棘球絳蟲(chóng)和其它寄生性絳蟲(chóng)乙酰膽堿酯酶和煙堿型乙酰膽堿受體作為潛在藥物靶點(diǎn)的研究進(jìn)展
2019-02-28 07:02:17劉川川樊海寧格日力
中國(guó)人獸共患病學(xué)報(bào) 2019年12期
劉川川,樊海寧,馬 蘭,格日力
棘球蚴病又稱(chēng)包蟲(chóng)病,是由多房棘球絳蟲(chóng)和(或)細(xì)粒棘球絳蟲(chóng)的幼蟲(chóng)棘球蚴寄生于機(jī)體所導(dǎo)致的慢性人獸共患疾病,其成蟲(chóng)主要寄生在狐貍、狗的腸道,經(jīng)糞便排出的蟲(chóng)卵具有傳染性,人為其中間宿主。包蟲(chóng)病可以在不同的宿主器官如肝臟、肺、甚至是心臟和腦中寄生。目前,尚無(wú)有效的預(yù)防棘球蚴病的疫苗[1],并且治療主要依賴(lài)于阿苯達(dá)唑,該藥通過(guò)干擾蟲(chóng)體的能量代謝發(fā)揮作用。但ABZ因其腸道吸收率低,且必須在肝臟轉(zhuǎn)化為砜類(lèi)才能發(fā)揮作用,對(duì)肝臟的副作用較明顯。因此,探索治療棘球蚴病新的治療靶點(diǎn)是十分必要的。絳蟲(chóng)、吸蟲(chóng)等寄生蟲(chóng)體內(nèi)缺乏循環(huán)系統(tǒng)和內(nèi)分泌腺體,而神經(jīng)系統(tǒng)是產(chǎn)生調(diào)節(jié)運(yùn)動(dòng)、生長(zhǎng)發(fā)育等信息物質(zhì)的關(guān)鍵結(jié)構(gòu)[2]。棘球絳蟲(chóng)等蠕蟲(chóng)寄生蟲(chóng)依賴(lài)于其神經(jīng)肌肉組織中的快速突觸傳遞,與外界環(huán)境相互作用并對(duì)其作出反應(yīng)[3]。因此,研究者希望通過(guò)特異性阻斷寄生蟲(chóng)神經(jīng)遞質(zhì)來(lái)達(dá)到驅(qū)除寄生蟲(chóng)的目的。目前,已經(jīng)上市銷(xiāo)售的一些驅(qū)蟲(chóng)藥(如左旋咪唑、噻嘧啶、莫奈太爾和敵百蟲(chóng)等)主要作用于線蟲(chóng)神經(jīng)肌肉系統(tǒng)上的nAchR以及由此引發(fā)的離子通道活性改變[3-5]。神經(jīng)肌肉活動(dòng)的抑制可能導(dǎo)致肌肉功能和基本活動(dòng)的喪失,包括宿主附著、進(jìn)食、交配以及信息的及時(shí)傳遞,從而干擾寄生蟲(chóng)的寄生和生長(zhǎng),導(dǎo)致寄生蟲(chóng)在宿主體內(nèi)被殺滅,效果顯著。而針對(duì)棘球絳蟲(chóng)和其它寄生性絳蟲(chóng)nAchR和AchE的藥物研發(fā)仍處于空白。
1 棘球絳蟲(chóng)的神經(jīng)系統(tǒng)
絳蟲(chóng)神經(jīng)結(jié)構(gòu)周?chē)狈γ黠@的鞘層或明確的邊界,相對(duì)與脊椎動(dòng)物結(jié)構(gòu)較為低等。棘球絳蟲(chóng)中樞神經(jīng)系統(tǒng)由2部分構(gòu)成[3,6]:第一部分是大腦神經(jīng)節(jié)和神經(jīng)循環(huán),吸盤(pán)的神經(jīng)支配以及與周?chē)窠?jīng)系統(tǒng)相連的縱向神經(jīng)索起源于該層;第二部分是一個(gè)旋轉(zhuǎn)的神經(jīng)環(huán)和它的分支組成,貫穿于蟲(chóng)體頭節(jié)以外的節(jié)片,主要分布于寄生蟲(chóng)的皮層、肌肉以及生殖系統(tǒng)發(fā)揮調(diào)節(jié)作用[7]。