兒茶酚胺濃度及β1—腎上腺素受體基因多態性與最大有氧能力的關聯性
2019-02-13 13:48:44樂曉卉孫忠芹陳洪魁楊留才史衛紅
安徽師范大學學報(自然科學版) 2019年6期
關鍵詞:差異
樂曉卉, 孫忠芹, 陳洪魁, 楊留才,王 瑾, 仲 衛, 史衛紅, 劉 爽
(1.鹽城工學院 體育學院,江蘇 鹽城 224000;2.鹽城體育運動學校 訓練系,江蘇 鹽城 224000;3.江蘇醫藥職業學院 體育部等,江蘇 鹽城 224005;4.黑龍江佳木斯大學 基礎醫學院,黑龍江 佳木斯 154000)
遺傳因素在運動員選才中的應用已受廣泛重視,目前主要集中在血管緊張素原基因、腺苷-磷酸脫氨酶1、低氧誘導因子1α、α輔肌動蛋白3等基因,而腎上腺素受體基因(adrenergic receptor,AR)的相關研究僅局限在β2和β3,但其主要分別存在于呼吸系統和脂肪組織中,而β1腎上腺素受體基因(β adrenergic receptor,β1AR)主要分布在心肌細胞中[1-4],其通過改變Gs蛋白與β1AR結合部位產生功能效應[5],按照對有氧運動能力起主要作用的骨骼肌來說,心肌供血比呼吸和脂肪作用更大,而檢索PUB-Med和CNKI,未見相關報道。
因此,本研究采用鹽城市的運動員和體育專業的學生作為樣本,比較有氧運動能力與血漿兒茶酚胺濃度的相關性,探討β1AR 49、389位蛋白質的密碼子SNPS及其與有氧運動能力的關聯性,為運動員的基因選才研究提供基礎數據。
1 研究對象與方法
1.1 研究對象
選擇鹽城運動學校、鹽城師范學院體育專業學生為實驗組,對照組為同時期同地區的非體育專業未經任何專業訓練的健康學生,均為漢族,并征得本人或家長同意,所有受試者簽署知情同意書,兩組試驗過程中均無脫落(脫落者刪除),其基線資料分析如表1所示,其年齡、性別、身高和BMI差異無統計學意義(P>0.05),具可比性。
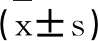
表1 二組基線資料比較表
注:▲與實驗組男相比◆與實驗組女相比
1.2 儀器和試劑
