老齡大鼠聽功能下降與線粒體DNA4834bp缺失突變的關系
2018-11-22 01:48:22黃海林劉俊
浙江臨床醫學 2018年9期
黃海林 劉俊?
老年性耳聾是老年人中發生率較高的慢性疾病,是衰老在聽覺器官的表現。研究表明,衰老與線粒體功能紊亂密切相關,而線粒體功能紊亂是線粒體DNA突變引起的。Bai等[1]報道mtDNA4977缺失與老年性耳聾存在可能相關性。Fischel-Ghodsian等[2]檢測5例老年性耳聾患者膜迷路和螺旋神經節中mtDNA編碼的細胞色素氧化酶Ⅱ基因,發現老年性耳聾患者mtDNA突變率高于老年對照組,但突變發生的部位和數量均存在較大個體差異。常見的線粒體DNA缺失突變片段有 13162bp,10422bp,7663bp,7436bp,4989bp,4977bp,其中人mtDNA4977bp(對應大鼠4834bp)是mtDNA中最為常見突變類型[3]。2015年10月至2016年3月作者通過實驗研究,探討老年性聾發生的可能機制。
1 材料與方法
1.1 實驗動物 SPF級SD大鼠20只,其中24個月齡SD大鼠10只,平均(647.83±10.35)g;3個月齡SD大鼠10只,平均(210.78±5.62)g,所有動物均無噪聲暴露史及其它藥物使用史,且無中耳炎,動物由華中科技大學同濟醫學院提供。
1.2 方法 (1)分組:20只SD大鼠經聽性腦干反應(ABR)檢測,排除中耳炎后按年齡分2組,各10只,24個月齡為觀察組;3個月齡為對照組。(2)檢測大鼠聽性腦干反應(ABR)反應閾:分別對對照組和觀察組大鼠進行ABR反應閾測定(測定方法參照孔維佳建立方法[4])。測試前兩組大鼠用10%水合氯醛按照0.4ml/kg腹腔內注射麻醉,使用GSI Audera腦干誘發電位儀測試。(3)蝸神經核及內耳組織mtDNA提取:兩組動物ABR測試結束后立即麻醉斷頭處死,首先提取腦蝸神經核:于中線處剪開顱骨,將腦組織輕輕從顱骨中取出,根據蝸神經核[5]定位提取腦蝸神經核,置入勻漿器中,加入2ml勻漿液(pH7.4,0.01mol/L Tris-HCI,0.1mmol/L,0.01mol/L蔗糖)勻漿至無明顯組織塊存在(冰浴操作)。勻漿液4℃離心1000g l0min,取上清液0℃離心12000g 15min,棄上清液,沉淀物即為腦組織蝸神經核線粒體。然后提取內耳組織:提取雙側聽泡,置于4℃預冷的充氧無鈣鎂細胞外液中(0.1mol/L PBS,pH 7.4),體視顯微鏡下剝去骨性蝸殼,解剖出耳蝸Corti器,血管紋,螺旋神經節等內耳組織。蝸神經核和內耳組織線粒體DNA提取方法:各組織置入勻漿管,加入RIPA裂解液,再加入蛋白酶 K,至終濃度10mg/ml,搖勻混合,55℃水浴過夜消化。最后用酚-氯仿-異戊醇抽提取線粒體DNA,所得 DNA 溶于 50μl TE 溶液,-20℃保存備用。取2μl DNA樣本溶液稀釋至50μl,用紫外線核酸檢測儀器檢測A260/A280純度值并測定其濃度大小。