肉牛及奶牛生長階段飼糧的氨基酸限制性順序及理想氨基酸模式研究現狀
2019-01-03 02:16:42刁其玉
飼料工業 2018年21期
■李 媛 刁其玉 屠 焰
(中國農業科學院飼料研究所奶牛營養學北京市重點實驗室,北京100081)
NRC(2012)[1]和NRC(1994)[2]已給出詳細的各個生長階段的豬和家禽的氨基酸營養需要量,NRC(2000)[3]也給出了生長肉牛的氨基酸需要量預測模型,而關于奶牛生長階段的氨基酸營養需要還缺乏系統的研究。NRC(2001)[4]指出后備牛的蛋白和氨基酸需要量的確定可劃分為四個階段:斷奶前、過渡期(2~4月齡)、4月齡~配種期、配種后~產犢前。研究后備母牛的氨基酸需要量有助于提升牛場的經濟效益和母牛的生產力。但目前氨基酸限制性順序及需要量的研究多集中在犢牛和成年牛,有關于生長階段后備牛氨基酸需要量的研究鮮見報道,飼糧氨基酸需要量及其理想氨基酸模式還處于空白。本文對牛生長階段的氨基酸營養研究現狀做一綜述,旨在為后備牛生長階段的氨基酸限制性順序及需要量模型的研究提供參考。
1 牛生長階段的限制性氨基酸
1.1 牛生長階段的主要限制性氨基酸
小腸是生長牛吸收氨基酸的主要場所。小腸中的氨基酸主要來源于飼糧中的瘤胃非降解蛋白和瘤胃微生物蛋白[5],所以飼糧氨基酸組成并不是決定牛生長階段的氨基酸需要量和限制性順序的唯一因素。當微生物蛋白的量不能滿足牛的生長需要時,某些氨基酸也會成為牛的限制性氨基酸。Titgemeyer等(1990)[6]研究表明,當真胃灌注蛋氨酸(Met)時,生長牛的氮(N)沉積增加17%。此外,只有當灌注Met時,N沉積才增加,所以Met可能是生長公牛的第一限制性氨基酸。但是僅僅灌注Met的效果不及混合灌注氨基酸的效果顯著。Greenwood等(1999)[7]和L?est等(2001)[8]研究表明,Met、組氨酸(His)、賴氨酸(Lys)、亮氨酸(Leu)和纈氨酸(Val)是生長荷斯坦公牛的限制性氨基酸。這些結果顯示,生長牛的限制性氨基酸不止一種,相比于只有一兩種關鍵氨基酸,到達真胃的氨基酸越豐富,牛生長性能越好。
1.2 牛生長階段的氨基酸限制性順序
目前對犢牛及生長公牛氨基酸限制性順序的研究很多。表1匯總了近年來國內外關于牛生長階段氨基酸限制性順序的研究結果。結果表明,生長牛的氨基酸限制性順序受品種、日齡、體重等多個因素影響。Lys和Met為牛生長階段的前兩種限制性氨基酸,蘇氨酸(Thr)、色氨酸(Trp)和His可能是牛生長階段的第三限制性氨基酸。