棘球絳蟲(chóng)體內(nèi)缺乏內(nèi)分泌腺體以及完整的循環(huán)系統(tǒng),神經(jīng)系統(tǒng)是產(chǎn)生激素、神經(jīng)遞質(zhì)以及調(diào)節(jié)肽的唯一結(jié)構(gòu),對(duì)棘球絳蟲(chóng)的生長(zhǎng)、發(fā)育、生殖至關(guān)重要。棘球絳蟲(chóng)的突觸結(jié)構(gòu)中也以突觸小泡作為物質(zhì)運(yùn)輸?shù)男问剑渫挥|結(jié)構(gòu)表現(xiàn)出比高等動(dòng)物更多樣的方式[8-9]。基于此,人們希望通過(guò)特異性阻斷神經(jīng)遞質(zhì)、神經(jīng)肽等物質(zhì)與相關(guān)受體的結(jié)合或影響其降解與合成過(guò)程,達(dá)到對(duì)抗絳蟲(chóng)而不影響機(jī)體的正常功能。
2 棘球絳蟲(chóng)膽堿能神經(jīng)系統(tǒng)
在棘球絳蟲(chóng)膽堿能系統(tǒng)中[6,10],AchE通過(guò)水解乙酰膽堿來(lái)調(diào)節(jié)乙酰膽堿與細(xì)胞乙酰膽堿受體之間的相互作用,使離子通過(guò)電化學(xué)梯度進(jìn)入或離開(kāi)蟲(chóng)體。AchE的主要作用是通過(guò)使神經(jīng)遞質(zhì)Ach的快速水解來(lái)終止膽堿能突觸的傳遞,具有很高的催化活性[11-12]。通過(guò)Na+、K+或Ca2+的內(nèi)流使膜去極化,信號(hào)通過(guò)陽(yáng)離子選擇性乙酰膽堿受體和Ach介導(dǎo)肌肉收縮,對(duì)蠕蟲(chóng)產(chǎn)生明顯的影響,通常表現(xiàn)為麻痹、癱瘓[13]。根據(jù)有關(guān)成年吸蟲(chóng)的研究顯示[14],外源性膽堿能激動(dòng)劑可能導(dǎo)致成年吸蟲(chóng)性麻痹,nAChR在曼氏血吸蟲(chóng)肌纖維的激活過(guò)程中起著重要作用。此外,當(dāng)乙酰膽堿酯酶功能活性受到抑制時(shí),乙酰膽堿酯酶的含量越高,對(duì)nAChR的刺激就越強(qiáng),導(dǎo)致nAChR的脫敏和離子通道的關(guān)閉[10,15]。在哺乳動(dòng)物細(xì)胞的研究中[16-17],17個(gè)已知同源亞基(α1-α10,β1-β4,γ、δ、ε)組裝成不同的nAchR亞型,包括肌肉型nAChRs和神經(jīng)型nAChR亞型。肌肉型nAchR的5種亞基組合形式在發(fā)育階段比較固定(α12β1γδ),能夠通過(guò)與乙酰膽堿結(jié)合而激活,從而導(dǎo)致緩慢的代謝反應(yīng)。隨著棘球絳蟲(chóng)基因組測(cè)序的完成,目前已識(shí)別出棘球絳蟲(chóng)nAchR中有α和β亞基,對(duì)于其它亞基還未見(jiàn)報(bào)道[18]。
3 棘球絳蟲(chóng)nAchRs
目前,只有少部分棘球絳蟲(chóng)nAchR亞基被鑒定出來(lái),其功能也未得到證實(shí)。煙堿乙酰膽堿受體是環(huán)配體門(mén)控離子通道(LGICs)超家族的成員,在神經(jīng)遞質(zhì)乙酰膽堿或外源配體(如尼古丁)結(jié)合后參與信號(hào)轉(zhuǎn)導(dǎo)。nAChRs包含兩個(gè)或多個(gè)α亞基,而α亞基與相鄰亞基組成的二聚體接合面是Ach結(jié)合的關(guān)鍵。每個(gè)nAChR亞基的跨膜域由四個(gè)螺旋跨膜結(jié)構(gòu)組成(M1-M4)[17]。nAchR各亞基的M2結(jié)構(gòu)域形成離子通道,該離子通道僅在Ach結(jié)合引起的變構(gòu)構(gòu)象變化時(shí)打開(kāi)。