狐源大腸桿菌毒力基因的檢測
2018-11-23 03:04:42王紹紅李政志張艷芳劉偉石
野生動物學報 2018年4期
王紹紅 柴 榮 李政志 張艷芳 劉偉石 薛 原 孫 穎
(東北林業大學野生動物資源學院,哈爾濱,150040)
大腸桿菌(Escherichiacoli)作為一種機會致病菌,含有多種毒力因子,這些毒力因子對分析大腸桿菌的致病性,研究其是否存在潛在的致病能力等方面起著至關重要的作用,因此檢測相關毒力基因具有重要意義。毒力基因廣泛存在于染色體,毒力島和質粒上[1],是否存在水平轉移現象現在還未可知,另毒力基因存在變異現象,因此研究毒力基因的發展,分析其致病性,進行毒力基因檢測,研究其內在聯系,對于疾病的預防以及臨床指導用藥具有十分重要的意義。
耶爾森菌強毒力島(high island,HPI)常出現在大腸桿菌強毒力菌株中,是主要由irp1、irp2、irp3、irp4、irp5、fyuA、ybtA等基因組成的毒力基因簇。HPI含有編碼攝取合成Ybt相關基因,與鐵攝取與調節有關,而鐵元素是保證細菌在宿主體內存活的關鍵因素,因此攜帶有HPI毒力島的菌株可攝取鐵元素,易在宿主體內存活,致病,具有較強的毒力,而且已有研究表明HPI毒力島可在兩種細菌間水平轉移,因此,HPI毒力島的研究對大腸桿菌致病性的研究具有重要意義。另,大腸桿菌的致病性也與黏附素、毒素、侵襲性酶以及多種因子有關[2]。
本研究采用PCR法對135株東北地區狐源大腸桿菌進行irp2,fyuA,ybtA,iucD,iss,fimH種毒力基因的檢測并進行克隆測序,研究種毒力基因出現的頻率,為后續毒力因子流行情況的研究提供試驗數據。