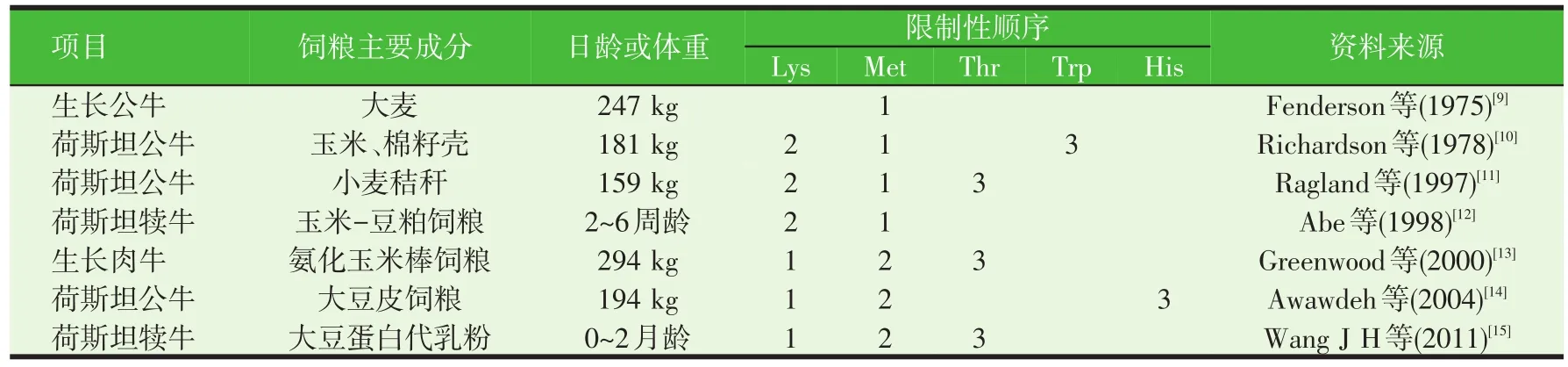
表1 牛生長階段的氨基酸限制性順序
大量研究表明飼糧的過瘤胃蛋白的含量是影響反芻動物生長階段氨基酸限制性順序的一個重要因素。大麥易被瘤胃降解,過瘤胃低,Fenderson等(1975)[9]研究表明,當生長公牛的飼糧含80%的大麥時,Met是第一限制性氨基酸。玉米蛋白的過瘤胃率較高,以玉米為基礎飼糧的生長公牛灌注Lys,發現Lys可能是第一限制性氨基酸。當飼糧蛋白組成多為瘤胃降解蛋白時,到達小腸的氨基酸多為微生物蛋白,此時限制性氨基酸多與微生物蛋白的組成有關。Nimrich等(1970)[16]等研究表明以尿素為唯一氮源的生長綿羊,氨基酸限制性順序為Lys、Met、Thr。
2 牛生長階段的氨基酸平衡模式
2.1 氨基酸平衡
氨基酸平衡是保證氨基酸利用效率最大化的關鍵。實際生產中都會嚴格把控氨基酸的添加量,考慮多種限制性氨基酸之間的平衡關系,確定適宜的氨基酸添加比例,做到精準飼喂,以確保氨基酸平衡。飼糧中某些氨基酸的不足和過量都會使氨基酸之間失去平衡,從而影響畜禽的生長發育。Abe等(1999)[17]研究發現,向體重約為103 kg生長牛真胃內投喂0.333 g/kg BW的Met,育成牛會出現明顯的中毒癥狀;Abe等(2001)[18]研究發現,150 kg的育成牛每天攝入64 g Lys時腹瀉率明顯增加。此外飼糧氨基酸平衡是降低飼養成本,節約蛋白飼料資源的一個首要手段。氨基酸平衡能保證最大程度發揮各個氨基酸的利用效率,從而降低飼糧蛋白水平,節約成本。
2.2 牛生長階段的理想氨基酸模式(IAAP)的研究進展
近年來,理想氨基酸模式在成年奶牛的研究較多,對生長階段反芻動物的氨基酸平衡模式的研究較少。Hill等(2008)[19]研究表明,5周齡前犢牛代乳料中Lys、Met和 Thr的適宜比例為 100∶31∶60;刁其玉[20]提出羔羊的氨基酸平衡模型為Lys、Thr和Trp比值為100∶50.5∶14.3。云強等(2011)[21]研究表明,8~16周齡犢牛的玉米-豆粕型開食料中適宜的Lys、Met比例為3.1∶1。
