飼料牛磺酸含量對不同生長階段尼羅羅非魚生長性能的影響
2019-01-03 08:40:16何凌云周銘文孫云章葉繼丹
飼料工業 2018年22期
關鍵詞:生長
■何凌云 周銘文 孫云章 宋 凱 葉繼丹
(集美大學水產學院廈門市飼料檢測與安全評價重點實驗室,福建廈門361021)
牛磺酸是一種含磺酸基的氨基酸,是動物機體內含量最多的氨基酸,但它并不參與蛋白質的合成[1]。牛磺酸有很多重要的生物學作用[2-3],如與膽酸合成膽汁酸,使其限速酶膽固醇7α-羧化酶的活性升高,加強膽固醇向膽汁酸轉化的能力[4],促進脂肪的合成與代謝。牛磺酸是多數動物機體的一種條件性必需氨基酸,其自身(貓科和狐貍除外)有一定合成牛磺酸的能力[5],但其合成能力又與動物種類、個體營養狀況、蛋白質進食量和胱氨酸有效性有關[6]。與陸生恒溫動物相比,魚類合成牛磺酸的能力均較低,并且這一能力在肉食性魚類及幼齡魚上表現得尤為明顯,其體內牛磺酸合成量大多不能滿足正常生長所需,需要從外源(食物)供給[5]。飼料中添加牛磺酸能顯著提高斜帶石斑魚(Epinepheluscoioides)[7]、黃尾鰤(Seriola quinqueradiata)[8]和大菱鲆(Scophthalmusmaximus L.)[9]的生長性能,當飼料中牛磺酸不足時,會降低真鯛(Pagrus major)[10]和牙鲆(Paralichthys olivaceus)[11]的生長性能。由于陸生植物牛磺酸含量很低或不含牛磺酸,而魚粉(包括其他動物產品)中牛磺酸含量較高,這在配制生產以植物蛋白源為主要蛋白源的水產飼料時如何合理使用牛磺酸,有針對性地為養殖魚類提供個性化的飼料配制方案,實現養殖魚類的最佳生長等方面有積極意義。