該通道對(duì)Na+和K+具有較高的滲透性,Ca2+約占其總滲透率的2.5%[19]。Ach與nAChR亞基的配體結(jié)合域(胞外部分)結(jié)合,活化的nAChR導(dǎo)致五聚體蛋白的結(jié)構(gòu)改變,導(dǎo)致配體門(mén)控離子通道的打開(kāi)。隨著對(duì)棘球絳蟲(chóng)測(cè)序的完成,目前所發(fā)現(xiàn)的棘球絳蟲(chóng)nAchR受體亞型仍然較少,但在參與哺乳動(dòng)物神經(jīng)信號(hào)傳遞的關(guān)鍵亞基(如α4β2)在棘球絳蟲(chóng)測(cè)序中卻未能發(fā)現(xiàn);對(duì)已發(fā)現(xiàn)亞基的功能研究仍未進(jìn)一步研究。棘球絳蟲(chóng)跨膜nAchR受體亞基胞外段也存在由13個(gè)氨基酸組成的特征性半胱氨酸環(huán)結(jié)構(gòu)(圖1),提示棘球絳蟲(chóng)nAChRs是環(huán)配體門(mén)控離子通道超家族的成員;同時(shí),該環(huán)結(jié)構(gòu)在參與Ach和nAchR的結(jié)構(gòu)結(jié)合和寄生蟲(chóng)的神經(jīng)傳遞有重要作用[20-21]。通過(guò)對(duì)已發(fā)現(xiàn)亞基模體結(jié)構(gòu)分析,發(fā)現(xiàn)棘球絳蟲(chóng)nAchR亞基中存在著高度保守的門(mén)控離子通道配體結(jié)合域和跨膜域(表1)。最近研究者提出了G蛋白與神經(jīng)元nAChRs之間存在直接耦合,這導(dǎo)致了一種新的假設(shè)[22],即神經(jīng)元煙堿乙酰膽堿受體的主要胞內(nèi)環(huán)中的某些氨基酸構(gòu)成了G蛋白的結(jié)合位點(diǎn),與G蛋白受體結(jié)合可以調(diào)節(jié)細(xì)胞nAChRs的活性和信號(hào)傳導(dǎo),這一過(guò)程在結(jié)構(gòu)上與配體門(mén)控離子通道的打開(kāi)有關(guān),并通過(guò)相關(guān)實(shí)驗(yàn)得到證實(shí)。
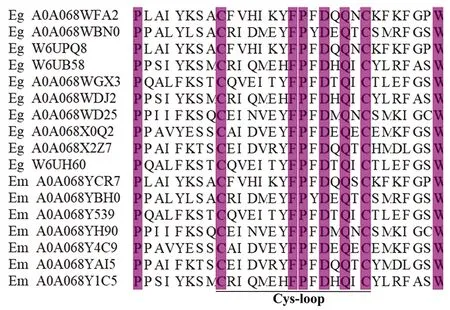
圖1 棘球絳蟲(chóng)跨膜nAchR受體C-FP-D-Q-C特征性結(jié)構(gòu)Fig.1 Characteristic structure of transmembrane nAchR receptor C-FP-D-Q-C of Echinococcus
5-羥色胺(5-hydroxytryptamine,5-HT)是一種抑制性神經(jīng)遞質(zhì),為G蛋白偶聯(lián)受體家族成員,廣泛分布哺乳動(dòng)物組織中,尤其是大腦皮層及神經(jīng)突觸內(nèi)。G蛋白與受體結(jié)合可以調(diào)控nAchR的活性。研究發(fā)現(xiàn)[23]絳蟲(chóng)5-HT受體在棘球絳蟲(chóng)原頭節(jié)和成蟲(chóng)以及包蟲(chóng)囊壁中均有表達(dá),該受體在調(diào)控寄生蟲(chóng)神經(jīng)肌肉控制方面起主導(dǎo)作用,同時(shí)在調(diào)控發(fā)育和繁殖方面也有重要作用。5-HT 在無(wú)脊椎動(dòng)物的各種生命活動(dòng)中同樣發(fā)揮了重要的作用,很多的5-HT受體已經(jīng)從不同物種中克隆出來(lái),但克隆出的寄生蟲(chóng)5-HT受體與哺乳動(dòng)物5-HT的親和力都很低,表明非脊椎動(dòng)物5-HT受體與哺乳動(dòng)物受體結(jié)構(gòu)存在差異,基于此篩選對(duì)寄生蟲(chóng)5-HT 受體高選擇性,而對(duì)非靶標(biāo)生物無(wú)害的藥物是新型殺蟲(chóng)劑的另一個(gè)研制目標(biāo)。