氨基酸平衡模式的構建除與飼糧和動物本身有關外,還受研究方法的影響。動物理想氨基酸模式的研究方法主要有析因法、計量效應法、屠體法和氨基酸部分扣除法。許多研究指出析因法和劑量效應法易受其他因素的影響,結果都存在一定的問題。陳正玲(2001)[22]研究用三種方法確定的肉鴨IAAP之間存在一定差異,屠體法測定的模式只能作為參考,劑量效應法測定的維持生長模式與AA部分扣除法確定的模式相對較為接近,AA部分扣除法確定的模式可能與肉鴨實際的IAAP更為接近。AA部分扣除法是基于析因法和劑量效應法兩者優點于一體的準確測定IAAP的有效方法。通過扣除飼糧部分氨基酸觀察動物的生長性能、飼料轉化率、N沉積等指標,從而確定動物的氨基酸限制性順序。Wang等(2010)[23]采用AA部分扣除法確定了豬的理想氨基酸模型,并作為NRC(2012)[1]修訂豬IAAP的一個重要依據。但氨基酸扣除法在反芻動物IAAP的研究中應用不多,有兩篇研究采用AA部分扣除法確定了犢牛和羔羊的IAAP。Wang J H等[15]采用氨基酸扣除法指出獲得最大ADG時0~2和4~6周齡犢牛的Lys、Met和Thr需求比例為100∶35∶63和100∶27∶67。李雪玲等[24]采用氨基酸扣除法確定了60~90日齡和90~120日齡羔羊獲得最大ADG、F/G和DP時,Lys、Met、Thr和Trp的適宜比例為100∶44∶42∶8和100∶41∶38∶11,并且采用屠體法驗證發現二種方法確立的羔羊氨基酸適宜比例相近。
目前氨基酸部分扣除法已應用于單胃動物的氨基酸限制性順序及平衡模式的研究,但此方法在反芻動物尤其在生長牛IAAP的研究方面的應用較少。由于反芻動物的特殊消化生理,該方法是否適用于不同階段生長牛的IAAP研究需進一步驗證,生長牛適宜的理想氨基酸模式研究方法也需進一步探索。
3 牛生長階段氨基酸需要量的研究
由于反芻動物的特殊消化過程,評定反芻動物的氨基酸需要量不能根據其攝入的氨基酸量進行評定,要根據其進入小腸并被吸收的氨基酸量,也就是代謝氨基酸量來確定。表2和表3匯總了近年來國外學者對生長階段牛的代謝氨基酸需要量的研究結果。我們可以看出品種、體重、日增重均是影響生長階段牛的氨基酸需要量的因素。
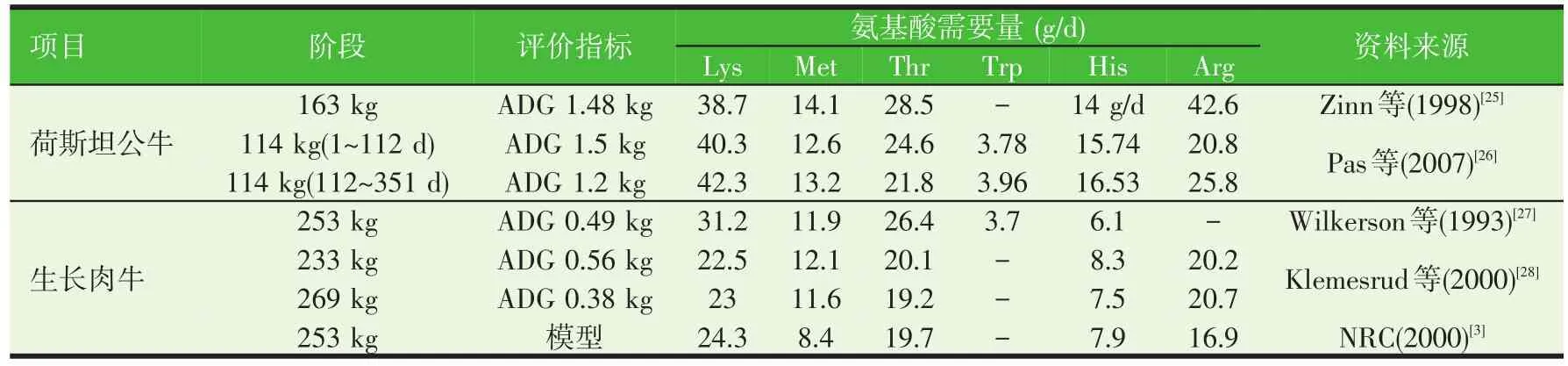
表2 牛生長階段代謝氨基酸的需要量
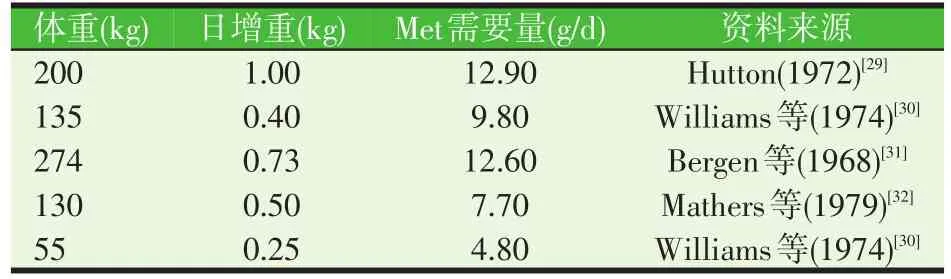