近年來,國內外學者研究了草魚(Ctenopharyngodon idellus)[12]、斜帶石斑魚[13]、虹鱒(Oncorhynchus mykiss)[14]、牙鲆[15]、鱸魚(Lateolabrax japonicus)[16]等多種養殖魚類對牛磺酸的需要量。目前關于尼羅羅非魚對飼料中牛磺酸的適當需求量已有研究[17],但尼羅羅非魚在關鍵生長期中的不同階段對牛磺酸需要的影響還未見報道,因而,該試驗目的是為研究飼料牛磺酸含量和不同生長階段對尼羅羅非魚生長性能的影響,為尼羅羅非魚飼料中牛磺酸的使用提供依據。
1 材料與方料
1.1 試驗飼料
試驗用酪蛋白、明膠和牛磺酸(純度為99%)均購自鄭州明瑞化工產品有限公司,維生素預混料和礦物質預混料均購自廈門嘉康飼料有限公司。該試驗是以酪蛋白和明膠(無牛磺酸)作為蛋白源,魚油、大豆油和大豆卵磷脂作為脂肪源,玉米淀粉作為糖源制備成尼羅羅非魚的基礎飼料。然后在飼料中添加比例為0、400、800、1 200 mg/kg的牛磺酸,制備成4種不同牛磺酸含量的尼羅羅非魚幼魚試驗飼料。當飼料中牛磺酸比例增加時,就相應的減少結晶纖維素比例。試驗基礎飼料配方及營養水平見表1。酪蛋白和明膠粉碎過篩(80目)。按飼料配方將各種飼料原料充分混勻,再加一定水攪拌,用雙螺桿制粒機制成直徑2.5 mm、3.5 mm的顆粒料,65℃烘箱烘干,裝自封袋密封,備用。
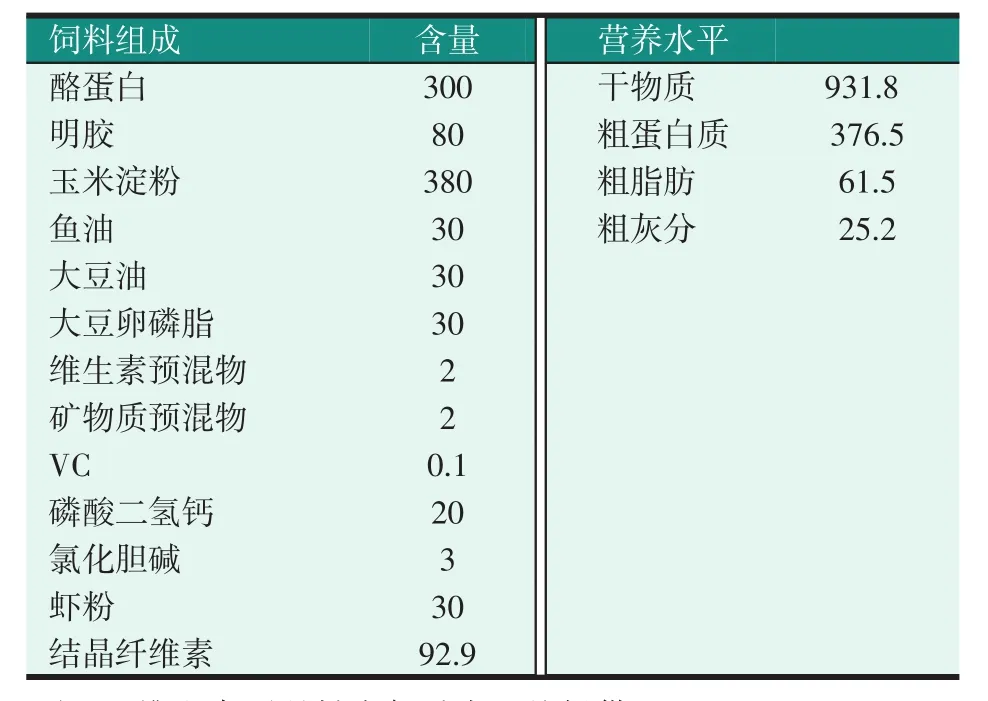