微量高速離心機(北京鼎昊源MicroSmart),生物安全柜(青島海爾HR40-II A2),漩渦混合器(江蘇海門其林貝爾XW-80A),掌上型迷你離心機(海門其林貝爾LX-400),全自動凝膠成像分析系統(ChampGelTM5000),全自動酶標儀(美國BIO-RAD);無核酸酶水、Goldstar Best MasterMix、無水乙醇、血液基因組柱式小提試劑盒和蛋白酶K(均為北京康為世紀),人兒茶酚胺酶聯免疫試劑盒(武漢新啟迪生物科技有限公司),瓊脂糖、電泳緩沖液(上海蘭衛檢驗實驗室)。
1.3 研究方法
1.3.1 運動方式 采用吳劍[6]法,實驗組和對照組均采用德國Ergoline 1200EL有氧功率車,方式為臥式蹬車的遞增定量負荷運動,以起始負荷50W和60r/min的頻率開始蹬車,每3分鐘增加負荷50W,直至力竭,平臥恢復3分鐘。
1.3.2 血樣品采集 所有實驗組和對照組均于運動結束即時抽取外周血3m1,放置至-80℃冰箱凍存。
1.3.3 ELISA法測定血漿中兒茶酚胺含量 根據兒茶酚胺含量測定試劑盒說明書進行操作,結果采用標準液進行倍比稀釋后,同法操作測定,用酶標儀在450nm測定吸光度值,制作標準曲線(如圖1);各樣品同法得到的吸光值,在標準曲線上找出對應的濃度。

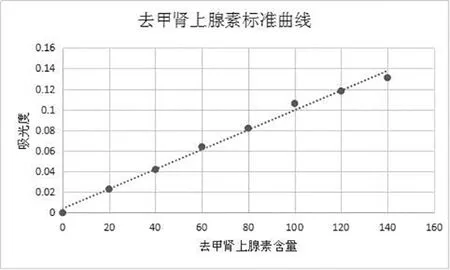
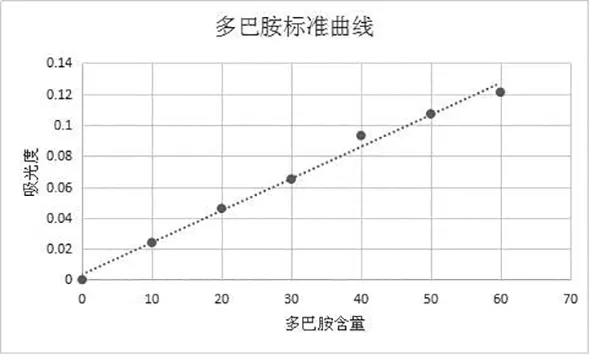
圖1 兒茶酚胺類物質濃度測定標準曲線圖
1.3.4 基因操作 1.3.4.1 DNA提取 取外周靜脈全血EDTA抗凝,分離白細胞,酚/氯仿法提取基因組DNA,利用ND-1000紫外/可見分光光度計測定DNA質量和濃度,并放置在-80℃冰箱凍存。
1.3.4.2 β1AR基因擴增 用Primer 3.0在線軟件設計引物進行PCR-RFLP,β1AR基因49位點上游引物為5’-CCUUUGTTCTUUUUTUTTCC-3’,下游引物為5’-UUCUAUUTUATUUCUA UUTAUC-3’,片段大小562bp;β1-AR基因389位點上游引物為5’-CATCATUUUCUTCTT CACUC-3’,下游引物為5’-TUUUCTTCUAUTTCA CCTUC-3’,片段大小547bp。PCR擴增體系:2×Goldstar Best Master Mix 15μ1,10μM引物2μl,模板100ng,補充ddH2O至總體積30μ1,置ABI2720熱循環儀(美國)中,反應條件:循環1次(99℃10min,98℃30sec),循環38次(60℃45 sec,72℃90sec)。