早在1994年[24-25],研究者采用免疫細(xì)胞化學(xué)技術(shù)鑒定了棘球絳蟲(chóng)神經(jīng)系統(tǒng)5-HT受體的定位和分布,神經(jīng)細(xì)胞體和神經(jīng)纖維對(duì)5-HT免疫反應(yīng)發(fā)生在成對(duì) 的外側(cè)神經(jīng)節(jié)、后側(cè)神經(jīng)節(jié)和星狀神經(jīng)節(jié),它們的連合部和頭節(jié)中的神經(jīng)環(huán),以及遍及蠕蟲(chóng)全身向后延伸的十條縱向神經(jīng)索中,同時(shí)發(fā)現(xiàn)其分布與乙酰膽堿酯酶的分布相一致,但5-HT受體表現(xiàn)出更廣泛的分布范圍。之后研究者采用間接免疫熒光技術(shù)鑒定在原頭節(jié)中5-HT受體和神經(jīng)肽的定位和分布[26],其在中樞神經(jīng)環(huán)、環(huán)狀神經(jīng)和支配吸盤(pán)的神經(jīng)叢中的定位與成蟲(chóng)相同。考慮到5-HT受體在寄生蟲(chóng)生物學(xué)中的重要作用,以及5-HT受體相對(duì)于哺乳動(dòng)物的差異,5-HT受體被認(rèn)為是抗包蟲(chóng)病的藥物靶點(diǎn)。在原頭節(jié)培養(yǎng)中添加5-HT后,短時(shí)間內(nèi)其運(yùn)動(dòng)能力明顯增強(qiáng);但在加入5-HT轉(zhuǎn)運(yùn)體抑制劑西酞普蘭后,消除了5-HT誘導(dǎo)的運(yùn)動(dòng)性增加,而5-HT受體拮抗劑昂丹司瓊卻無(wú)明顯作用,表明西酞普蘭能抑制5-HT轉(zhuǎn)運(yùn)蛋白,也證明了其受體結(jié)構(gòu)與哺乳動(dòng)物的差異[27];同時(shí)原頭節(jié)用含5-HT培養(yǎng)基孵育幾天后誘導(dǎo)向中絳期分化,而中絳期的幼蟲(chóng)在宿主體內(nèi)能以無(wú)性繁殖的方式產(chǎn)生原頭蚴,表明5-HT在棘球蚴的發(fā)育過(guò)程中具有一定的神經(jīng)功能和促發(fā)育作用[27]。但到目前為止,在棘球絳蟲(chóng)神經(jīng)肌肉連接處,5-HT的確切作用機(jī)制尚不清楚。在其它無(wú)脊椎動(dòng)物的研究中發(fā)現(xiàn)[23,28],5-HT的應(yīng)用增加了可釋放突觸小泡的總突觸池的大小,以響應(yīng)突觸前膜末端的動(dòng)作電位。可以假設(shè),在添加5-HT后,類(lèi)似的作用機(jī)制可能會(huì)以類(lèi)似的方式作用于棘球絳蟲(chóng)的神經(jīng)肌肉連接處,但還需要更多的實(shí)驗(yàn)來(lái)驗(yàn)證這一假設(shè)。
然而,乙酰膽堿和寄生蟲(chóng)的乙酰膽堿受體結(jié)合如何導(dǎo)致離子通道的打開(kāi)仍有待研究。根據(jù)哺乳動(dòng)物的研究以及有關(guān)棘球絳蟲(chóng)的研究,預(yù)測(cè)棘球絳蟲(chóng)潛在膽堿能信號(hào)通路,如圖2。
4 棘球絳蟲(chóng)AchE