表1 基礎飼料配方及營養水平(風干基礎,g/kg)
1.2 養殖管理
羅非魚幼魚暫養2周后,從中挑選健康魚400尾(體質量約為4.25 g),隨機分放到16個的150 L容量的水族箱中,每個水族箱25尾魚。試驗分為4個飼料處理組,每個飼料組設4個水族箱(重復組)。
養殖試驗期為84 d。養殖過程中,每日投喂兩次(08:30和18:30),每次投喂至表觀飽食。在投喂后0.5 h吸去糞便和殘餌,每次換水量約1/4。將試驗期分為3個生長階段(1~28、29~56、57~84 d),每個階段為28 d。養殖期間水溫為25~31℃,溶解氧含量為5.7~6.6 mg/l,硝酸鹽氮含量0.10~0.32 mg/l。
1.3 樣品采集
試驗在第28、第56 d和第84 d分別對每個水族箱的魚稱總重。再放回到原來的水族箱,24 h后,從每個水族箱中隨機抽取6尾魚,丁香酚麻醉,逐尾魚稱重并測量體長,解剖并取肝臟、腸等內臟,分別稱重。在第84 d時,從每個水族箱中隨機抽取3尾魚用于全體體成分分析。
1.4 樣品測定
測定飼料原料、試驗飼料和魚體樣品中的常規營養成分。水分測定采用105℃烘箱常壓干燥法烘至恒重;粗蛋白質測定采用半微量凱氏定氮法;粗脂肪測定采用索氏抽提法;粗灰分測定采用馬弗爐(550℃)灼燒法。
1.5 數據統計與分析
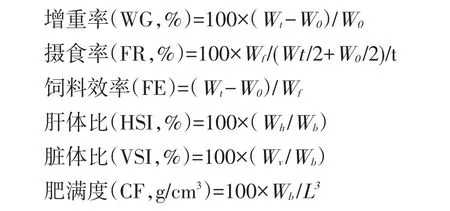
式中:W0——初始均質量(g);
Wt——終末均質量(g);
Wf——攝食的飼料總量(g);
Wb——樣品魚的體質量(g);
Wh——樣品魚肝臟質量(g);