1.3.4.3 酶切 β-AR基因49位點PCR產物,采用NEB公司(美國)的限制性內切酶Eco O109I在37℃下水浴中酶切8h;β-AR基因389位點PCR產物,采用NEB公司(美國)的限制性內切酶BcgI在37℃下水浴中酶切4h。
1.3.4.4電泳 兩種酶切產物,各取3μ1,以100bp DNA為Marker,5%瓊脂糖凝膠電泳時間45分鐘(電壓90V),然后在紫外線燈下顯色觀察。
1.3.4.5 進行質譜檢測,并收集數據 基質輔助激光解析電離飛行時間質譜(Matrix-assisted laser desorption ionization time of flight mass spectrometry,MALDI-TOF MS)反應;Typer 4.0軟件檢測質譜峰,并根據質譜峰圖判讀各樣本目標位點基因型。
1.4 運動能力檢測
采用高炳宏[7]一口氣接一口氣法,運用心肺功能測試儀(Cortex Biophysik,德國產)測試,每10s連續記錄氣體代謝的各項指標,包括平均體重最大耗氧量(VO2max/kg)、最大二氧化碳排出量(VCO2max)、最大呼吸商(respiratory quotient,RQmax)、最大通氣量(Maximal voluntary ventilation,MVV),最大呼吸頻率(Respiratory frequency,RFmax)、最大氧脈搏(O2Plusemax),最大功率(Pmax),最大運動時間(Tmax)、最大負荷的工作時間(Wmax-T),并測定運動后即刻、運動后3min和恢復后心率。
1.5 數據處理
2 結果
2.1兒茶酚胺類物質濃度比較
實驗組和對照組兒茶酚胺類物質濃度比較如表2所示,運動后腎上腺素、去甲腎上腺素和多巴胺濃度,實驗組均高于對照組,同性別相比,實驗組亦均高于對照組,差異具有統計學意義(P<0.05)。
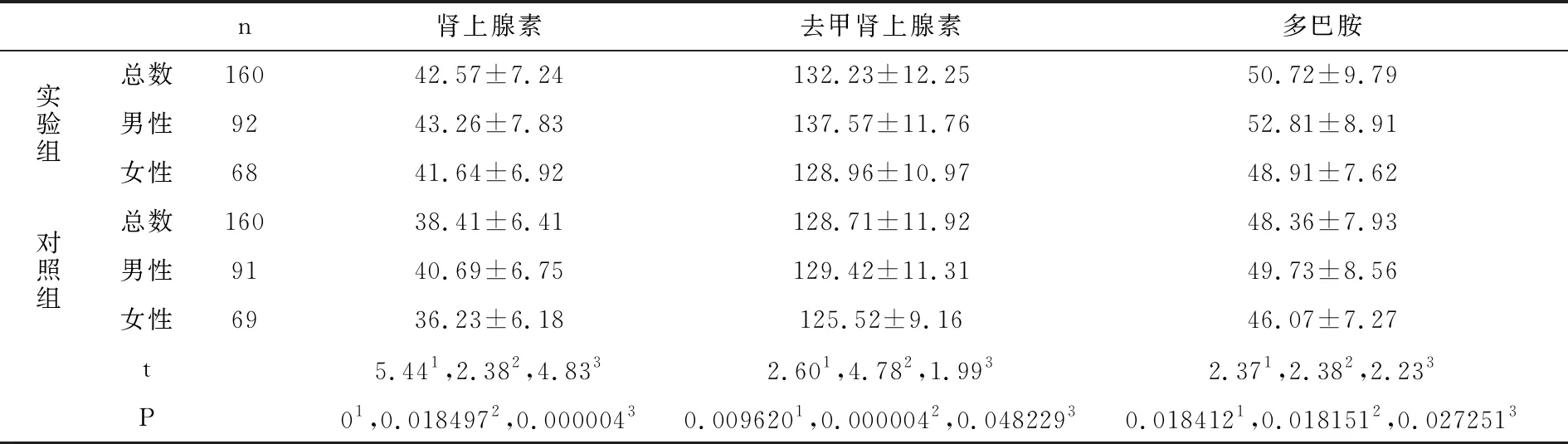
表2 兒茶酚胺類物質濃度比較表(pg/ml,均數±標準差)