1 材料與方法
1.1 菌株
本實驗檢測東北地區狐源大腸桿菌共135株,分離的菌株及經過生理生化鑒定為大腸桿菌。
1.2 主要試劑
膠回收試劑盒,質粒抽提試劑盒均購于愛思進公司;瓊脂糖,LB,50×TAE緩沖液,DL2000 DNA maker,TaqDNA聚合酶,DH5α和pMD-18T等均購自寶生物生物技術有限公司。
1.3 引物合成
根據相關文獻記載的大腸桿菌HPI毒力島主要結構基因及其相關基因合成對引物,分別用于irp2,fyuA及fimH,iucD,iss,ybtA種毒力基因的擴增,引物由博仕生物有限公司合成。引物序列見表1。
1.4 PCR法擴增
irp2,fyuA及fimH,iucD,iss,ybtA基因。
表1 大腸桿菌毒力基因的引物

Tab.1 Primers for E.coli virulence genes
1.5 PCR產物電泳鑒定
取PCR產物5 μL,進行1%瓊脂糖凝膠電泳,以DNA Maker DL2000為參照,EB染色后置于紫外燈下觀察。
2 結果
2.1 單個毒力基因的檢測結果
對135株大腸桿菌進行PCR檢測:共57株檢出irp2,陽性率為42.22%;11株檢出ybtA,陽性率為 8.15%;13株檢出fyuA,陽性率為9.63%;56株檢出fimH,陽性率為41.48%;17株檢出iucD,陽性率為12.59%;1株檢出iss,陽性率為0.07%(圖1~6)。

