(xiàn)槲皮素能夠結(jié)合EV-71 3C蛋白的底物識(shí)別位點(diǎn)從而抑制其活性,以及抑制宿主體內(nèi)的病毒復(fù)制,推斷槲皮素抑制病毒復(fù)制的機(jī)制可能與3C蛋白的活性有關(guān)[15]。
3.2調(diào)控細(xì)胞凋亡 病毒感染宿主后可導(dǎo)致宿主細(xì)胞發(fā)生凋亡。研究表明小RNA病毒的3C蛋白可通過激活半胱天冬酶(caspase)活性,裂解PinX1蛋白、真核翻譯起始因子(eukaryotic translation initiation factor 4GI,eIF4GI)和受體相互作用蛋白激酶Ⅰ(Receptor-interacting protein kinase-1,RIPK1)調(diào)控細(xì)胞凋亡[16]。Li等研究結(jié)果顯示,當(dāng)神經(jīng)細(xì)胞表達(dá)EV-71的3C蛋白時(shí),細(xì)胞以DNA片段化和聚腺苷二磷酸核糖聚合酶(poly(ADP-ribose) polymerase,PARP)裂解的形式發(fā)生凋亡。其中,PARP裂解是caspase被激活的特異性標(biāo)志,說明3C蛋白可激活caspase誘導(dǎo)細(xì)胞凋亡[17]。Song等發(fā)現(xiàn)EV-71感染宿主后,3C蛋白可與caspase -8,9相互作用而激活caspase-3誘發(fā)凋亡,當(dāng)3C失去水解活性后,細(xì)胞caspase -8,9活性明顯降低,細(xì)胞凋亡明顯減少[18]。Chau等發(fā)現(xiàn)CV-B3的3C蛋白裂解宿主細(xì)胞的eIF4GI,導(dǎo)致細(xì)胞翻譯過程受到抑制,最終細(xì)胞發(fā)生形態(tài)改變,皺縮等形式的凋亡[19]。Li等研究表明當(dāng)EV-71感染宿主細(xì)胞時(shí),3C蛋白能裂解PinX1蛋白中Q50-G51的特異性位 點(diǎn),PinX1表達(dá)降低可使細(xì)胞DNA損傷,增加細(xì)胞凋亡的易感性[20]。與上述不同的是,Sarah等發(fā)現(xiàn)HRV-16的3C蛋白和caspase 8能裂解外在凋亡途徑的關(guān)鍵中間體,即RIPK1。二者裂解RIPK1的位點(diǎn)和功能不同,caspase 8裂解RIPK1產(chǎn)生一個(gè)38 kD的蛋白,促進(jìn)細(xì)胞發(fā)生早期凋亡,而3C蛋白進(jìn)一步裂解38 kD的蛋白產(chǎn)生C末端23 kD的片段,抑制了凋亡的進(jìn)一步發(fā)展[21]。
3.3逃避免疫應(yīng)答 病毒感染宿主后,宿主啟動(dòng)固有免疫應(yīng)答,如產(chǎn)生干擾素(interferon,IFN)和免疫因子抵御感染,而病毒通過一些策略逃避固有免疫應(yīng)答的清除。研究表明3C蛋白通過抑制IFN的產(chǎn)生和核轉(zhuǎn)錄因子-κB(NF-κB)通路逃避免疫應(yīng)答(圖2)。Rui等研究發(fā)現(xiàn)CV-A6、CV-A16、和EV-D68等小RNA病毒感染細(xì)胞,3C蛋白能與黑色素瘤分化相關(guān)基因(Melanoma differentiation-associated gene 5,MDA5)、維甲酸誘導(dǎo)基因-I(Retinoic acid-inducible gene I,RIG-I)相互作用,破壞了RIG-I 和MAD5與通路下游線粒體抗病毒信號(hào)蛋白(mitochondrial antiviral signaling protein,MAVS)的結(jié)合,從而使MDA5、RIG-I介導(dǎo)的IFN I的產(chǎn)生受到抑制。3C蛋白還可抑制IRF3的磷酸化,從而抑制IFN I的產(chǎn)生[22]。