注:1總數相比,2男性相比 3女性相比
2.2 β1AR基因PCR擴增檢測結果
根據劉建華[1]的研究,β1AR的Ser49Gly基因多態性:Ser49Ser對應堿基為AA,Ser49Gly對應AG,GIy49Gly對應GG;Arg389G1y基因多態性:Arg389Arg對應堿基為CC,Arg389G1y對應CG,G1y389G1y對應GG。
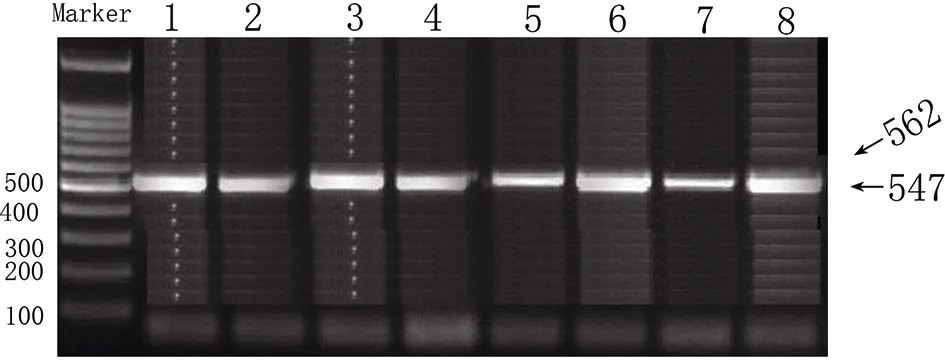
圖2 β1AR基因49位點和389位點PCR-RFLP擴增檢測結果
β1AR基因49位點和389位點PCR擴增目的片段長分別在562bp和547bp,如圖2所示。
2.3 β1AR49和389位點多態性基因型分型
β1AR49基因分布為GG、AG、AA三種,而389位點多態性基因型分布為CC、CG和CC三種,(如圖3、4所示)。
2.4 β1AR基因49位點多態性
使用Eco O109I對β1AR基因49位點酶切可區分出3種基因型:GG型(219,343bp2條條帶)、AG型(562,343,219bp3條條帶)和AA型(562bp1條條帶),見圖5。兩組β1AR基因49位點多態性分析:實驗組和對照組均符合H-W遺傳平衡定律(P>0.05),具有群體代表性。兩組β1AR基因49位點基因型和等位基因頻率比較見表3所示,基因型GG、AG、AA分布的比較,差異無統計學意義(P>0.05),兩組等位基因G、A頻率的比較,差異亦無統計學意義(P>0.05)。
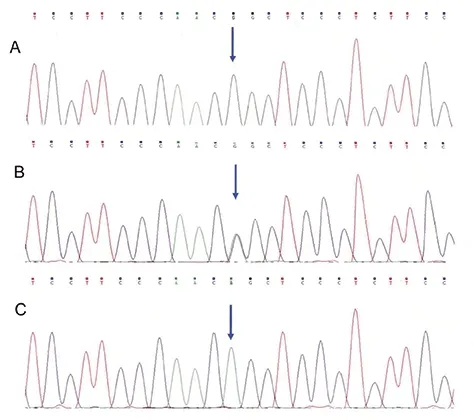
圖3 β1AR基因49位點基因型分布圖
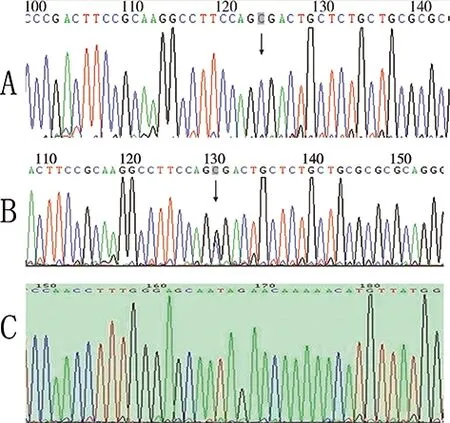
圖4 β1AR基因389位點多態性的測序圖譜
2.5 β1AR基因389位點多態性
使用BcgI對β1AR基因389位點酶切可區分出3種基因型:GG型(547bp1條條帶)、CG型(4,171,342,547bp4條條帶)和CC型(34,171,342bp3條帶),見圖5。兩組β1AR基因389位點多態性分析:實驗組和對照組均符合H-W遺傳平衡定律(P>0.05),具有群體代表性。兩組β1AR基因389位點基因型和等位基因頻率比較見表5所示,實驗組基因型CC和等位基因G、C基因型頻率與對照組相比,差異均具有統計學意義(P<0.05),而兩組CG、GG基因型比較,差異無統計學意義(P>0.05)。