早期研究發(fā)現(xiàn)[29],一種特殊的酶存在于棘球絳蟲(chóng)頭節(jié)勻漿和生發(fā)囊中,其對(duì)乙酰膽堿的水解率是丁酰膽堿的45倍,其活性可被毒扁豆堿和乙基四磷酸酯(有機(jī)磷殺蟲(chóng)劑)抑制。西班牙科學(xué)家實(shí)驗(yàn)發(fā)現(xiàn)[30],細(xì)粒棘球蚴會(huì)釋放乙酰膽堿酯酶等多種酶類(lèi),這些酶可以穿過(guò)生發(fā)層和囊壁,進(jìn)入宿主組織,破壞宿主產(chǎn)生的乙酰膽堿;棘球蚴產(chǎn)生這些酶可能是對(duì)抗棘球蚴病化療藥物的有吸引力靶標(biāo),可以通過(guò)藥物抑制酶的活性導(dǎo)致乙酰膽堿累積,阻塞神經(jīng)功能。這些研究都證實(shí)了棘球絳蟲(chóng)有乙酰膽堿酯酶(AchE)的存在。乙酰膽堿和乙酰膽堿酯酶抑制劑可以放松蠕蟲(chóng)的肌肉組織,減少寄生蟲(chóng)的蠕動(dòng),最終導(dǎo)致癱瘓[31]。
大多數(shù)單克隆抗體僅能與寄生蟲(chóng)的乙酰膽堿酯酶相互作用,而不與脊椎動(dòng)物宿主乙酰膽堿酯酶發(fā)生作用,表明宿主與寄生蟲(chóng)酶具有不同的表位結(jié)構(gòu)及空間構(gòu)象[32]。因而針對(duì)棘球絳蟲(chóng)及其它寄生蟲(chóng)AchE特異性的抗原表位是藥物設(shè)計(jì)和疫苗開(kāi)發(fā)的靶點(diǎn),同時(shí)藥物可能表現(xiàn)出對(duì)脊椎動(dòng)物較低的毒性。棘球絳蟲(chóng)等蠕蟲(chóng)乙酰膽堿酯酶分布于寄生蟲(chóng)的表面,可用于評(píng)估抗蠕蟲(chóng)藥物的有效性[33]。研究者使用鈣調(diào)素拮抗劑處理棘球蚴引起棘球蚴表層被膜明顯改變,頭鉤脫落,并且在其培養(yǎng)上清液中檢測(cè)出AchE的表達(dá)明顯增加,但有趣的是在未處理組培養(yǎng)上清液中也檢測(cè)到AchE的存在,對(duì)此一個(gè)可能的解釋是由于AchE與細(xì)胞膜上成分的相互作用的方式導(dǎo)致很容易被釋放到培養(yǎng)基中,是一種獨(dú)特的附著部分[33],同時(shí)也表明AchE可能也分布于棘球蚴表面。研究者使用白藜蘆醇處理絳蟲(chóng)后[6],導(dǎo)致吸盤(pán)松動(dòng)并出現(xiàn)麻痹,其乙酰膽堿酯酶的表達(dá)量下降,表明白藜蘆醇可能通過(guò)抑制其乙酰膽堿酯酶的活性而導(dǎo)致絳蟲(chóng)體壁肌肉松弛。
表1 棘球絳蟲(chóng)煙堿乙酰膽堿受體保守結(jié)構(gòu)分析
Tab.1 Conserved structure analysis of nicotinic acetylcholine receptors in echinococcus
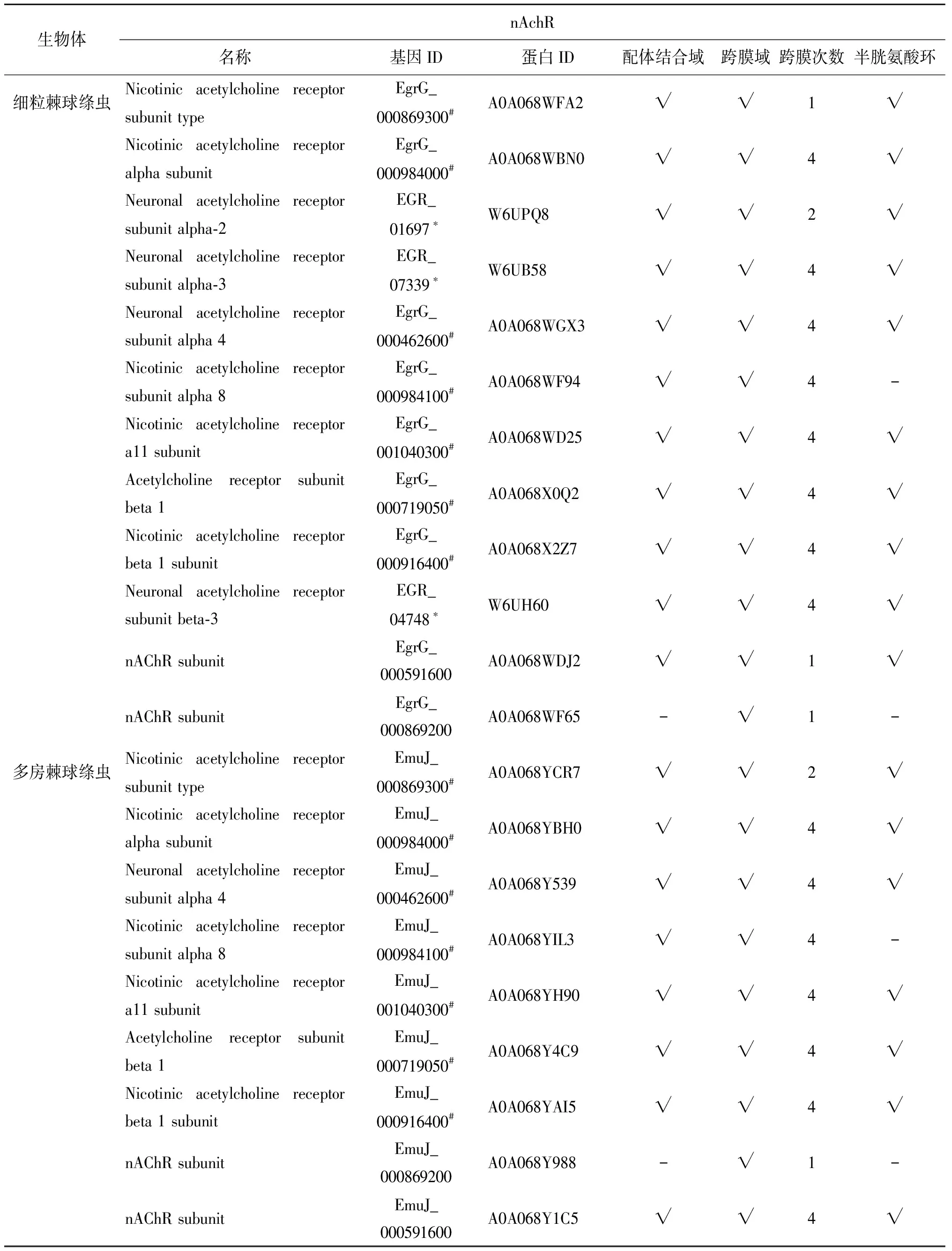
生物體nAchR名稱(chēng)基因ID蛋白ID配體結(jié)合域跨膜域跨膜次數(shù)半胱氨酸環(huán)細(xì)粒棘球絳蟲(chóng)Nicotinic acetylcholine receptor subunit typeEgrG_000869300#A0A068WFA2√√1√N(yùn)icotinic acetylcholine receptor alpha subunitEgrG_000984000#A0A068WBN0√√4√N(yùn)euronal acetylcholine receptor subunit alpha-2EGR_01697?W6UPQ8√√2√N(yùn)euronal acetylcholine receptor subunit alpha-3EGR_07339?W6UB58√√4√N(yùn)euronal acetylcholine receptor subunit alpha 4EgrG_000462600#A0A068WGX3√√4√N(yùn)icotinic acetylcholine receptor subunit