Wv——樣品魚內臟質量(g);
L——樣品魚體長(cm);
t——試驗天數(d)。
試驗數據均使用SPSS17.0進行單因素(One-way ANOVA)和雙因素方差(Two-way ANOVA)分析;若結果差異顯著時(P<0.05),則采用Student-Newmnan-Keuls法進行多重比較。檢驗結果以“平均值±標準誤(Mean±SE)”表示。
2 結果
2.1 飼料中牛磺酸含量對不同生長階段尼羅羅非魚幼魚生長性能的影響(見表2、表3)
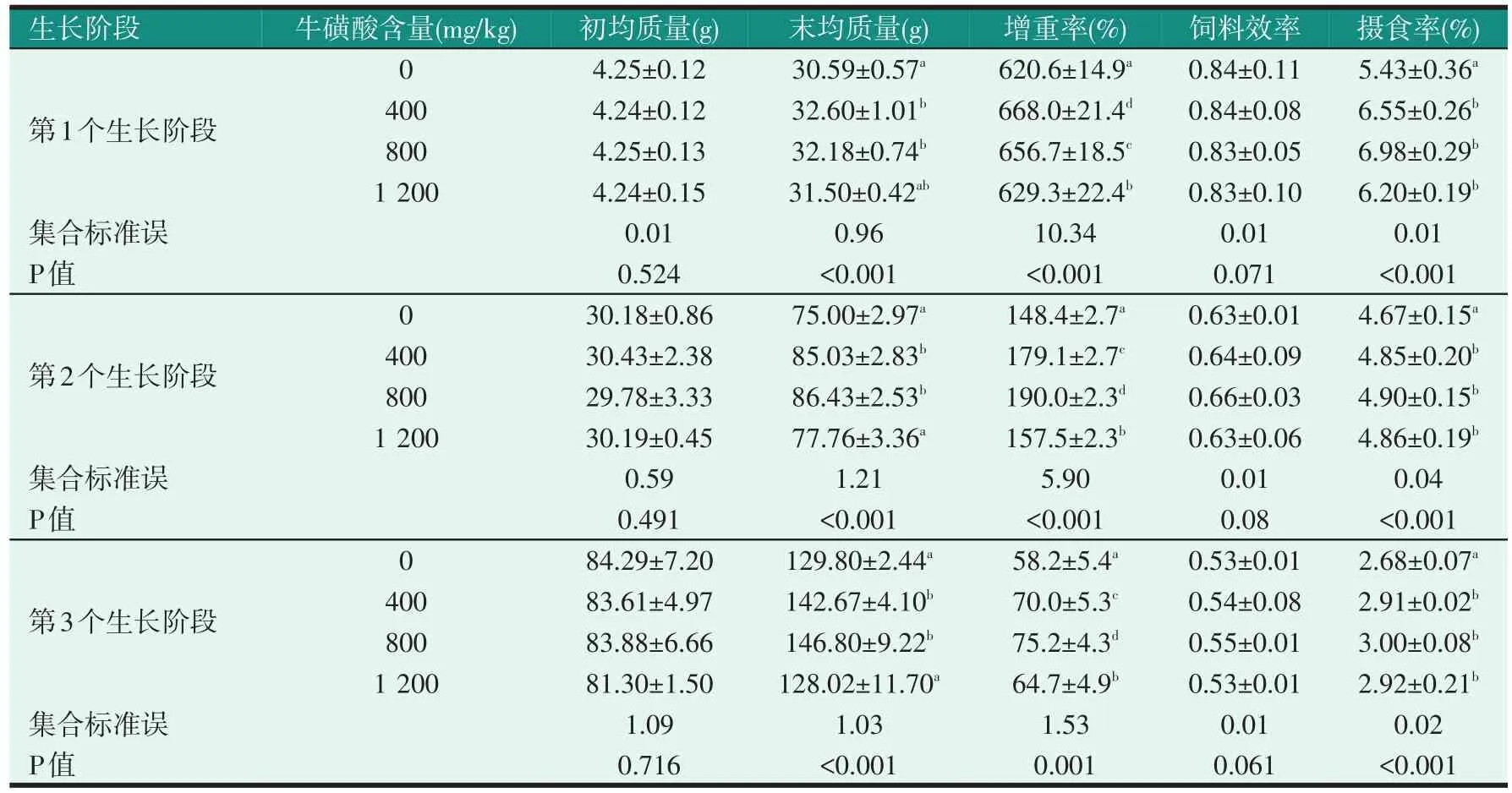
表2 飼料中牛磺酸含量對不同生長階段尼羅羅非魚幼魚生長性能的影響
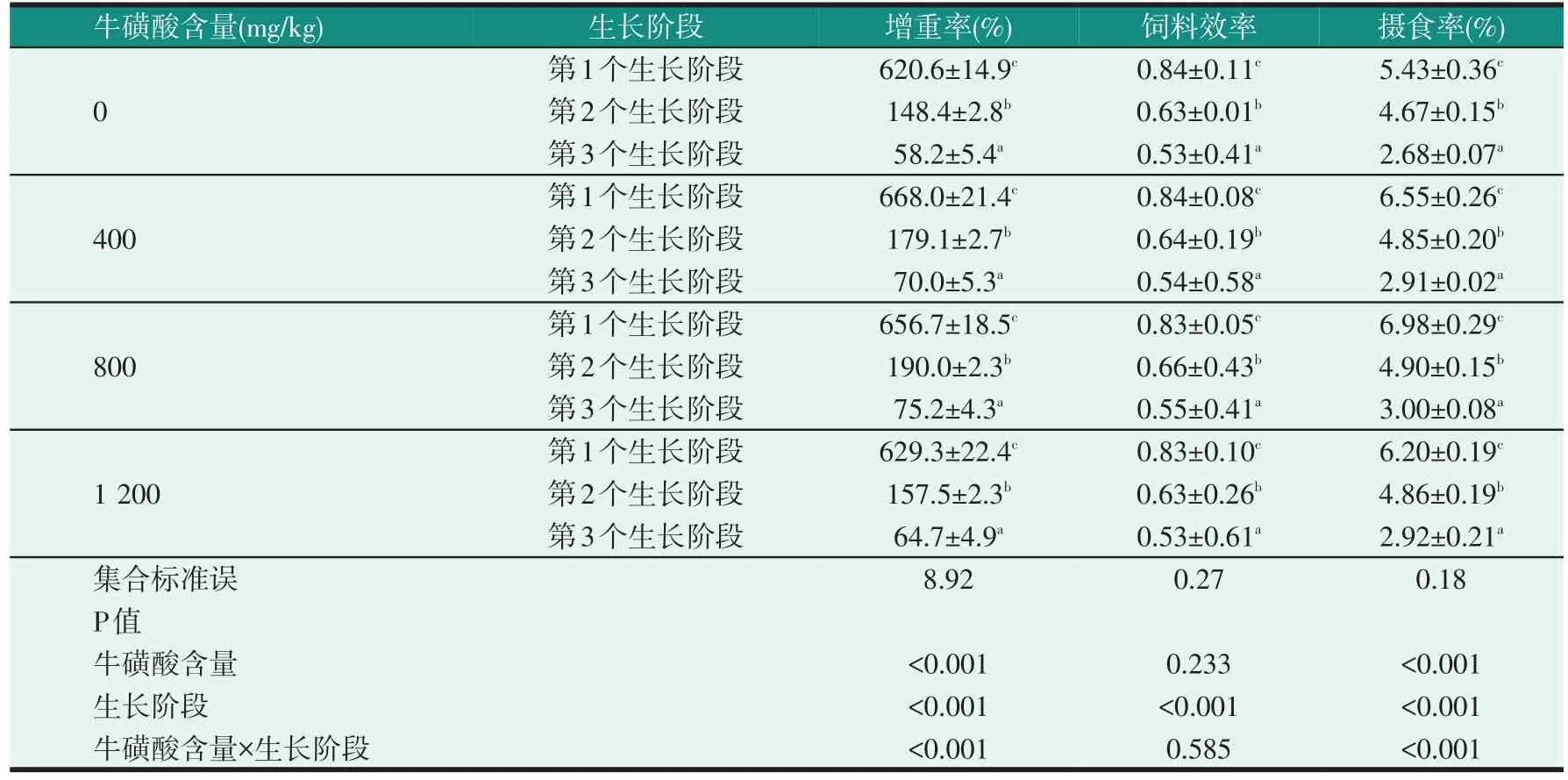
表3 飼料中牛磺酸含量和不同生長階段對尼羅羅非魚幼魚生長性能的影響

表4 飼料中牛磺酸含量(X)與尼羅羅非魚幼魚不同生長階段增重率(Y)的關系
由表2可知,在每一個生長階段中,飼料中添加牛磺酸對試驗魚的增重率和攝食率有明顯的影響。牛磺酸添加組的增重率均明顯高于對照組(P<0.05),且隨著牛磺酸含量的增加表現出先升高后降低的趨勢。牛磺酸添加組的攝食率明顯高于對照組(P<0.05),且牛磺酸添加組之間無明顯差異(P>0.05)。飼料效率不受飼料中牛磺酸含量的影響(P>0.05)。表3中,每個飼料中牛磺酸含量下,試驗魚的第1個生長階段的增重率、飼料效率和攝食率明顯高于第2個和第3個生長階段(P<0.05),飼料中牛磺酸含量和不同生長階段對增重率和攝食率有明顯的影響(P<0.05),并存在明顯交互作用。飼料效率不受牛磺酸含量的影響(P>0.05)。