*為實際數和理論數的比較

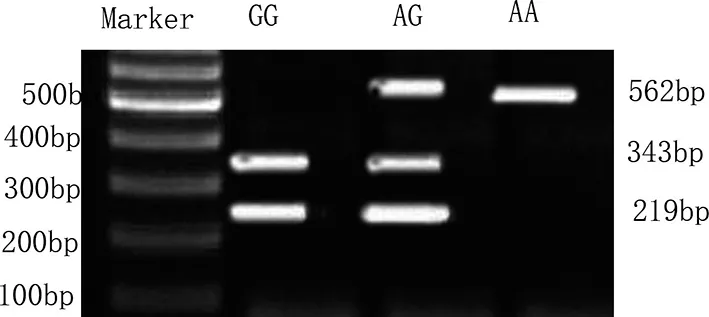
圖5 β1AR基因49和389位點多態性PCR-RFLP電泳結果

表4 兩組β1AR基因389位點基因型和等位基因頻率比較(n/%)
*實際數和理論數比較
2.6 血液兒茶酚胺濃度與β1AR基因多態性的關系
2.6.1 血液腎上腺素濃度與β1AR基因多態性的關系 血液腎上腺素濃度與β1AR基因多態性的關系見表5所示,實驗組的血液腎上腺素濃度與β1AR基因389位點的CC明顯高于對照組,差異均具有顯著統計學意義(P<0.01);而兩組β1AR基因49位點和β1AR基因389位點的CG、GG基因型比較,差異無統計學意義(P>0.05)。
兩組中,49位點的AG基因型的血液腎上腺素濃度明顯高于AA型,差異均具有顯著統計學意義(P<0.01),但與GG基因型無差異,差異無統計學意義(P>0.05)。而389位點的CC基因型的血液腎上腺素濃度明顯高于CG型,差異均具有顯著統計學意義(P<0.01),但與GG基因型無差異,差異無統計學意義(P>0.05)。

表5 血液腎上腺素濃度與β1AR基因多態性的關系
*P<0.01/0.05
2.6.2 血液去甲腎上腺素濃度與β1AR基因多態性的關系 血液去甲腎上腺素濃度與β1AR基因多態性的關系見表6所示,實驗組的血液去甲腎上腺素濃度與β1AR基因389位點的CC明顯高于對照組,差異均具有顯著統計學意義(P<0.01),而兩組β1AR基因49位點和β1AR基因389位點的CG、GG基因型比較,差異無統計學意義(P>0.05)。
兩組中,49位點的AG基因型的血液去甲腎上腺素濃度明顯高于AA型,差異均具有顯著統計學意義(P<0.01),但與GG基因型無差異,差異無統計學意義(P>0.05)。而389位點的基因型的血液去甲腎上腺素濃度無差異,差異無統計學意義(P>0.05)。
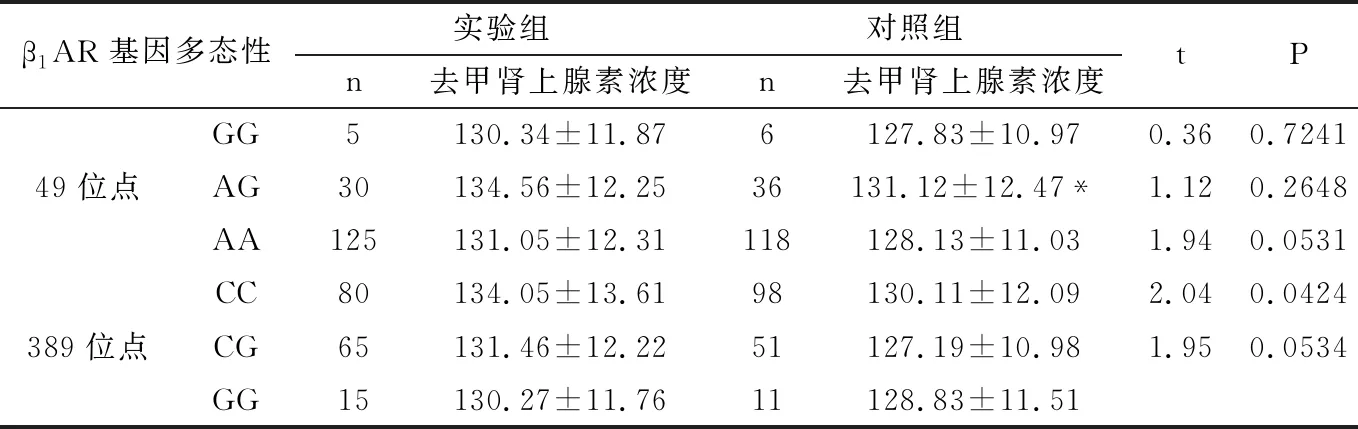