alpha 8EgrG_000984100#A0A068WF94√√4-Nicotinic acetylcholine receptor a11 subunitEgrG_001040300#A0A068WD25√√4√Acetylcholine receptor subunit beta 1EgrG_000719050#A0A068X0Q2√√4√N(yùn)icotinic acetylcholine receptor beta 1 subunitEgrG_000916400#A0A068X2Z7√√4√N(yùn)euronal acetylcholine receptor subunit beta-3EGR_04748?W6UH60√√4√nAChR subunitEgrG_000591600A0A068WDJ2√√1√nAChR subunitEgrG_000869200A0A068WF65-√1-多房棘球絳蟲(chóng)Nicotinic acetylcholine receptor subunit typeEmuJ_000869300#A0A068YCR7√√2√N(yùn)icotinic acetylcholine receptor alpha subunitEmuJ_000984000#A0A068YBH0√√4√N(yùn)euronal acetylcholine receptor subunit alpha 4EmuJ_000462600#A0A068Y539√√4√N(yùn)icotinic acetylcholine receptor subunit alpha 8EmuJ_000984100#A0A068YIL3√√4-Nicotinic acetylcholine receptor a11 subunitEmuJ_001040300#A0A068YH90√√4√Acetylcholine receptor subunit beta 1EmuJ_000719050#A0A068Y4C9√√4√N(yùn)icotinic acetylcholine receptor beta 1 subunitEmuJ_000916400#A0A068YAI5√√4√nAChR subunit EmuJ_000869200A0A068Y988-√1-nAChR subunitEmuJ_000591600A0A068Y1C5√√4√
#基因ID來(lái)源于http://www.genedb.org/Homepage;*基因ID來(lái)源于https://www.ncbi.nlm.nih.gov/;蛋白ID來(lái)源于https://www.uniprot.org/
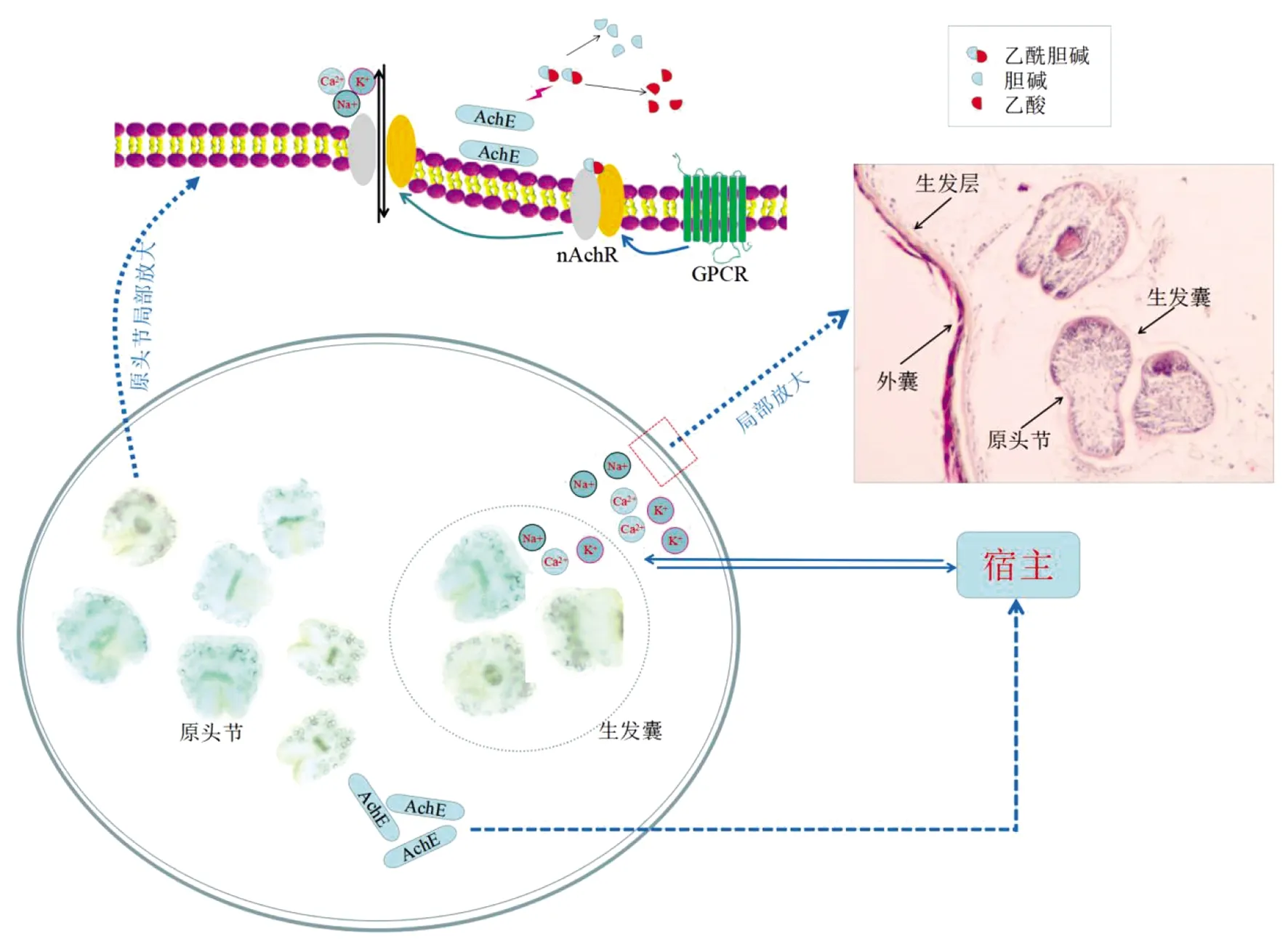
圖2 預(yù)測(cè)棘球絳蟲(chóng)膽堿能信號(hào)途徑Fig.2 predicted the cholinergic signal pathway of echinococcus
表2 棘球絳蟲(chóng)AchE基本信息
Tab.2 Basic information of AchE in echinococcus

生物體AchE基因[11]蛋白ID信號(hào)肽/亞細(xì)胞定位細(xì)粒棘球絳蟲(chóng) EgrG_001075400#A0A068WK19MRIPLSSVLILLAPTALA/胞外EGR_04525?EgrG_000732400#U6JKM7MGHWEVVMFILALEILPTLT/胞內(nèi)K4EPC5MGHWEVVMFILALEILPTLT/胞外EGR_07877?W6UGL0無(wú)/胞外EGR_07474?W6UHU3MLRPLGCLAYLGLFLSECLA/胞外多房棘球絳蟲(chóng)EmuJ_001075400#A0A068YES8MRIPLSCVLILLVPTALA/胞外EmuJ_000732400#A0A068Y5F4MGHREVVMFILALEILPTLT/胞內(nèi)