為確定3個生長階段中尼羅羅非魚幼魚對飼料牛磺酸的需要量,我們建立了羅非魚增重率與飼料牛磺酸含量之間的回歸方程(見表4),擬合得到3個生長階段下尼羅羅非魚幼魚對牛磺酸的需要量分別為0.72%、0.68%和0.64%。由此可見,尼羅羅非魚魚齡越大,牛磺酸的需求量就越低。
2.2 飼料中牛磺酸含量對不同飼喂天數尼羅羅非魚幼魚形態學指標的影響(見表5、表6)
由表5可知,無論在哪個飼喂階段,試驗魚的肝體比和臟體比都受到飼料牛磺酸含量的顯著影響,添加了牛磺酸組的肝體比和臟體比明顯高于對照組(P<0.05),但各牛磺酸組的肝體比、臟體比沒有明顯差異(P>0.05);飼料中牛磺酸含量不影響肥滿度(P>0.05)。從表6可以看出,在每個飼料中牛磺酸含量下,試驗魚的肝體比和臟體比,在飼喂到第28 d時明顯高于飼喂到第56 d和第84 d時(P<0.05)。飼料中牛磺酸含量不影響肥滿度,但飼喂天數對其有顯著的影響(P<0.05),飼喂到第28 d的肥滿度明顯高于飼喂到第56 d和第84 d(P<0.05)。
2.3 飼料牛磺酸含量對飼喂84 d尼羅羅非魚幼魚體成分的影響(見表7)
該試驗測定了養殖結束時各牛磺酸處理組尼羅羅非魚的體成分。由表7可知,牛磺酸添加組魚體水分含量明顯低于對照組(P<0.05),但各牛磺酸添加組之間差異不明顯(P>0.05)。對照組魚體粗脂肪和粗蛋白質含量明顯低于牛磺酸添加組(P<0.05),且800 mg/kg牛磺酸組的魚體粗脂肪含量明顯高于其它組(P<0.05)。魚體粗灰分含量不受飼料中牛磺酸含量的影響(P>0.05)。
