大豆異黃酮對褐牙鲆頭腎巨噬細胞免疫相關基因表達的體外研究
2018-12-11 01:31:52陳京華王正麗
水產科學 2018年6期
姜 秦,楊 寧,袁 瑞,陳京華,王正麗
( 青島農業大學 海洋科學與工程學院,山東 青島 266109 )
褐牙鲆(Paralichthysolivaceus)是我國主要的海水養殖魚類之一,具有很高的經濟價值,在我國北方大面積養殖。但隨著集約化養殖的發展,由于養殖密度高、投餌頻率增加及水體污染等原因,褐牙鲆感染疾病的幾率大大增加,病害問題已成為制約褐牙鲆養殖業健康發展的瓶頸。探討免疫防治新途徑,提高魚體自身免疫力,是實現褐牙鲆健康養殖的可靠保障。而通過營養調控提高魚類的免疫力,已被證實是一種行之有效的方法[1]。
大豆異黃酮是來自大豆的主要植物雌激素[2],是一種具有類雌激素性質的抗營養因子[3],現有研究顯示,在陸生動物中,大豆異黃酮無論在體內還是體外都具有免疫調節作用[4-6]。魚類中相關研究較少,有研究表明,飼料中添加低含量大豆異黃酮能夠促進異育銀鯽(Carassiusauratusgibelio)生長[7];飼料中添加適量的大豆異黃酮可以增強卵形鯧鲹(Trachinotusovatus)免疫活性,降低死亡率[8]。但大豆異黃酮對褐牙鲆免疫效應研究相對較少,國內僅見大豆異黃酮對褐牙鲆免疫細胞功能的體外研究[9]。白細胞介素(IL)是由活化的單核——巨噬細胞和淋巴細胞等產生的作用于淋巴細胞、巨噬細胞或者其他細胞的細胞因子[10],在免疫活動中進行信號傳遞、聯絡白細胞群等功能。免疫球蛋白(Ig)是蛋白質的一種,在魚體受到外界刺激之后,由體內的免疫系統產生[11]。免疫球蛋白的產生機制是在體內接觸過曾經對機體具有刺激作用的物質之后,產生一種可以進行抵抗的蛋白質類抗體[12]。干擾素(IFN)由單核吞噬細胞高度分裂分化之后的巨噬細胞產生,通過干擾外來刺激物在體內的分裂分化和結合過程,進而發揮免疫調節作用[13]。熱休克蛋白(HSPs)是一種短期內產生但具有高效作用的蛋白質。熱休克蛋白的作用機制是與體內免疫系統分泌的淋巴細胞進行聯合,一起作用于外來物質,起到免疫預防的功能[14]。本研究分離褐牙鲆頭腎巨噬細胞,在體外培養條件下,研究不同質量濃度大豆異黃酮對褐牙鲆4種免疫相關基因相對表達量的影響,探討大豆異黃酮的免疫作用,以期為大豆異黃酮作為廉價的免疫增強劑提供基礎資料。
1 材料與方法
1.1 試驗材料
試驗用褐牙鲆購自日照某養殖場,體質量(250±12) g,健康,活力好。暫養于水循環系統。暫養期間水溫17~19 ℃,鹽度32~40,溶解氧>8.0 mg/L,每日飽食投喂2次。
1.2 試驗方法
1.2.1 頭腎巨噬細胞的分離
褐牙鲆頭腎巨噬細胞的分離參考文獻[15]的方法。在無菌的條件下,剖取褐牙鲆頭腎,研磨并通過100目篩絹,將研磨液轉移到34%/51% Percoll分離液表層,4 ℃,2200 r/min離心25 min,吸取中間層巨噬細胞,加入吸取量3倍體積的0.15 mol/L磷酸緩沖鹽溶液,1200 r/min離心5 min,洗滌2次,加入2% FBS-L-15(雙抗1%)重懸細胞。用臺盼藍檢測調整細胞密度至2×107個/mL,按600 μL/孔接種到24孔板中,在20 ℃條件下,恒溫培養2 h使細胞貼壁備用。
1.2.2 頭腎巨噬細胞的培養
頭腎巨噬細胞貼壁培養2 h后,用0.15 mol/L磷酸緩沖鹽溶液洗去未貼壁細胞,再分別加入大豆異黃酮質量濃度為0、0.1、0.5、1.0、1.5、2.0 mg/mL的5%FBS-L-15細胞培養液,且每個梯度設置3個平行,放于20 ℃恒溫培養箱培養,分別在培養后6、12、24 h收集細胞,提取RNA。
添加脂多糖刺激時,吸棄未貼壁的細胞及培養液,分別加入大豆異黃酮質量濃度為0、1.0 mg/mL的5%FBS-L-15細胞培養液,并加入脂多糖使其終質量濃度為100 ng/mL,刺激1 h后,更換為不含脂多糖的大豆異黃酮細胞培養液,培養方法同上,分別在不含脂多糖的培養液中培養3、6、12、24 h后收集細胞,提取RNA。
1.2.3 熒光定量PCR測定
吸棄培養液,收集貼壁細胞,然后采用Trizol法提取細胞RNA,按照康為世紀HiFiScript反轉錄試劑盒要求合成第一鏈cDNA,以此為模板進行熒光定量PCR。以-actin為內參基因,RT-PCR反應體系及過程參照Takara PrimeScript熒光定量試劑盒。引物序列見表1。
1.2.4 數據分析
試驗所得Ct值采用2-ΔΔCt法計算各基因在不同質量濃度大豆異黃酮培養下的相對表達量。試驗結果用平均值±標準差表示,所有數據用SPSS 19.0軟件進行分析,兩組數據間比較采用t檢驗,多組數據間比較采用單因素方差分析,并進行Duncan多重比較,P<0.05表示差異顯著。