#基因ID來(lái)源于http://www.genedb.org/Homepage;*基因ID來(lái)源于https://www.ncbi.nlm.nih.gov/;蛋白ID來(lái)源于https://www.uniprot.org/
5 其它寄生蟲(chóng)絳蟲(chóng)AchE和nAchRs研究進(jìn)展
近年來(lái),隨著對(duì)多種棘球絳蟲(chóng)基因組的研究,對(duì)寄生性絳蟲(chóng)的寄生機(jī)制及潛在的作用靶點(diǎn)的識(shí)別提供了理論依。但目前對(duì)于棘球絳蟲(chóng)及其它寄生性絳蟲(chóng)相關(guān)受體的鑒定及功能研究相對(duì)缺乏,而在吸蟲(chóng)和線蟲(chóng)相關(guān)受體的研究取得了較大的進(jìn)步。早期的研究表明[34],在秀麗隱桿線蟲(chóng)體內(nèi)乙酰膽堿是控制運(yùn)動(dòng)的主要興奮性神經(jīng)遞質(zhì),而AchE是調(diào)節(jié)的關(guān)鍵酶,并證實(shí)了四種AchE基因可以編碼不同的酶。最近的研究表明,線蟲(chóng)分泌乙酰膽堿酯酶的功能之一可能是改變宿主細(xì)胞因子環(huán)境,抑制M2巨噬細(xì)胞的發(fā)育,從而增進(jìn)寄生蟲(chóng)的生存。在成年肝片吸蟲(chóng)腦神經(jīng)細(xì)胞胞體和腦神經(jīng)節(jié)的神經(jīng)束中也檢測(cè)到乙酰膽堿酯酶的存在[35],進(jìn)一步研究還發(fā)現(xiàn)乙酰膽堿酯酶反應(yīng)產(chǎn)物與含有透明球狀突觸泡的突觸末端相關(guān),反應(yīng)產(chǎn)物定位于突觸前端和突觸后端對(duì)位區(qū)的突觸接觸部位,這種酶的分布與它作為膽堿能傳遞降解酶的功能一致[33,35];同時(shí),肝片吸蟲(chóng)能影響機(jī)體膽堿和嘌呤的代謝,進(jìn)而影響疾病的發(fā)病機(jī)制和進(jìn)程[36]。在血吸蟲(chóng)中發(fā)現(xiàn)AchE存在于血吸蟲(chóng)表面,并能調(diào)節(jié)葡萄糖代謝[37-38];研究者使用RNA干擾技術(shù)敲除AchE在血吸蟲(chóng)成蟲(chóng)和蟲(chóng)卵中的表達(dá),這導(dǎo)致了它的轉(zhuǎn)錄減少,影響了膽堿能信號(hào)傳導(dǎo)和葡萄糖攝取的相關(guān)重要基因的表達(dá)[39]。在大腸桿菌中表達(dá)重組AChE后免疫接種小鼠,接種疫苗組的雄性蠕蟲(chóng)數(shù)量、肝肉芽腫密度和成熟腸卵數(shù)量均較對(duì)照組減少,表明乙酰膽堿酯酶在雄性蠕蟲(chóng)的代謝中起著重要作用,并間接影響雌性的繁殖力,導(dǎo)致未成熟卵的釋放量增加和肝肉芽腫體積減小[39]。新近研究發(fā)現(xiàn)[40],聚吡啶釕(II)能抑制血吸蟲(chóng)AchE活性,處理后對(duì)葡萄糖的吸收能力明顯下將,糖原儲(chǔ)存明顯減少。這些研究都進(jìn)一步證明了AchE在寄生蟲(chóng)的生長(zhǎng)發(fā)育、運(yùn)動(dòng)及影響宿主方面的中藥作用。在寄生蟲(chóng)的不同發(fā)育階段[41],乙酰膽堿酯酶的分子形式和功能活性的存在較大的差異,在成蟲(chóng)階段表現(xiàn)更為突出,這種變化可能反應(yīng)了復(fù)雜神經(jīng)系統(tǒng)的存在,尤其在寄生蟲(chóng)發(fā)育的晚期是必須的。