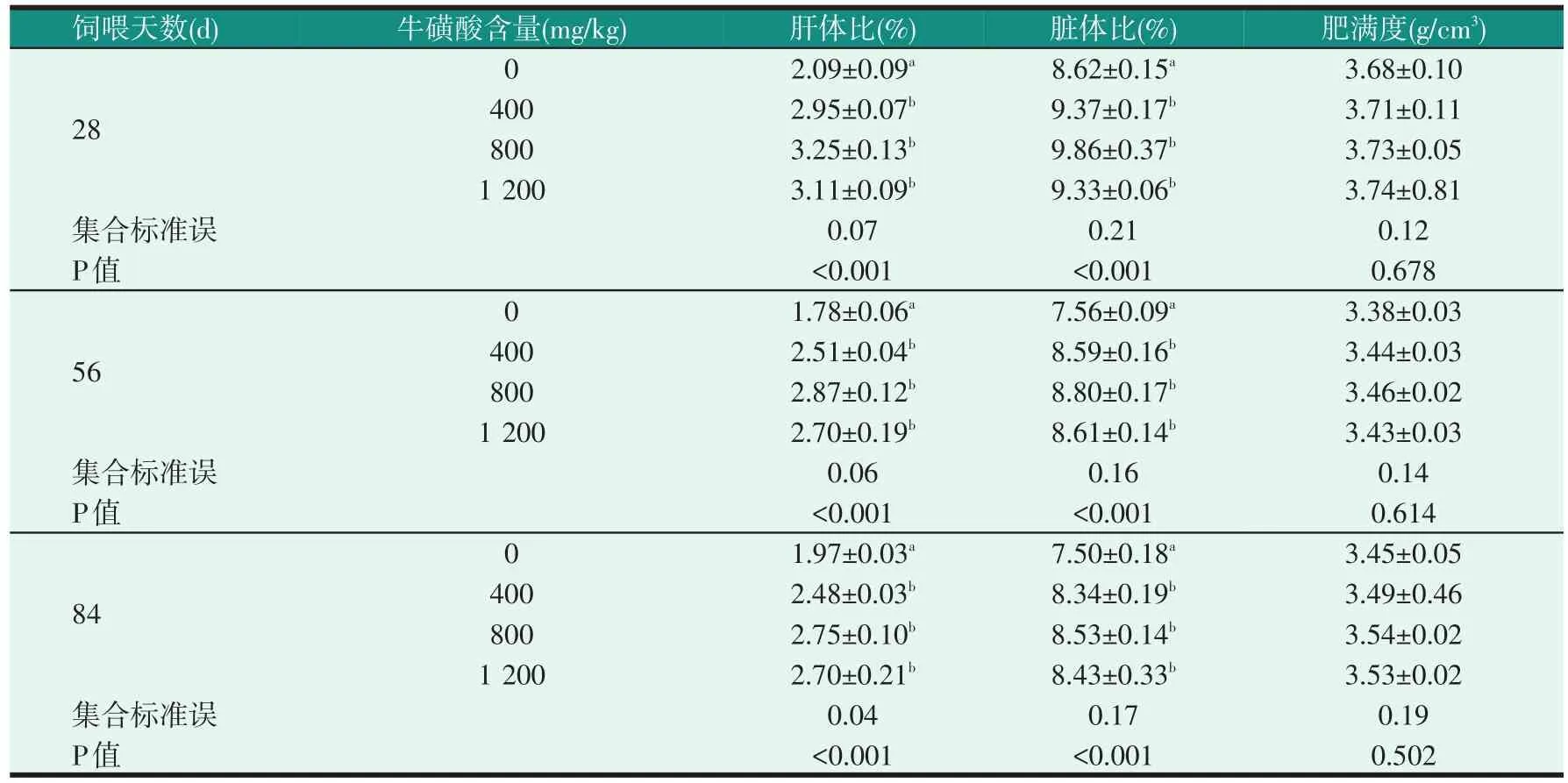
表5 飼料中牛磺酸含量對不同飼喂天數尼羅羅非魚幼魚形態指標的影響
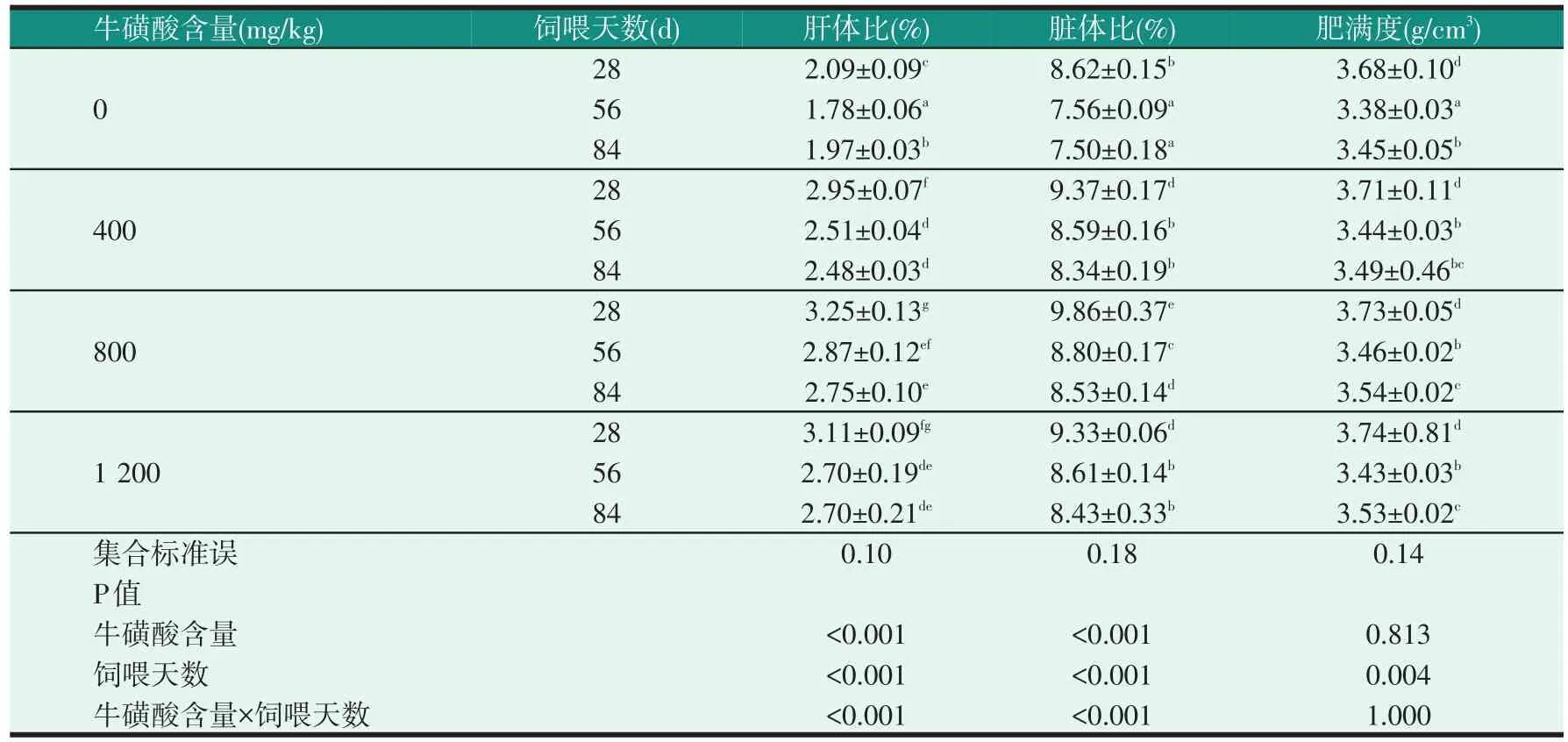
表6 飼料中牛磺酸含量和不同飼喂天數對尼羅羅非魚幼魚形態學指標的影響

表7 飼料中牛磺酸含量對飼喂84 d尼羅羅非魚體成分的影響
3 討論
已有研究顯示,飼料中添加牛磺酸能夠明顯促進魚類的生長[18-19]。該試驗結果表明,在飼料中添加一定含量的牛磺酸能夠使尼羅羅非魚幼魚的生長性能明顯提高,而當飼料中不添加牛磺酸時,其生長速度變緩,這與在真鯛[20]、牙鲆[11]和草魚[12]的試驗結果一致,表明飼料牛磺酸含量不足時會抑制魚類生長。在該試驗的3個生長階段中,隨著飼料中牛磺酸含量的增加,增重率均呈現先升高后降低的趨勢(見表2),這與在虹鱒[21]、斜帶石斑魚[7]、花鰻鱺(Anguillamarmorata)[22]、青魚(Mylopharyngodonpiceu)[23]的試驗結果類似。
在不考慮飼料中牛磺酸含量的情況下,3個生長階段中,尼羅羅非魚幼魚的生長速度以第1個生長階段為最大,第2個生長階段次之,以第3個生長階段為最慢(見表3),表明其生長速度隨飼喂天數的增加而降低,其生長趨勢符合魚類生長的規律。回歸分析擬合得出試驗魚3個生長階段的適宜牛磺酸含量分別為0.72%(第1個生長階段初始體質量為4.24 g)、0.68%(第2個生長階段初始體質量為30.14 g)和0.64%(第3個生長階段初始體質量為83.27 g)。