Lei等研究表明EV-71的 3C蛋白與RIG-I的N端相互作用,抑制RIG-I與MAVS形成復(fù)合物,同時(shí)3C蛋白與β干擾素TIR結(jié)構(gòu)域銜接蛋白 (TIR-domain-containing adaptor inducing interferon-β, TRIF)相互作用后誘導(dǎo)TRIF裂解,3C蛋白還可直接裂解IRF7 Q189和S190之間的位點(diǎn),最終上述現(xiàn)象都抑制IFN I產(chǎn)生[23-25]。
NF-κB通路對(duì)于宿主抵抗病毒感染也至關(guān)重要。病毒感染后宿主可通過激活NF-κB通路,刺激機(jī)體產(chǎn)生抗病毒相關(guān)的免疫因子與細(xì)胞因子。3C蛋白可通過裂解TRAF家族與轉(zhuǎn)化生長因子激酶1(transforming growth factor-βactivating kinase 1,TAK1)、TRIF、NF-κB相關(guān)激活因子(TRAF family member-associated NF-κB activator,TANK)抑制NF-κB通路活性。Lei等研究發(fā)現(xiàn)EV-71的3C蛋白裂解TAK1、TAK1結(jié)合蛋白1(TAK1 binding protein 1,TAB1)、TAB2和TAB3,破壞TAK1與TAB1、TAB2和TAB3形成復(fù)合物,從而抑制TAK 1復(fù)合物對(duì)NF-κB通路的激活和細(xì)胞因子的產(chǎn)生[26]。Xiang等研究表明在EV-D68中,作為半胱氨酸酶的3C蛋白可裂解TRIF的312和653位點(diǎn),而導(dǎo)致TRIF失活。失活的TRIF抑制對(duì)NF-κB通路的激活,最終抑制宿主的免疫應(yīng)答[27]。Huang等人研究EMCV發(fā)現(xiàn)3C蛋白能裂解TANK 蛋白的197和291谷氨酰胺位點(diǎn)。TANK即TRAF家族與NF-κB相關(guān)激活因子(TRAF family member-associated NF-κB activator,TANK),是NF-κB的關(guān)鍵調(diào)控因子。3C蛋白對(duì)TANK的裂解破壞這種抑制作用,可抑制TRAF-6介導(dǎo)的NF-κB通路的激活,這是一種新的小RNA病毒逃避宿主免疫的策略[28]。總而言之,小RNA病毒通過3C蛋白逃避免疫應(yīng)答較為復(fù)雜,對(duì)3C蛋白的功能的探索有助于進(jìn)一步闡明小RNA病毒的致病機(jī)制。
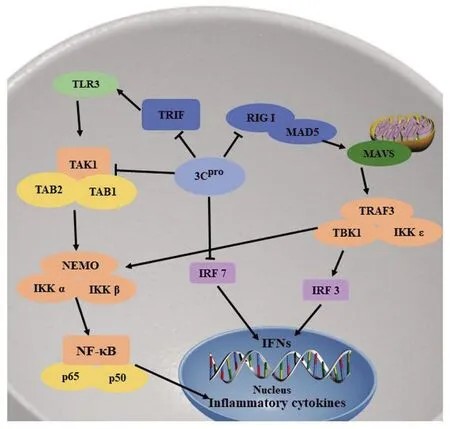
圖2 小RNA 病毒3C蛋白調(diào)控免疫應(yīng)答的機(jī)制Fig.2 Mechanisms of picornavirus 3C protein regulate the immune response
4 展 望
3C蛋白在小RNA病毒自身復(fù)制和與宿主的相互作用中發(fā)揮了重要作用。目前,3C蛋白抑制劑的研究是抗病毒藥物的熱點(diǎn),研究小RNA病毒3C蛋白的功能,不僅能為深入研究小RNA病毒的致病機(jī)制奠定基礎(chǔ),還可為研發(fā)病毒疫苗以及抗病毒藥物提供新的思路。
利益沖突:無
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
哲學(xué)評(píng)論(2021年2期)2021-08-22 01:53:34
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
影視與戲劇評(píng)論(2016年0期)2016-11-23 05:26:01
