羽衣甘藍β-胡蘿卜素羥化酶基因的克隆及表達分析
2019-01-24 10:09:26王玉書周明慧
浙江農業學報 2019年1期
關鍵詞:分析
王玉書,王 歡,郭 宇,周明慧,陳 璐,陳 陽
(1.齊齊哈爾大學 生命科學與農林學院,黑龍江省抗性基因工程與寒地生物多樣性保護重點實驗室,黑龍江 齊齊哈爾 161006; 2.齊齊哈爾大學 化學與化學工程學院,黑龍江 齊齊哈爾 161006)
類胡蘿卜素是在自然界植物、細菌和真菌中廣泛存在的一類色素類物質,同時在植物光合作用中起重要作用[1-2]。β-胡蘿卜素羥化酶在植物類胡蘿卜素生物合成代謝途徑中具有非常重要的作用,其作用是催化β-胡蘿卜素通過中間代謝物β-隱黃質生成玉米黃質,因其增加了植物細胞中玉米黃質的含量,所以該反應在植物葉黃素循環中有非常重要的意義[3]。有研究表明,β-胡蘿卜素羥化酶基因的過量表達有助于細胞中玉米黃質的過量積累,進而提高細胞中類胡蘿卜素含量[4];而利用反義抑制和T-DNA突變獲得抑制β-羥化酶基因表達的擬南芥植株,由于β-羥化酶基因的缺陷,它們合成隱黃素等類胡蘿卜素的能力減弱,抗逆性隨之大幅下降[5-6]。目前已從擬南芥[7]、煙草[8]、柑橘[9]、辣椒[10]等植物中分離鑒定了許多β-胡蘿卜素羥化酶基因。通過生物信息學比對分析發現,這些基因均具有非常保守的同源序列。
羽衣甘藍葉片顏色鮮艷,較耐嚴寒、干旱,因此成為冬春季節中重要的綠化景觀植物。研究表明,在羽衣甘藍生長過程中,葉色與葉綠素、類胡蘿卜素及花青素的含量密切相關[11]。然而到目前為止,關于羽衣甘藍中類胡蘿卜素生物合成途徑的相關基因研究還鮮有報道。本研究采用同源克隆和RT-PCR技術獲得羽衣甘藍β-胡蘿卜素羥化酶基因BoBCH,并對其進行了生物信息學分析,研究該基因在不同組織、不同發育階段的表達情況,為羽衣甘藍β-胡蘿卜素羥化酶基因的功能鑒定及調節機制提供理論基礎。
1 材料與方法
1.1 材料
紫葉羽衣甘藍DH系D07于2017年1月種植于齊齊哈爾大學園藝試驗基地,分別取D07的根、莖、葉不同組織和幼苗期、蓮座期、觀賞期3個時期的幼嫩葉片,經液氮速凍后-80 ℃保存,備用。
1.2 方法
1.2.1 總RNA的提取及cDNA的合成
利用Trizol(Invitrogen,USA)試劑盒提取羽衣甘藍總RNA,之后經NanoDrop 2000分光光度計(Thermo Scientific,USA)測定濃度及D260/D280,瓊脂糖凝膠電泳檢測RNA完整性。檢測合格后,取經DNase處理后RNA作為模板,使用反轉錄試劑盒將2 μg植物總RNA反轉錄為cDNA,檢測待用。
1.2.2 引物設計與基因克隆
基于作者前期全基因組測序篩選得到的BoBCH基因序列,設計克隆引物F(5′-ATGGCGGCAGCACTCTCATCAATCTC-3′)和R(5′-AGAGGTGGAAACCTTGTTGTATAATTTGTAA-3′)。cDNA擴增使用ABI Veriti PCR 擴增儀進行。PCR體系:2×Ultra-Pfu Master Mix酶(Dye Plus)5 μL、模板1 μL、正反引物各0.8 μL和ddH2O 2.4 μL。PCR反應條件為:94 ℃ 2 min;94 ℃ 30 s,53 ℃ 30 s,72 ℃ 1 min,30個循環;72 ℃ 8 min。PCR產物電泳分析后,回收、連接、轉化及測序,得到BoBCH的cDNA序列信息。
1.2.3 生物信息學分析
利用NCBI網站對BoBCH的cDNA序列在線查找開放閱讀框(ORF);利用蛋白質分析在線工具(http://web.expasy.org/protparam/)在線程序分析編碼蛋白的理化性質(氨基酸組成、相對分子質量、等電點等);利用ExPasy-ProtScale預測氨基酸的親/疏水性;利用NCBI在線工具CDD(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析氨基酸序列的保守結構域;利用SignalP4.1和Wolf-Psort進行信號肽和蛋白亞細胞定位預測;TMHMM2.0對蛋白進行跨膜區分析;用MEGA6.0軟件采用鄰接法進行氨基酸序列比對以及構建系統發育樹。
1.2.4 基因表達分析
根據測序結果,設計熒光定量引物分別為BoBCH-F和BoBCH-R,以18S rRNA作為內參,引物為18S rRNA-F和18S rRNA-R(表1)。引物采用Roche LCPDS2軟件設計并由北京擎科新業生物技術有限公司合成。qRT-PCR反應根據QuantiFast? SYBR? Green PCR Kit試劑盒(Qiagen,Germany)說明書,在Light Cycler?480Ⅱ型熒光定量PCR儀(Roche,Swiss)上進行反應。每個反應采用3個生物學重復和3個技術重復。反應結束后,應用2-△△Ct算法進行相對定量計算[12]。
表1PCR引物
Table1The primers of PCR used in this study
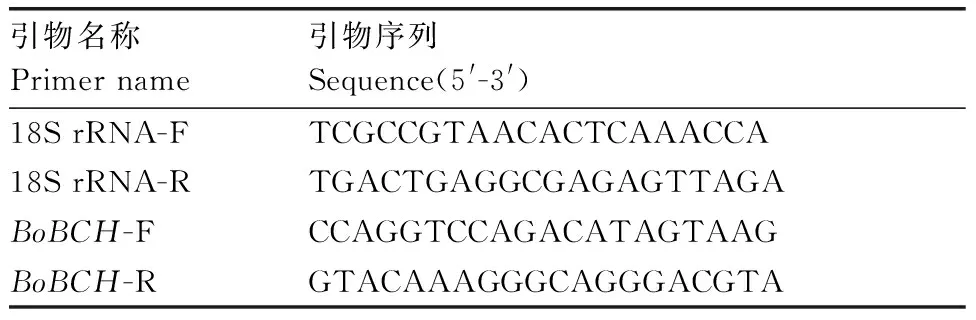
引物名稱Primer name引物序列Sequence(5'-3')18S rRNA-FTCGCCGTAACACTCAAACCA18S rRNA-RTGACTGAGGCGAGAGTTAGABoBCH-FCCAGGTCCAGACATAGTAAGBoBCH-RGTACAAAGGGCAGGGACGTA
2 結果與分析
2.1 BoBCH基因的克隆
以羽衣甘藍D07觀賞期葉片cDNA為模板,通過PCR擴增后獲得BoBCH基因片段,經過瓊脂糖凝膠電泳檢測獲得條帶與預期大小相符(圖1)。對擴增結果進行測序分析,將該基因命名為BoBCH(GenBank登錄號為MH016242)。
2.2 BoBCH基因cDNA序列的生物信息學分析
2.2.1BoBCH基因編碼氨基酸序列分析
利用ORF finder在線工具翻譯BoBCH的cDNA序列,BoBCH基因編碼301個氨基酸。利用蛋白質分析在線工具(http://web.expasy.org/protparam/)分析BoBCH蛋白的物理化學性質,結果表明BoBCH蛋白由20種氨基酸組成,其中絲氨酸(Ser)含量最多,半胱氨酸(Cys)含量最少,帶負電氨基酸殘基25個,帶正電氨基酸殘基37個;分子量約為33.8 ku,理論等電點為9.67;親水性平均系數(GRAVY)為-0.072,說明BoBCH蛋白為親水蛋白。利用ExPasy-ProtScale預測氨基酸的親/疏水性表明:該蛋白的最高值為2.133,在第140個氨基酸處,疏水性最強;最低值為-3.044,在第170個氨基酸處,親水性最強。同時,從整條多肽鏈的氨基酸預測值來看,親水性氨基酸殘基所占比例高于疏水氨基酸殘基,由此推斷,BoBCH蛋白是一種親水性蛋白,與基因編碼氨基酸序列的分析結果一致。利用NCBI在線工具CDD分析BoBCH氨基酸保守蛋白結構域,結果表明,BoBCH屬于FA_hydroxylase蛋白超家族(圖2)。