表6 血液去甲腎上腺素濃度與β1AR基因多態性的關系
*P<0.01/0.05
2.6.3 血液多巴胺濃度與β1AR基因多態性的關系 血液多巴胺濃度與β1AR基因多態性的關系見表6所示,實驗組的血液多巴胺濃度與β1AR基因389位點的CC明顯高于對照組,差異均具有顯著統計學意義(P<0.01),而兩組β1AR基因49位點和β1AR基因389位點的CG、GG基因型比較,差異無統計學意義(P>0.05)。
兩組中,49位點的AG基因型的血液多巴胺濃度明顯高于AA型,差異均具有顯著統計學意義(P<0.01),但與GG基因型無差異,差異無統計學意義(P>0.05)。而389位點的基因型的血液去甲腎上腺素濃度無差異,差異無統計學意義(P>0.05)。
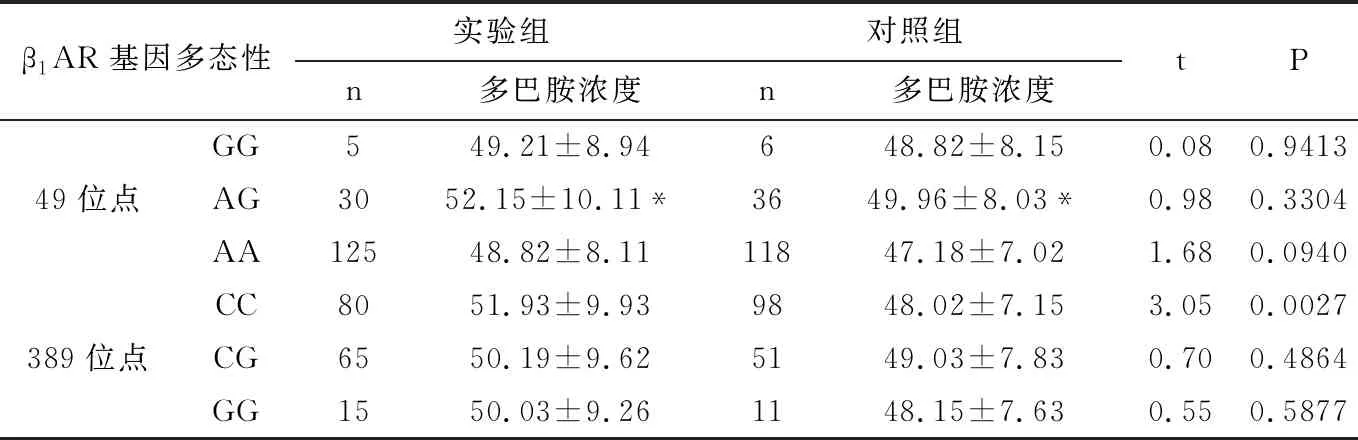
表7 血液多巴胺濃度與β1AR基因多態性的關系
*P<0.01/0.05
2.7 有氧代謝能力指標與β1AR基因多態性的關系
有氧代謝能力指標與β1AR基因多態性的關系見表8-9所示,其中VO2max/kg、O2Plusemax、VCO2max、RQmax、MVV、RFmax、Tmax、Wmax-T、運動后即刻和運動后3min心率等指標中實驗組的β1AR基因389位點的CC明顯高于對照組,并且實驗組的CC亦高于CG和GG組,差異均具有顯著統計學意義(P<0.05/0.01),而兩組β1AR基因49位點和β1AR基因389位點的CG、GG基因型比較,差異無統計學意義(P>0.05)。而恢復后心率所有實驗組均低于對照組,差異均具有顯著統計學意義(P<0.05)。
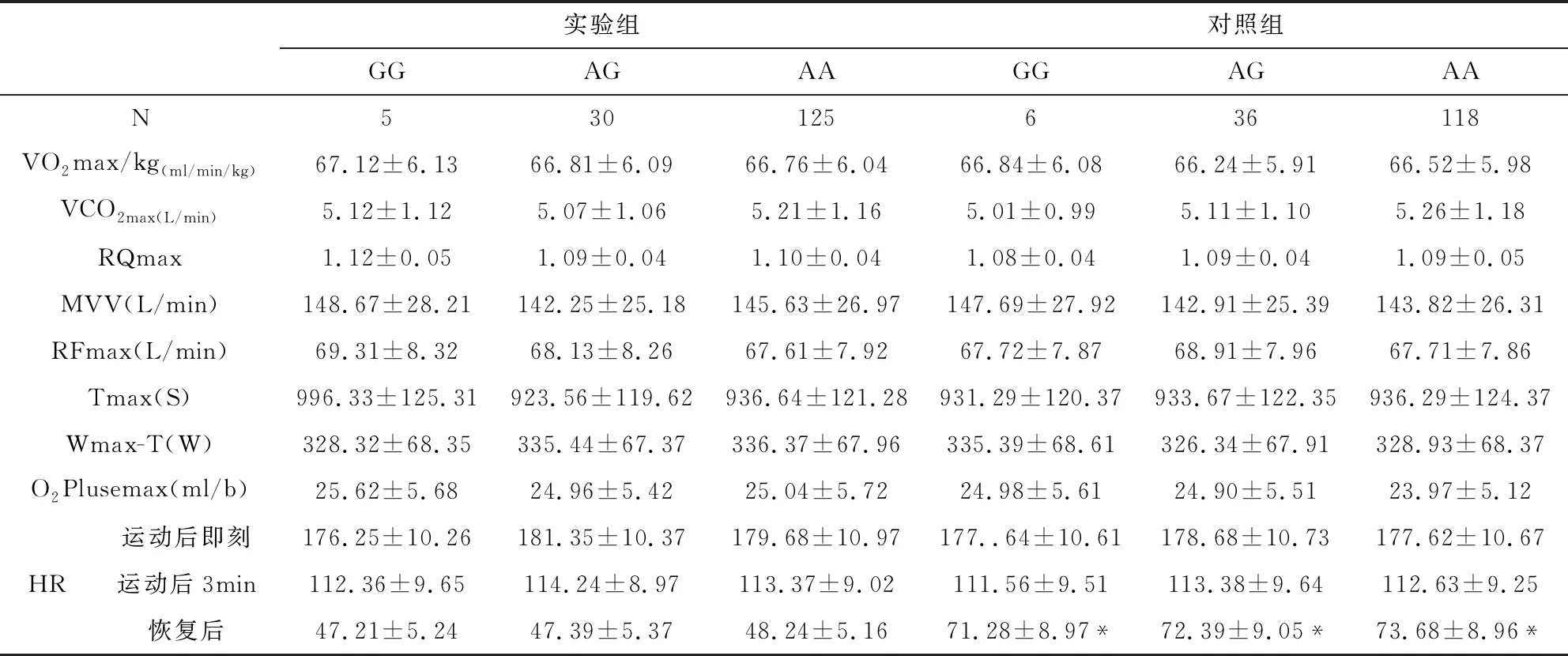
表8 有氧代謝能力指標與β1AR49位點基因多態性的關系
#與同組同位點CC相比P<0.05,*與實驗組同位點相比P<0.01
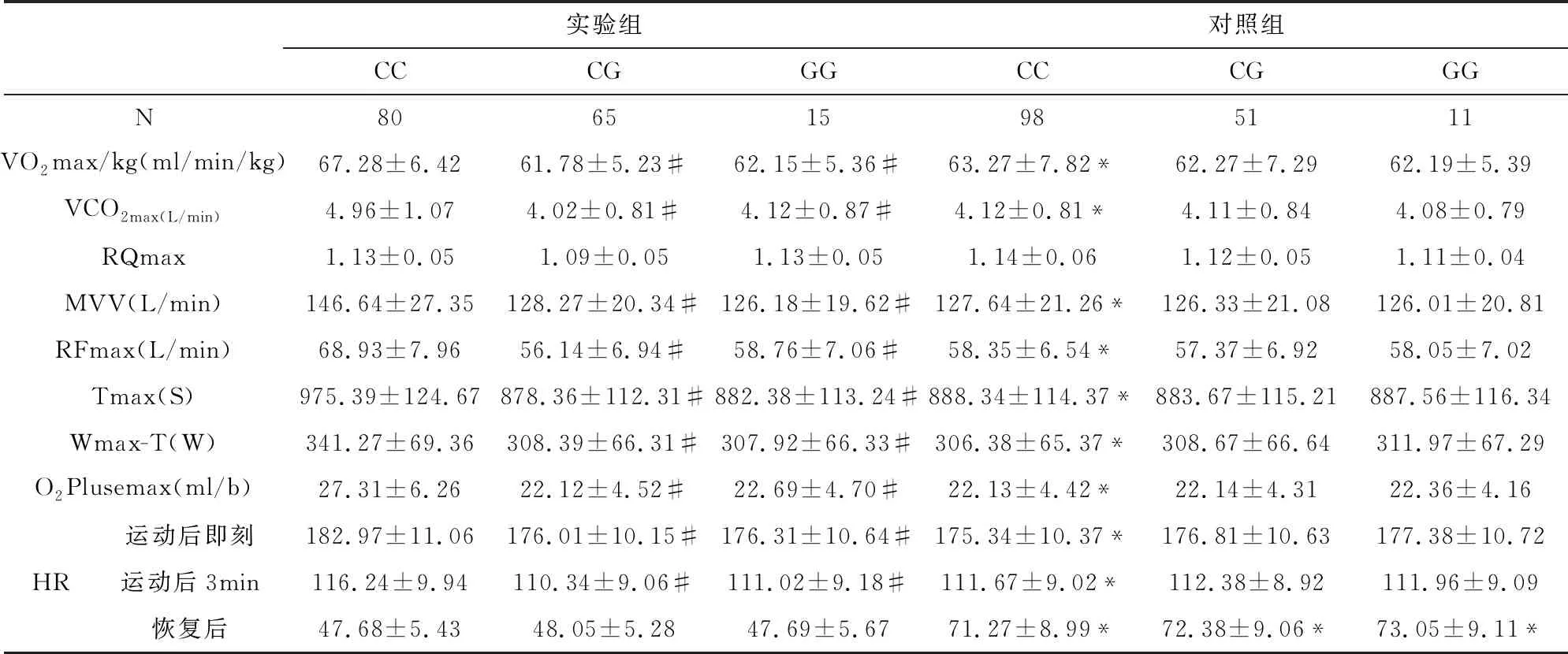
表9 有氧代謝能力指標與β1AR389位點基因多態性的關系
#與同組同位點CC相比P<0.05,*與實驗組同位點相比P<0.01
3 討 論