由此可見,隨年齡增加,尼羅羅非魚幼魚對牛磺酸的需要量逐漸減少。雖然本試驗中尼羅羅非魚幼魚的生長速度從第1個生長階段到第3個生長階段是逐漸降低的,但它們仍是羅非魚養殖過程中生長速度較快的3個階段。
無論在試驗的哪個飼喂階段,飼料中添加牛磺酸明顯提高了尼羅羅非魚幼魚的攝食率,表明牛磺酸對尼羅羅非魚幼魚有一定的誘食作用(見表2),該結果支持了前期的發現[24]。但該試驗中各牛磺酸添加組的攝食率沒有明顯差異,說明在滿足牛磺酸需要的前提下,繼續增加牛磺酸含量沒有進一步促進尼羅羅非魚幼魚攝食。同時,每個生長階段的飼料效率均不受飼料中牛磺酸含量的影響。由此可以看出,牛磺酸并非是通過提高飼料效率而是通過增加攝食量來提高羅非魚的生長,該研究結果與在牙鲆[15]中的研究結果一致。然而,該試驗還發現,在飼料中牛磺酸含量過高(1 200 mg/kg牛磺酸),試驗魚的增重率有所下降,原因不明。有人認為飼料中添加過量的牛磺酸會引起其適口性下降[25],攝食量隨之降低,從而導致魚體生長速度下降。
牛磺酸的一個重要作用是可以與游離膽汁酸結合形成牛磺膽酸,促進脂類的消化吸收[26]。由試驗結果可知,牛磺酸添加組魚體粗脂肪含量明顯高于對照組(見表7),這與在草魚[12]、羅非魚[27]中的研究結果一致,但與虹鱒[14]、大菱鲆[28]、石斑魚[7]的結果相反。魚體粗脂肪增加的原因可能是飼料中添加的牛磺酸促進了甲狀腺激素T3分泌,而T3是戊糖磷酸循環中的重要調節因子,主要調節其關鍵酶1,6-磷酸脫氫酶,它的分泌促進了脂肪酸合成及其關鍵酶的轉錄,促進脂肪合成并在魚體沉積,最終表現為魚體粗脂肪增加[29]。