圖1 irp2基因片斷的PCR擴增產物Fig.1 PCR amplification product of the irp2 gene fragmentM:DNA Makers DL2000Lane1-5:The positive strains for target genes
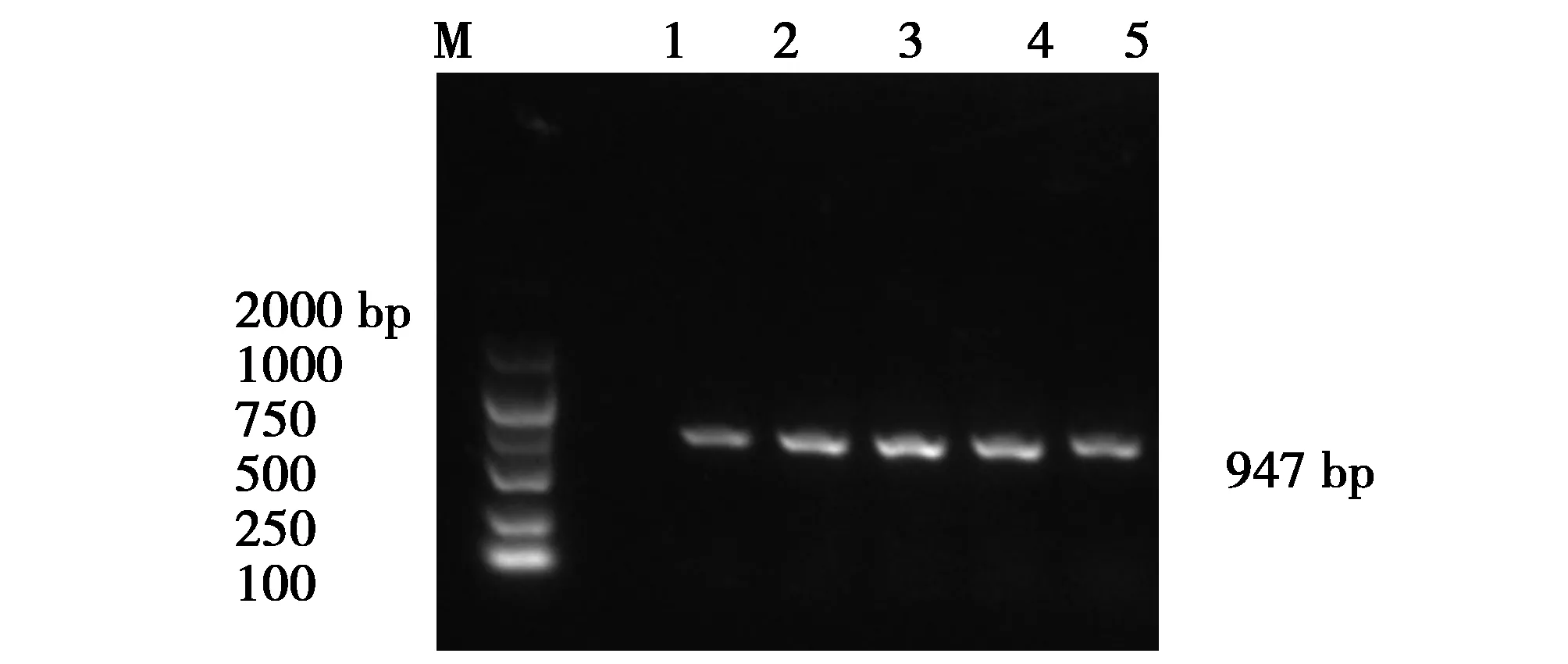
圖2 fyuA基因片斷的PCR擴增產物Fig.2 PCR amplification product of the fyuA gene fragmentM:DNA Makers DL2000Lane1-5:The positive strains for target genes
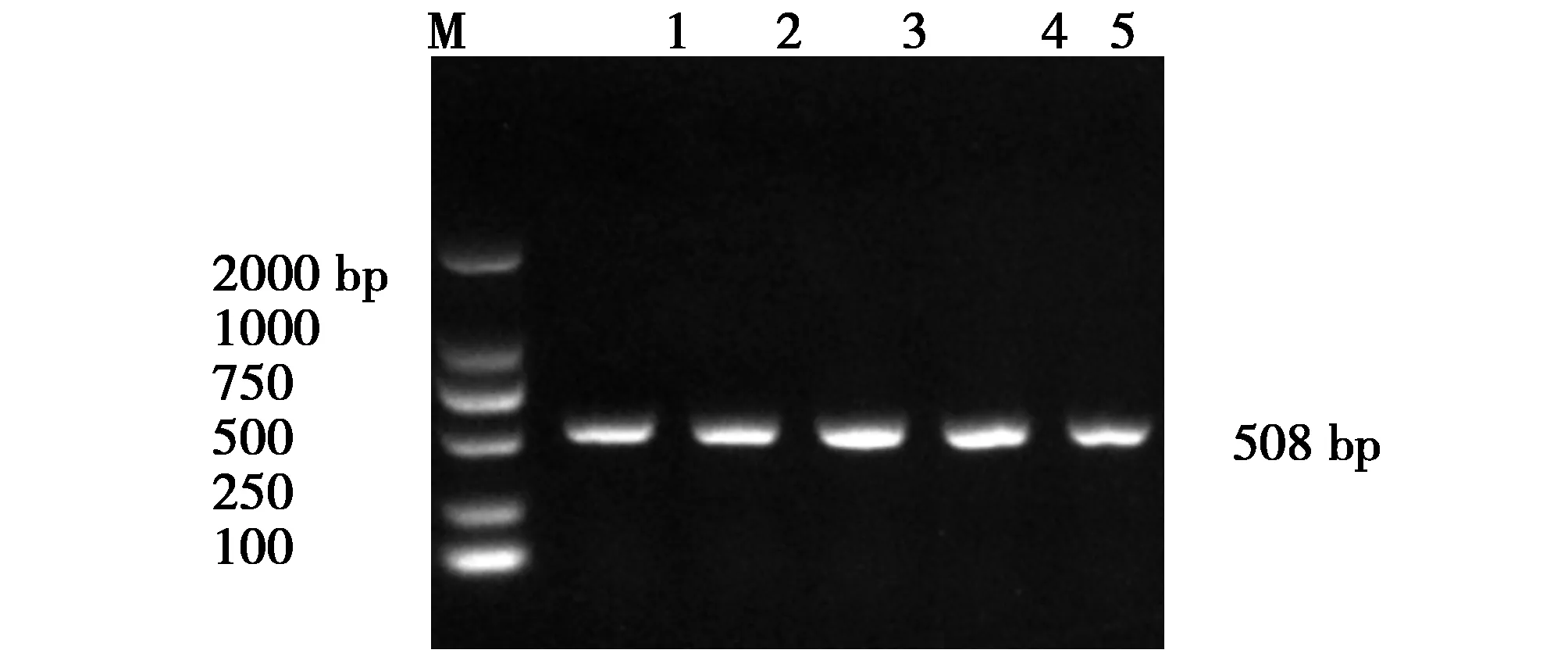
圖3 fimH基因片斷的PCR擴增產物Fig.3 PCR amplification product of the fimH gene fragmentM:DNA Makers DL2000Lane1-5:The positive strains for target genes
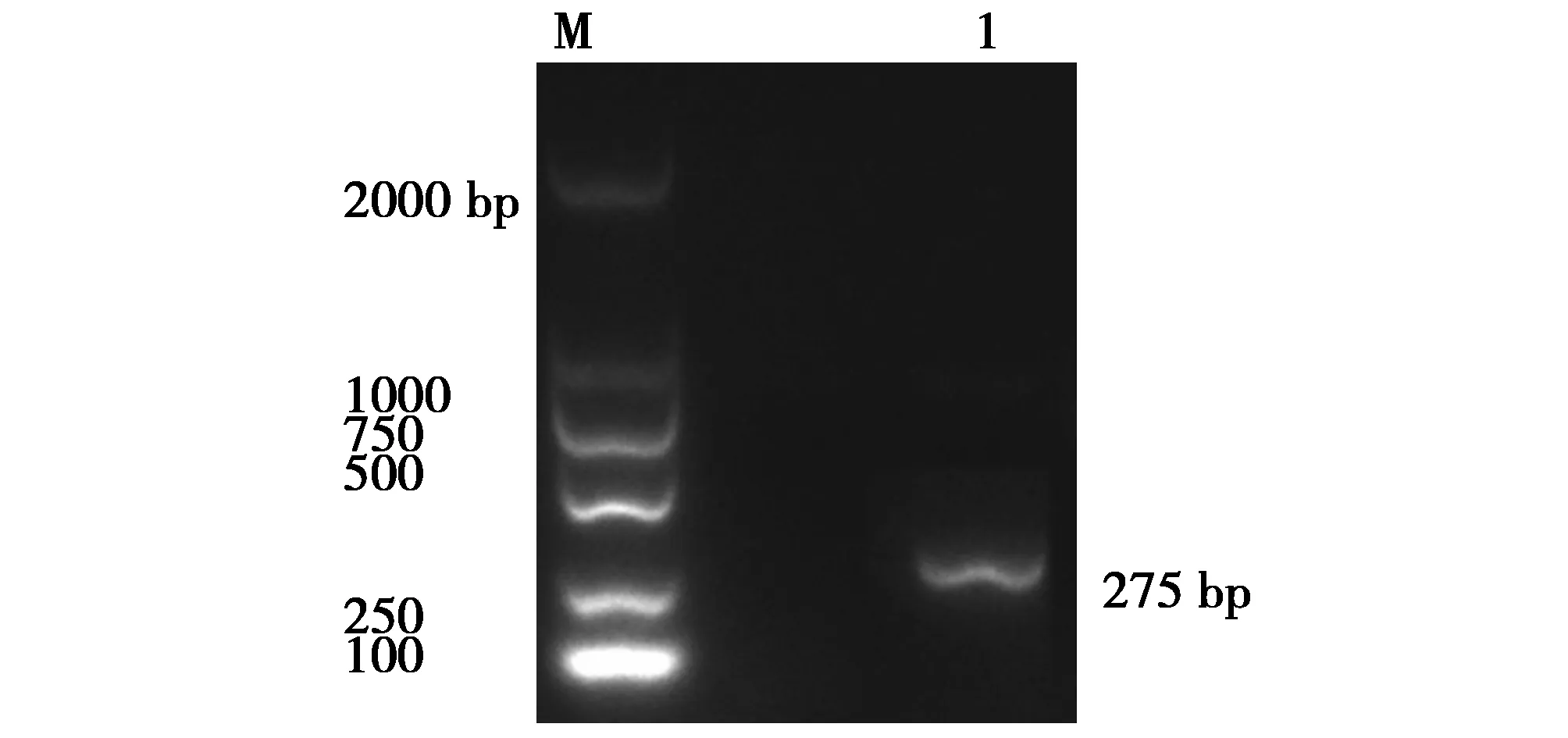
圖4 iss基因片斷的PCR擴增產物Fig.4 PCR amplification product of the iss gene fragmentM:DNA Makers DL2000Lane1:The positive strains for target genes