表3 生長肉牛Met的需要量
R.A.Zinn(1988)[33]根據表3中的研究成果擬合出了生長肉牛Met的需要量模型METR=1.956+0.029 2PG+0.029 0W(PG:蛋白增重,W:體重,R2=0.92),用此模型計算得的氨基酸需要量與其動物試驗測定的生長肉牛氨基酸需要量相近。R.A.Zinn等(1998)[25]等將上述公式具體化,得出了依據體重和日增重確定Met需要量的模型:METR=(1.956+0.029 2×ADG[268-(29.4×0.055 7BW0.75×ADG1.097)÷ADG]+0.112×BW0.75),經過100頭生長公牛的飼養試驗驗證得出本公式計算的小腸Met的提供量與實際測定量之間的相關性達0.99,此模型可用于計算肉牛生長階段Met需要量。Monta?o等(2016)[34]用192頭公牛驗證了NRC(2000)[3]氨基酸需要量計算模型的準確性,認為NRC(2000)[3]提供了可靠的計算飼糧提供給小腸代謝氨基酸量及生長牛代謝氨基酸的需要量的模型。但目前很少有研究證實此計算模型在生長牛增重關鍵時期的實用性。
由于瘤胃發酵的轉化導致進入生長牛小腸的氨基酸的數量和種類不統一,計算到達小腸的氨基酸量非常困難,所以近些年有關建立不同階段生長牛的氨基酸需要量模型研究很少。但模型的建立是確定各階段反芻動物氨基酸需要量的基礎,通過模型的擬合確立各階段生長牛的氨基酸需要量是有待研究的重點。模型建立需要以大批量數據為基礎,但目前生長階段后備牛的氨基酸需要量還處于空白,所以研究生長后備牛的氨基酸需要量,為后續模型的建立提供基礎刻不容緩。
4 牛生長階段的微生物蛋白(MCP)的氨基酸組成及影響因素研究
瘤胃微生物以發酵產生的ATP和揮發性脂肪酸為能量和碳源,以氨基酸和氨為底物合成微生物蛋白質。微生物蛋白質是一種“高質量”的蛋白質,它會同過瘤胃蛋白質隨食糜進人真胃和小腸,供動物體吸收和利用。大量研究表明,MCP可提供成年奶牛所需的絕大部分氨基酸;犢牛由于瘤胃發育不完善,MCP合成有限,微生物提供的氨基酸較少,其氨基酸主要由飼糧提供;生長牛的瘤胃發育基本完善,已具備基本的合成MCP的能力,所以MCP為生長牛氨基酸的另一主要來源。因此,MCP的氨基酸組成也可能會影響生長牛的氨基酸平衡及需要量。