在心臟功能調節中,交感神經-腎上腺素能系統具有重要作用。應激狀態時,交感神經興奮,交感神經節前纖維支配的腎上腺髓質分泌大量的腎上腺素和去甲腎上腺素,進入血液,與βAR結合。而βAR有四個亞型,包括β1-AR、β2-AR、β3-AR和β4-AR,其中β1-AR主要分布在心臟,由10號染色體短臂q24-26基因編碼,其屬于G蛋白偶聯受體超家族,分為β1-AR、β2-AR和β3-AR三種亞型,而G蛋白系單體蛋白,N端在細胞膜外,C端在胞膜內,其肽鏈跨膜七次,形成膜內外的環狀結構,與配體腎上腺素及去甲腎上腺素結合后,別構激活G蛋白,基礎狀態下,G蛋白由結合GDP的α亞基和β、γ三個亞基構成,當兒茶酚胺類物質與β1-AR結合后,則G蛋白的α亞基上原來結合GDP的位置改為與GTP結合,這時α與βγ分離,游離的結合GTP的α亞基激活腺苷酸環化酶,使ATP形成cAMP,后者再激活蛋白激酶A(PKA),PKA能夠使L型Ca2+通道磷酸化,促進Ca2+內流,從而使心率加快、血壓上升和心肌收縮增強[8,9]。在本實驗中,運動后,三種兒茶酚胺濃度實驗組均高于對照組(P<0.05),證明了此點。
β1-AR基因的rs1801252即第145位的堿基A被G替換,表現在蛋白質的49位絲氨酸(Ser)被甘氨酸(Gly)所代替[1,9],經Eco O109I酶切有GG、AG和AA型三種基因型,與劉建華[1]、石力[9]和Mason DA[10]等的研究一致,而兩組的基因型分布和等位基因頻率的比較,無差異(P>0.05),提示β1AR基因EcoO109I多態性表達與運動能力無關聯。
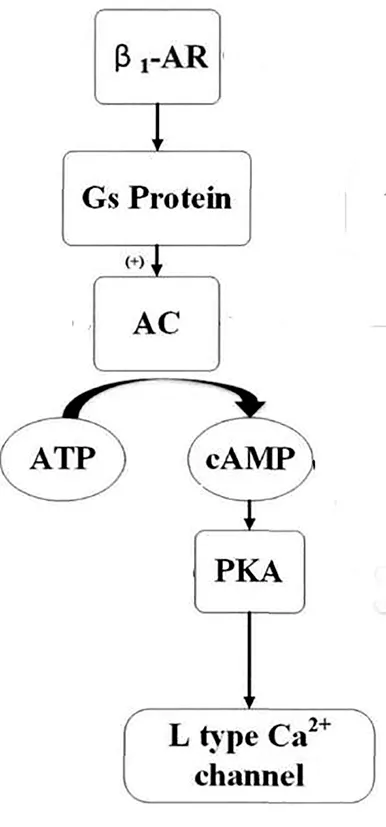
圖6 β1-AR作用機理圖
β1AR1165位C堿基突變為G堿基,引起蛋白質的389位精氨酸(Arg)變成了甘氨酸(Gly)突變,經BcgI酶切有GG、CG和CC型三種基因型,亦與前者[1,9,10]研究一致,但實驗組基因型CC和等位基因G、C基因型頻率與對照組相比,差異顯著(P<0.05),而兩組CG、GG基因型比較無差異(P>0.05),這與Mason DA[10]等人研究結果相仿,說明β1AR基因389位點位于細胞表面,特別是CC基因型和等位基因的C,細胞內羧基末端附近,鄰近磷酸化位點,是G蛋自藕聯及隨后的信號轉導的關鍵點,可能影響了β1-AR的G蛋白偶聯過程,改變了腺苷酸環化酶的活化程度,從而影響受體功能,這種改變極大地影響了心肌細胞的興奮、收縮及傳導功能[11]。但49位點的AG基因型的血液去甲腎上腺素和多巴胺濃度升高顯著,機理尚不清楚,有待進一步增加樣本量進行研究。