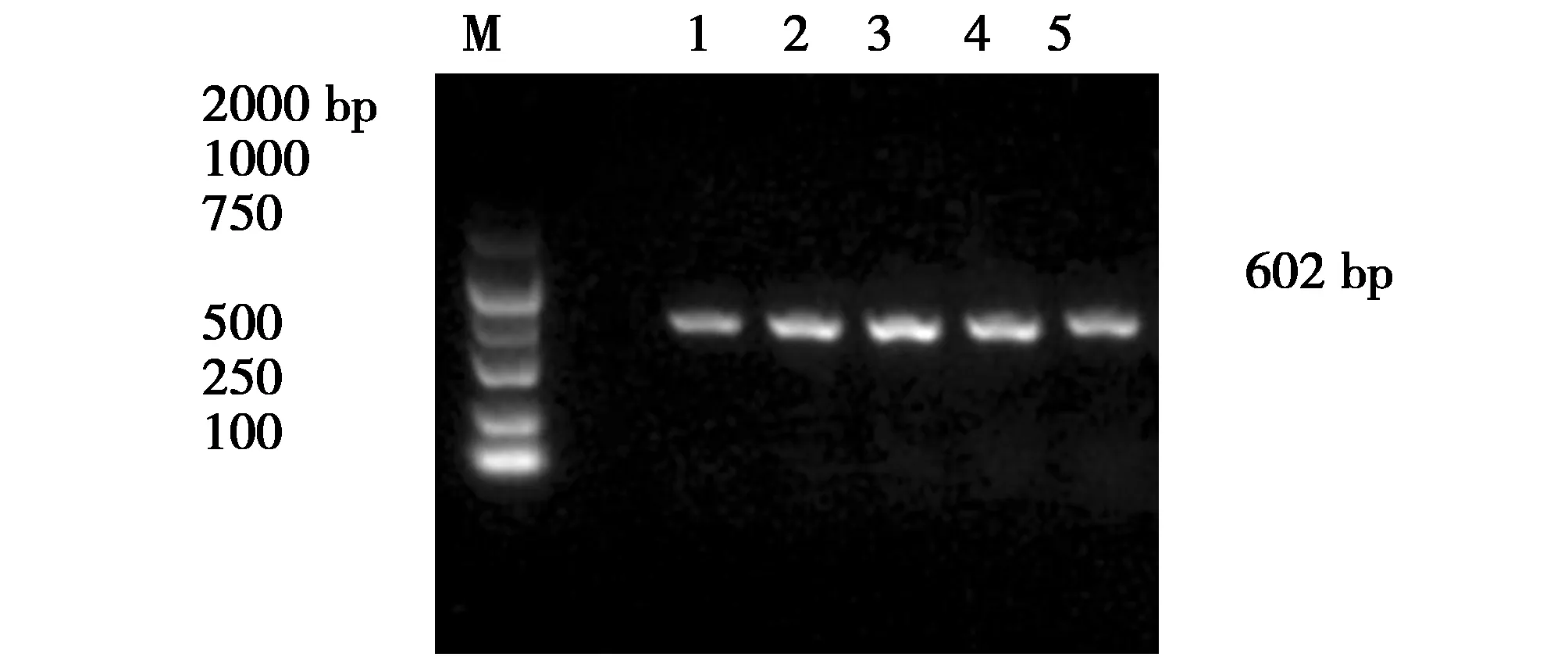
圖5 iucD基因片斷的PCR擴增產物Fig.5 PCR amplification product of the iucD gene fragmentM:DNA Makers DL2000Lane1-5:The positive strains for target genes
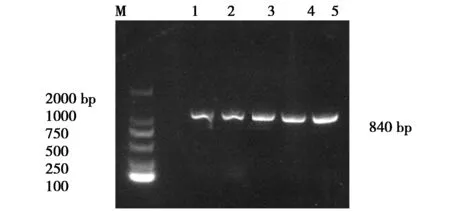
圖6 ybtA基因片斷的PCR擴增產物Fig.6 PCR amplification product of the ybtA gene fragmentM:DNA Makers DL2000Lane1-5:The positive strains for target genes
2.2 2-3個毒力基因的檢出結果
由表2可知,Irp2,fyuA以及fimH,ybtA和Irp2,fyuA,fimH分別同為陽性的菌株均有3株,分別占2.22%;Irp2,fimH同為陽性的菌株有8株,占5.93%;fimH,fyuA同為陽性的菌株有2株,占1.48%;Irp2,iucD同為陽性的菌株有9株,占6.66%;fimH,iucD同為陽性的菌株有4株,占2.96%;Irp2,ybtA同為陽性的菌株有6株,占4.44%;fyuA,iucD以及Irp2,iucD,fimH分別同為陽性的菌株均有1株,分別占0.74%,未發現有同時攜帶3個以上毒力基因的菌株。
表2 毒力基因檢測結果統計