大量研究證明,MCP的氨基酸組成與微生物的種類直接相關。瘤胃微生物主要分為細菌、原蟲和微生物三種,此外還可根據其存在方式分為液相菌(游離于瘤胃液)和固相菌(附著于飼料顆粒)。表4匯總了近幾年研究中的瘤胃微生物蛋白的主要氨基酸組成。從表中可以看出不同群系MCP主要氨基酸組成存在一定差異,Rodríguez等[35]和周勃等[36]結果發現固相菌中Lys含量高于液相菌;王洪榮等(2009)[37]研究發現,原蟲、細菌和微生物的氨基酸組成存在一定的差異,原蟲的Val含量高于細菌和微生物,細菌和微生物的Lys含量高于原蟲。
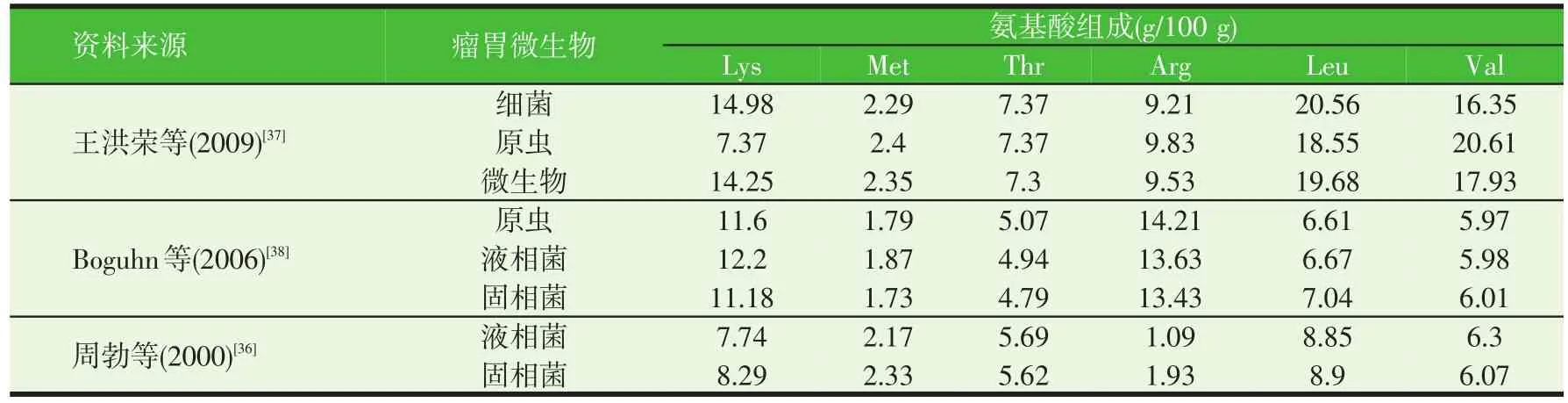
表4 瘤胃MCP的氨基酸組成
有研究表明,微生物區系的變化是造成瘤胃微生物中某些氨基酸含量和比例差異較大的原因。而瘤胃微生物區系可能受飼糧等其他因素的影響。Vogel等(1983)[39]研究表明,飼糧對瘤胃微生物的氨基酸組成沒有影響;而Erasmus等(1994)[40]、王洪榮(1998)[41]的研究表明瘤胃微生物的氨基酸組成會受飼糧的影響。迄今,有關MCP的氨基酸的研究已有很多,但結果不一,瘤胃微生物氨基酸組成是否會改變,改變的原因及機理仍不清楚。作為牛生長階段氨基酸主要來源之一,MCP的氨基酸組成對其氨基酸平衡及需要量的影響未見研究。所以以MCP氨基酸組成研究為切入點,研究其對生長牛氨基酸平衡的影響是生長牛氨基酸營養研究的另一個創新點。
5 小結
奶牛理想氨基酸模型的建立已取得一定的進展,但大多局限于成年奶牛和犢牛,后備牛理想氨基酸模型的研究還處于空白。飼糧是決定后備牛氨基酸平衡的一個重要因素,研究飼糧對氨基酸需要量的影響及生長牛小腸氨基酸的吸收及轉運機制對氨基酸平衡模式的構建有重要意義。此外,還需把MCP的氨基酸組成作為重點,加強對MCP氨基酸組成及改變機理的研究。最終以飼糧和MCP氨基酸含量和組成為基礎,篩選合適的理想氨基酸模型的構建方法,成功構建后備牛理想氨基酸模式。目前的氨基酸需要量研究大多以動物血液或肉中的氨基酸量為基礎,反向倒推氨基酸需要量;或者以飼糧和小腸食糜氨基酸含量為基礎確定氨基酸需要量。MCP的氨基酸組成很少被考慮,所以MCP是否對生長牛氨基酸平衡和需要量有影響也是建立生長牛理想氨基酸模型需考慮的另一主要因素。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