AchR作為潛在驅(qū)蟲(chóng)藥物的靶點(diǎn)越來(lái)越受到關(guān)注,AchR是膽堿能激動(dòng)劑或拮抗藥物的主要靶點(diǎn),代表藥物有左旋咪唑、噻嘧啶、三苯雙脒和得曲恩特[42]。寄生蟲(chóng)體內(nèi)存在多種類(lèi)型的nAchR,只有少部分受體亞型被鑒定出來(lái),各受體亞型的功能仍不確定,并且針對(duì)nAchR不同亞基藥物聯(lián)合使用表現(xiàn)出比單藥使用更加明顯的療效[43]。秀麗隱桿線蟲(chóng)體內(nèi)已被證明具有兩種刺激性AchR[42,44]:一種對(duì)左旋咪唑敏感,由3個(gè)α亞基(LEV-8、Cel-UNC-38、UNC-63)和2個(gè)非α亞基(LEV-1和UNC-29)組成的五聚體;另一種對(duì)尼古丁敏感,由acr-8和acr-16組成,acr-8的缺失對(duì)信號(hào)的傳遞無(wú)影響,而acr-16亞基的基因消融可以消除體壁肌肉中的非左旋膽堿能反應(yīng)。從豬蛔蟲(chóng)中分離出的3種AchR亞型為N型、L型和B型,介導(dǎo)蛔蟲(chóng)體壁肌肉的收縮,每種亞型對(duì)膽堿能驅(qū)蟲(chóng)劑的敏感性不同[45]。研究者在爪蟾卵母細(xì)胞中表達(dá)編碼蛔蟲(chóng)unc-29和unc-38亞基發(fā)現(xiàn),兩亞基的比例會(huì)影響驅(qū)蟲(chóng)藥物的敏感性[46]。通過(guò)對(duì)寄生線蟲(chóng)模型的4個(gè)nAchR亞型的鑒定,證明每個(gè)亞型都獲得了獨(dú)特的功能特征,這些功能是基于轉(zhuǎn)基因秀麗隱桿線蟲(chóng)的表型研究和非洲爪蟾卵母細(xì)胞受體重組的電生理學(xué)基礎(chǔ)上獲得的[47]。基于秀麗隱桿線蟲(chóng)的研究發(fā)現(xiàn)[47],寄生蟲(chóng)可以通過(guò)改變nAchR受體的組合形式,導(dǎo)致其功能發(fā)生差異,藥物因作用靶點(diǎn)結(jié)構(gòu)改變而失效。
大量的研究表明,寄生蟲(chóng)nAchR亞基存在不同的亞型,不同的亞型執(zhí)行特有的功能,同時(shí)各亞型在組合成多聚體時(shí)能發(fā)生構(gòu)象改變,從而導(dǎo)致寄生蟲(chóng)對(duì)藥物敏感性下降和耐藥的產(chǎn)生。
6 小 結(jié)
抗包蟲(chóng)病藥物引起的耐藥性、低生物利用度和水溶性差、嚴(yán)重的副作用限制了其使用。因此,迫切需要研制治療包蟲(chóng)病感染的新型藥物。AchE和nAchR在寄生性蠕蟲(chóng)神經(jīng)肌肉和其他信號(hào)系統(tǒng)中起著重要作用,是目前市場(chǎng)上銷(xiāo)售的許多驅(qū)蟲(chóng)藥物的靶點(diǎn)。了解AchE和nAchR的分子特征和功能特性以及它們?cè)谥袠猩窠?jīng)系統(tǒng)的分布,將推動(dòng)高效驅(qū)蟲(chóng)劑的研發(fā)進(jìn)程。有關(guān)棘球絳蟲(chóng)的研究還遠(yuǎn)落后于脊椎動(dòng)物,許多功能性分子的作用還未證實(shí)。目前,應(yīng)用RNA干擾技術(shù)在了解寄生蟲(chóng)各亞基功能上取得了一定的進(jìn)步。隨著對(duì)棘球蚴病早期診斷試劑研發(fā)的不斷進(jìn)展[48-49],對(duì)包蟲(chóng)病治療特效藥物的研發(fā)提出了更加嚴(yán)格的考驗(yàn)。
利益沖突:無(wú)