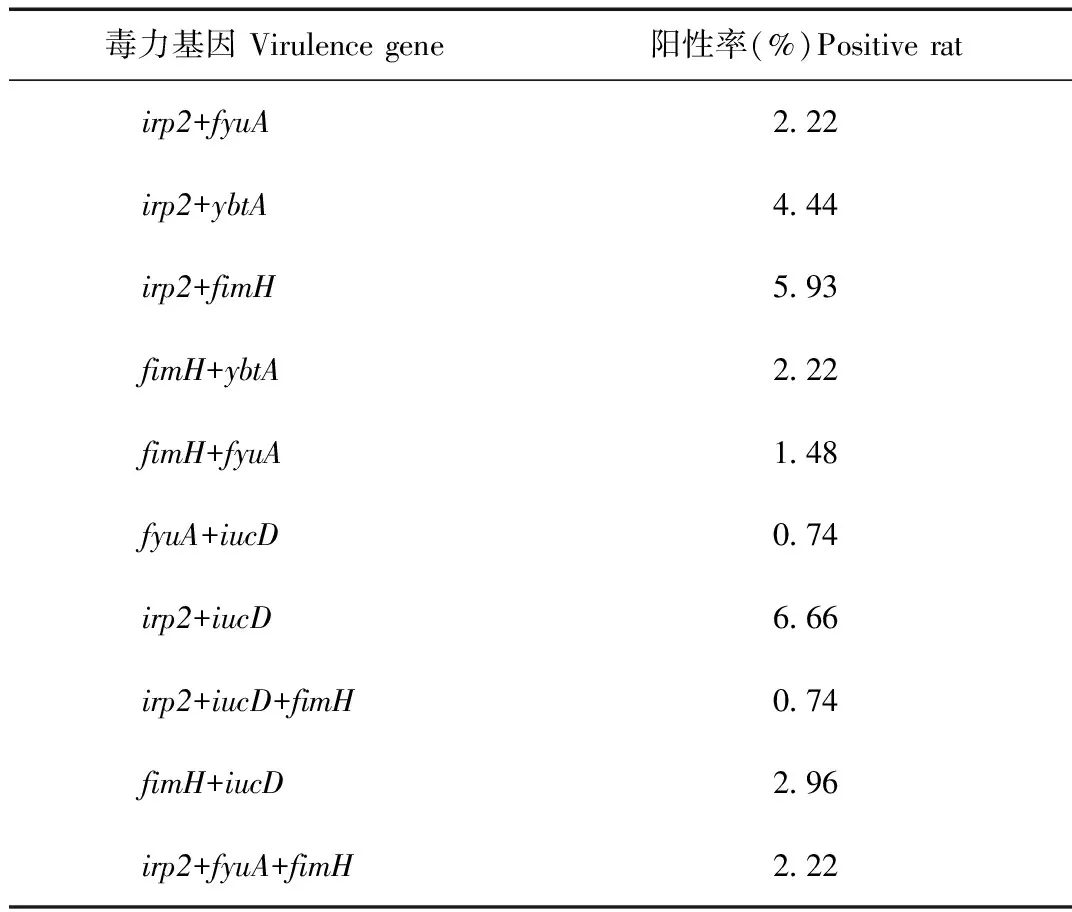
Tab.8 Virulence gene test results statistic
3 討論
大腸桿菌致病性主要與其菌毛、毒素、抗血清活性因子、侵襲性酶及鐵獲取系統和生物膜形成的相關蛋白有關,它們提供了定植的能力,以及形成生物膜,捕獲鐵等基本營養物質,造成上皮損傷,逃避宿主的防御機制,從而增強了大腸桿菌引起全身感染的能力。其中大腸桿菌產生的特定的毒力因子是決定其致病性的關鍵因素。
實驗檢測了6種毒力基因,編碼不同的毒力因子,分別在大腸桿菌侵入,定植,保護細菌免受宿主細胞自噬機制的識別和捕獲,以及營養、分泌毒素的過程中起著重要的作用。
其中,fimH(黏附素)能夠暴露于細菌表面而不是菌毛的組成部分,是菌毛的次要成分,對D-甘露糖受體結合起著獨特的作用[7]。可賦予1型菌毛結合D-甘露糖,并因此結合許多類型的真核細胞(包括腸、肺、膀胱、腎上皮和各種炎癥細胞)的能力對保護大腸桿菌免受吞噬有直接或間接的作用[8]。
實驗結果顯示作為HPI毒力島主要結構基因的irp2和黏附相關基因fimH的檢出率最高,同樣,在趙李祥的研究中fimH檢出率也非常高,達到90%以上[9]。在鐘杏好等的研究中,fimH檢出率也高達87.1%~91.3%[10],與本實驗結果一致,而fimH檢出率普遍很高,這是否說明這個基因比較穩定,以及其在大腸桿菌的生存與定植過程中是否發揮著不可或缺的作用,是否所有有定植能力的大腸桿菌都有這種毒力基因,種種問題還有待進一步驗證。