在不考慮飼喂天數的情況下,試驗魚肝體比和臟體比是隨牛磺酸添加比例的增加而升高的,這與魚體粗脂肪的變化趨勢相似,這可能是牛磺酸與膽固醇結合形成膽鹽,進而激活了肝脂肪酶[30]。另外,在肝臟中,牛磺酸能和膽酸結合形成膽汁酸,促進肝臟脂肪的合成,最終導致肝臟脂肪沉積。該試驗中,添加牛磺酸組的魚體粗蛋白質含量都高于對照組,說明飼料中添加牛磺酸可以促進尼羅羅非魚幼魚體蛋白質的合成,該研究結果同大菱鲆[9]、細點牙鯛(Dentex dentex)[30]和塞內加爾鰨(Solea senegalensis Kaup)[31]一致,這可能是因為牛磺酸增強了蛋白質合成有關激素的分泌[32]。在3個生長階段中,第1個生長階段(1~28 d)的肝體比和臟體比均明顯高于第2個生長階段(29~56 d)和第3個生長階段(57~84 d),且后兩者之間差異不明顯。因此認為,尼羅羅非魚幼魚在第1個生長階段時身體各器官處于快速發展的階段,但從第2個生長階段起,魚體各器官的生理功能都趨向于成體,其生長發育也呈現出相對穩定的狀態。
4 結論
飼料中添加適量的牛磺酸能夠促進尼羅羅非魚幼魚的生長。尼羅羅非魚幼魚在3個生長階段所需的牛磺酸含量分別是0.72%(第1個生長階段初始體質量為4.24 g)、0.68%(第2個生長階段初始體質量為30.14 g)和0.64%(第3個生長階段初始體質量為83.27 g),說明隨著年齡增加,尼羅羅非魚幼魚對牛磺酸的需求量逐漸降低。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