表1 引物序列
2 結 果
2.1 大豆異黃酮體外對褐牙鲆頭腎巨噬細胞IL-1相對表達量的影響
研究結果顯示,大豆異黃酮在體外能夠顯著促進褐牙鲆頭腎巨噬細胞的IL-1基因相對表達(P<0.05),且促進作用隨著培養時間的延長而增強(圖1)。用添加大豆異黃酮的培養液培養6 h后,0.5、1.0、1.5 mg/mL質量濃度組的IL-1基因相對表達量顯著高于對照組(P<0.05);培養12 h后,添加大豆異黃酮試驗組的IL-1基因相對表達量均顯著高于對照組(P<0.05),且1.0、1.5 mg/mL試驗組顯著高于其他大豆異黃酮添加組;至24 h,大豆異黃酮質量濃度不低于0.5 mg/mL各試驗組IL-1的相對表達量顯著高于對照組(P<0.05),且1、1.5 mg/mL試驗組的IL-1基因相對表達量最高。即在試驗梯度范圍內,培養液中大豆異黃酮質量濃度為0.5~1.5 mg/mL時,能夠顯著增強IL-1基因的相對表達量。
2.2 大豆異黃酮體外對褐牙鲆頭腎巨噬細胞IFN-相對表達量的影響

圖1 大豆異黃酮對褐牙鲆頭腎巨噬細胞IL-1的影響不同小寫字母表示差異顯著(P<0.05),下同.

圖2 大豆異黃酮對褐牙鲆頭腎巨噬細胞IFN-的影響
2.3 大豆異黃酮體外對褐牙鲆頭腎巨噬細胞IgM相對表達量的影響
離體條件下大豆異黃酮能夠顯著增強褐牙鲆頭腎巨噬細胞IgM基因相對表達(P<0.05)(圖3)。培養6 h后,培養液中添加0.5、1.0 mg/mL大豆異黃酮試驗組的IgM基因相對表達量顯著高于對照組(P<0.05);培養12 h后,大豆異黃酮為1.0、1.5 mg/mL試驗組的IgM基因相對表達量顯著高于對照組及其他各組(P<0.05);至24 h后,0.5、1.0 mg/mL大豆異黃酮試驗組的褐牙鲆IgM基因相對表達量顯著高于對照組和其他各組(P<0.05)。即在試驗梯度范圍內,培養液中大豆異黃酮質量濃度為0.5~1.0 mg/mL時,在體外能夠顯著增強褐牙鲆頭腎巨噬細胞IgM基因的相對表達量。
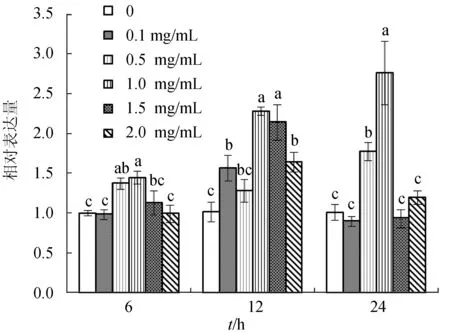
圖3 大豆異黃酮對褐牙鲆頭腎巨噬細胞IgM的影響
2.4 大豆異黃酮體外對褐牙鲆頭腎巨噬細胞HSP70相對表達量的影響
離體條件下,培養液中大豆異黃酮能夠顯著增強褐牙鲆頭腎巨噬細胞HSP70基因相對表達(P<0.05)(圖4)。培養6 h和12 h后,大豆異黃酮質量濃度為0.5 mg/mL及以上試驗組褐牙鲆頭腎巨噬細胞中HSP70基因的相對表達顯著高于對照組(P<0.05);至24 h后,大豆異黃酮質量濃度為0.5、1.0、1.5 mg/mL的試驗組HSP70的相對表達量顯著高于對照組和其他各添加組(P<0.05)。即在試驗梯度范圍內,體外培養液中大豆異黃酮質量濃度為0.5~1.5 mg/mL時,能夠顯著增強頭腎巨噬細胞HSP70基因的相對表達量。