有氧能力是運動的基礎能力,出色的有氧能力使運動員獲得較多的營養物質,增加肌肉組織獲取ATP,減少對肌組織氧化酶的抑制作用,同時,增加運動性疲勞產生的5-羥色胺的排泄,減少對肌組織的抑制,促進神經性疲勞的恢復;同時,出色的有氧能力,能促進運動后堆積在神經-肌接頭處的乙酰膽堿的排泄;最后,由于運動造成的鈣通道控制能力降低,高氧促進鈣通道控制能力恢復,使運動產生的肌細胞膜自由基減少,使肌肉恢復正常的興奮性。這樣,運動員出色的有氧能力利于運動員更好地消除在訓練或比賽中積累的疲勞,使機體能夠承受更大的負荷,提高訓練效果[8-13]。
根據璩勝[14]的研究,VO2max/kg反映機體的有氧代謝和供能水平,而MVV反應機體攝氧利用能力;最大氧脈搏(O2plusemax)是VO2/HR的最大值,是反映心臟每搏攝取的最大氧量,是反應心肺功能的綜合指標,其值越高,運動員心肺功能越強,血氧飽和度就高,其是評估心臟輸氧效率高低的最佳指標;在相同遞增負荷的運動模型中,總運動時間(Tmax)在最大負荷階段的運動時間長短的不同,展示了機體運動能力的大小和總運動量,而最大負荷的工作時間(Wmax-T)反應機體承受負荷的能力。本研究顯示:實驗組CC獲氧量明顯高于對照組,并且實驗組的CC亦高于CG和GG組,說明389位點的CC具有較強的獲氧能力,可能與CC基因型鄰近磷酸化位點,增強了腺苷酸環化酶的活化,從而促進了心肌的興奮、收縮及傳導功能,增強了獲氧量,從而增加肌組織獲取ATP,增加運動性疲勞物質5-羥色胺排泄以及神經肌接頭乙酰膽堿排泄[11]。
4 結 論
運動后,運動員的血液兒茶酚胺濃度升高顯著;具有β1AR的389位點C等位基因者,運動能力較好,具有G等位基因者,運動能力相對較差。
本實驗僅研究兒茶酚胺濃度和β1AR多態性與運動能力的相關性,但對心室肌、血管的影響是下一步研究的課題。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