M, DNA marker; 1~3, RT-PCR擴增產物。M, DNA marker; 1-3, RT-PCR products.圖1 羽衣甘藍BoBCH基因PCR擴增結果Fig.1 PCR amplification of BoBCH gene from the kale
2.2.2 BoBCH蛋白的信號肽、跨膜結構及亞細胞定位預測
運用在線工具Signal P4.1 server進行信號肽分析預測,結果顯示,C值、S值、Y值均小于閾值0.45,推測BoBCH蛋白無信號肽結構,屬于非分泌蛋白。蛋白跨膜結構預測分析顯示, BoBCH蛋白在93~115、130~152、183~200和204~226處有4個跨膜結構域(圖3)。亞細胞定位分析結果顯示,BoBCH可能定位于葉綠體中。
2.2.3 BoBCH蛋白序列的進化分析
為了預測BoBCH基因的功能,從NCBI中查找到其他植物的BCH同源蛋白序列,采用MEGA6.0鄰接法與羽衣甘藍BCH蛋白序列對比并構建無根系統進化樹(圖4)。羽衣甘藍與結球甘藍、甘藍型油菜、擬南芥等10個高等植物的BCH序列比對顯示,這11個植物BCH氨基酸序列被聚為兩大類。羽衣甘藍BCH與結球甘藍處于同一分支,其親緣關系最近,其序列一致性高達99%;與醉蝶花、蘿卜、大白菜等物種的BCH蛋白在進化上親緣關系較遠。
2.3 BoBCH基因特異性表達分析