iss(保護素)是pTJ100相關基因,是在大多數APEC菌株中發生的一組質粒相關基因[11],賦予細菌攝取鐵和抗血清的作用,編碼與組織侵襲相關的毒力因子,與鐵的攝取與調節有直接或間接的關系[12],在禽類菌株中更常見,而iucD在人類菌株中更為普遍[12],在本次實驗中iss的檢出率僅為0.07%,iucD檢出率卻為12.59%,另外,兩種基因檢出率都不高與Ewers等的82.7%、78.0%有較大出入[13],iss檢出率也不高于Ragione等研究中的88.7%[8],而相較于呂殿紅等人研究的28.7%也有一定的差距[14],Bonnet等認為iss為禽致病性大腸桿菌的標志基因之一[2],而在本實驗中iss毒力基因的檢出率要明顯低于其他基因且唯一檢出的1株恰好沒有其他5種毒力基因。
另外,在此次研究中,有7個分離株同時攜帶3種毒力基因,占全部菌株的5.19%,47個菌株同時攜帶2種毒力基因,占全部菌株的34.81%,并未發現有攜帶更多毒力基因的菌株,根據Wang等的研究,大腸桿菌菌株的致病性與毒力相關基因的數量和組合模式密切相關[15],因此,這里還可以做進一步的研究。
此外,在耶爾森氏菌的鐵代謝的高致病性島(HPI)中,小鼠模型顯示HPI增加了腸外感染中的大腸桿菌毒力[16]。fyuA,irp2,ybtA屬于HPI毒力島的核心和功能基因,是它的的3個啟動區。其中ybtA屬Arac轉運調節家族,促進fyuA、irp2、irp6等啟動子的表達抑制自身啟動子[4],對Ybt的合成攝取起調節作用[16]。irp2與攝鐵能力有關,參與鐵載體的合成和表達,可作為HPI的標志基因;fyuA可編碼鐵抑制外膜蛋白,是Ybt細菌素及巴氏桿菌素的外膜受體,與大腸桿菌毒力密切相關。