圖4 大豆異黃酮對褐牙鲆頭腎巨噬細胞HSP70的影響
2.5 脂多糖刺激下大豆異黃酮離體對褐牙鲆頭腎巨噬細胞免疫相關基因的影響
研究結果顯示,在脂多糖刺激后,添加1.0 mg/mL大豆異黃酮培養的褐牙鲆頭腎巨噬細胞4種免疫相關基因隨刺激后培養時間的變化趨勢與未添加大豆異黃酮組基本一致,但是在某些時間點,其相對表達量顯著增加。在脂多糖刺激后繼續培養6、24 h時,大豆異黃酮添加組的褐牙鲆頭腎巨噬細胞的IL-1基因表達量顯著高于對照組(P<0.01);脂多糖刺激后繼續培養24 h時,大豆異黃酮添加組IFN-基因的表達量顯著高于對照組(P<0.05);IgM基因則在6 h時顯著高于對照組(P<0.01);而大豆異黃酮添加組的HSP70基因自脂多糖刺激繼續培養3 h開始,其相對表達量均顯著高于對照組(P<0.01)。

3 討 論
褐牙鲆的頭腎巨噬細胞是固有免疫的效應細胞,在機體免疫防御過程中起重要作用。IL-1、IFN-、IgM和HSP70是4種比較典型的免疫相關基因,均在褐牙鲆頭腎巨噬細胞中表達,因此本試驗選擇離體條件下用不同質量濃度大豆異黃酮培養褐牙鲆頭腎巨噬細胞,在不同時間測定上述4種基因的相對表達量,以此探討大豆異黃酮與免疫的相關性。研究結果顯示,大豆異黃酮能夠顯著影響這4種基因的相對表達,且不同基因在大豆異黃酮不同質量濃度、不同作用時間均存在顯著差異。
脂多糖是革蘭氏陰性菌細胞壁的成分之一,具有很強的免疫原性,可以調控細胞的信號通路,影響相關基因的表達[20]。李勝亮等[21]研究顯示,脂多糖刺激可以導致巨噬細胞IL-1釋放增多;于東云等[22]研究顯示,HSP70在脂多糖誘導的肺泡上皮細胞中應激表達,認為HSP70對肺損傷起保護作用。本研究在脂多糖刺激條件下,比較研究含1.0 mg/mL大豆異黃酮組與不含大豆異黃酮的對照組的免疫基因表達特點,結果發現,脂多糖刺激后,4種基因表達隨時間變化而變化,而添加質量濃度為1.0 mg/mL大豆異黃酮組與對照組的變化趨勢基本一致。但是試驗組IL-1基因表達在6 h和24 h顯著高于對照組;IFN-基因在24 h顯著高于對照組;IgM基因在6 h顯著高于對照組;而HSP70基因自3 h開始均顯著高于對照組。這些變化表明,在受到免疫刺激的情況下,大豆異黃酮可以提高IL-1、IFN-、IgM和HSP70基因的相對表達量,可能提高機體免疫力。
綜上所述,在本試驗條件下,培養液中大豆異黃酮質量濃度為0.5~1.5 mg/mL時,能夠顯著上調體外培養條件下褐牙鲆頭腎巨噬細胞IL-1、IFN-、IgM和HSP70基因的相對表達,從而增強褐牙鲆頭腎巨噬細胞的免疫力。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