圖2 BoBCH的保守結構域預測Fig.2 Prediction of BoBCH conserved domains

圖3 BoBCH蛋白跨膜結構域預測Fig.3 Prediction of transmembrane domains in BoBCH protein
為了研究BoBCH基因在羽衣甘藍不同組織中的表達情況,對不同組織提取的RNA進行了實時熒光定量qRT-PCR分析。結果表明,觀賞期BoBCH在不同器官中均有表達,但表達具有組織特異性,其在葉片中表達量最高,極顯著高于其他器官(P<0.01);其次是莖;而在根中表達量最少,與其他組織表達差異極顯著(P<0.01)。BoBCH基因在D07幼苗期、蓮座期和觀賞期3個時期葉片中的表達量也存在顯著差異,其中觀賞期時表達量最高,蓮座期和幼苗期表達量差異不顯著(P>0.05),觀賞期BoBCH的表達量約為其他兩個時期表達量的10倍(圖5)。

圖4 羽衣甘藍BoBCH與其他植物BCH蛋白的系統進化樹Fig.4 Phylogenetic tree analysis of BoBCH protein in kale and BCHs of other plants
3 討論
本研究利用同源克隆技術從羽衣甘藍D07中克隆得到BoBCH基因的全長cDNA序列。通過序列分析發現,BoBCH與結球甘藍、甘藍型油菜、擬南芥、琴葉擬南芥等植物的BCH氨基酸序列相似性達76%以上,它們均屬于FA_hydroxylase蛋白超家族,表明該結構域在分析進化過程中穩定性較好,在類胡蘿卜合成及抵抗非生物脅迫中發揮類似的作用[13-14]。氨基酸序列同源性分析表明,BoBCH蛋白與結球甘藍蛋白在同一分支,進化關系最近,因此推測它們應該具有相類似的功能,可能與類胡蘿卜素合成有關。跨膜結構預測和亞細胞定位預測得知,BoBCH蛋白含有脂肪酸羥化酶超家族保守域(BoBCH保守區域分別為93~115、130~152、183~200和204~226處殘基),可能定位于葉綠體中。這與目前己報道的其他物種中β-胡蘿卜素羥化酶定位于葉綠體類囊體膜上的結論一致[15],推測該類酶經由葉綠體導肽從細胞質運輸到葉綠體類囊體膜上。這些生物信息分析對于將來研究BoBCH蛋白相關功能具有指導意義。

不同大寫字母代表不同組織或發育時期間表達量差異顯著(P<0.01)。The bars with different capital letters showed the significance(P<0.01).圖5 BoBCH在D07不同組織和不同發育時期中的相對表達量Fig.5 Relative expression levels of BoBCH in different tissues and different developmental stages of D07
研究發現,羽衣甘藍β-胡蘿卜素羥化酶基因BoBCH在根、莖、葉中均能表達,但不同組織器官中存在顯著性差異,BoBCH在葉片中的表達量最高,其次是莖,根中表達量最低。在不同發育階段的結果表明,在幼苗期與蓮座期時,BoBCH基因表達量均較低,并且無顯著差異,然而觀賞期時BoBCH的表達量高達幼苗期與蓮座期的10倍之多,表明在觀賞期時葉片中BoBCH基因積累較多,并發揮其催化作用。這可能是因為BoBCH的活性與葉片中類胡蘿卜素含量密切相關[16],說明本研究克隆得到的BoBCH基因可能在羽衣甘藍葉片的類胡蘿卜素合成途徑中發揮重要作用。
本研究克隆獲得羽衣甘藍BoBCH基因的cDNA序列,并對該基因在不同組織及不同發育時期的表達模式進行了分析,推測BoBCH基因在羽衣甘藍類胡蘿卜素代謝調控中起重要作用。后續研究中,將會就該基因的瞬時表達、轉基因驗證等方面進行BoBCH基因功能的研究,并對該基因與類胡蘿卜素生物合成途徑中相關酶基因之間的互作關系進行深入研究,進而全面解析羽衣甘藍類胡蘿卜素合成代謝調控的分子機制。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