實驗結果表明,同為HPI毒力島主要結構基因,同時攜帶irp2與fyuA基因的菌株卻少于同時攜帶有irp2與fimH的菌株且fyuA的檢出率低于irp2。事實上,HPI毒力島不完整的現象在胡靜等人的研究中曾有記載,fyuA在HPI核心區的邊緣,在進化和轉移過程中容易破壞和丟失,所以fyuA的檢出率往往低于irp2[17],且在HPI核心區中除邊緣丟失的情況外,還存在其他形式的缺失。且在胡靜的實驗中,EAggEC菌株所攜帶的與耶爾森菌同源的HPI毒力島,即便HPI存在,正常情況下并不表現強毒力和生長抑制,也并不意味著細菌獲得了致病性,猜想與菌種的遺傳差異有關,那么是否HPI在水平轉移過程中也存在這種情況,那么HPI僅存在于強毒力菌株是否還成立呢?另外,irp2與fimH,irp2與iucD同時存在的菌株數頗多是否是偶然,是否說明irp2與fimH,iucD之間存在某種關聯還有待深入研究。
研究表明,大量的菌株可形成一個重要的毒力基因庫,在某些情況下,并與其他因素結合,可能會產生新的致病菌株。新的致病形式的大腸桿菌往往通過不同毒力基因的組合而出現,而大腸桿菌也能夠將毒力基因輸出到其他革蘭氏陰性生物。因此,檢測環境中的大腸桿菌毒力基因是至關重要的。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