(4)線粒體DNA缺失檢測分析:根據大鼠線粒體基因組全序列(Genbank序列號X 14848),并遵循引物設計原則,引物由上海生工生物工程公司合成:P1:5'-GCGAAGCTTAGAGCGTTAAC-3',P2:5'-AGT GAG ATA AGG AAG CCT GC-3',引物合成后用滅菌超純水稀釋成10μmol/L 的工作液,-20℃凍存。配制25μl PCR反應體系:Taq DNA 聚合酶(2.5U)0.5μl,引物(10μM) 各 1μl,dNTPs 2μl,PCR buffer2.5μl。PCR擴增反應條件為:95℃預變性5min,94℃變性30s,56℃退火30s,72℃延伸45s,反應35個循環;末次72℃延伸7min。引物P1、P2如可以擴增597bp片段,則提示樣本中有mtDNA4834bp缺失。如樣本中無4834bp缺失突變,應在現實反應條件下無法合成近5000bp片段,故無法擴增出597bp PCR產物。(5)PCR產物鑒定與分析:各組PCR擴增產物經1.0%瓊脂糖凝膠電泳,溴化乙啶染色,電泳緩沖液1 X TAE,電壓100V,用凝膠成像儀掃描成像并保存結果。在紫外燈下確定597bp目的基因條帶位置,將凝膠與產物一起切出,用DNA凝膠回收試劑盒(AXYGEN APGX-50G)分離純化。加樣后將離心管置37℃水浴1h,酶切產物經2. 5%瓊脂糖凝膠電泳,溴化乙啶染色,凝膠成像儀掃描成像并保存結果。
1.5 統計學分析 采用SPSS18.0統計軟件。計量資料以()表示,用t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 ABR反應閾 對照組大鼠ABR平均聽閾(19.50±3.69)dB, 觀 察 組 大 鼠 ABR平 均 聽 閾(46.00±3.94)dB,兩組差異有統計學意義(P<0.05)。
2.2 mtDNA4834bp缺失突變電泳結果 對照組大鼠腦組織蝸神經核樣本(10例)及內耳組織樣本(10只)均未檢測出597bp(提示4834bp缺失)擴增條帶(見圖1),觀察組大鼠蝸神經核樣本檢出率為100%;內耳組織樣本檢出率90.0%。見圖2。
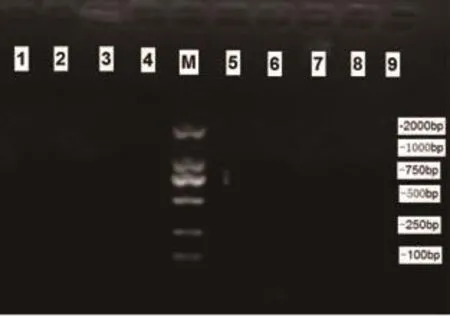
圖1 對照組各組織標本PCR結果
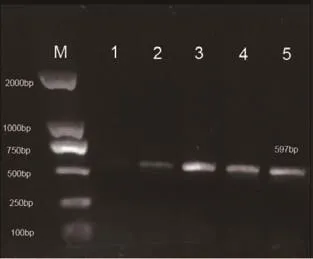
圖2-1 觀察組蝸神經核組織mtDNA PCR結果
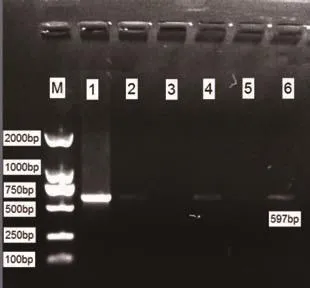
圖2-2 觀察組內耳組織mtDNA PCR結果
2.3 PCR酶切電泳 根據大鼠線粒體基因組全序列(Genbank序列號X 14848),根據DNASTAR軟件選擇限制性內切酶(HphⅠ)酶切,酶切位點見圖3,結果證實酶切產物來自mtDNA4834bp特異型缺失擴增片段,見圖4。

圖3 HphⅠ限制性內切酶酶切位點
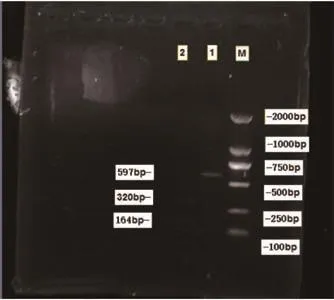
圖4 PCR產物酶切電泳圖
3 討論
老年性耳聾發生機制目前尚不明確,大多學者認為其由多種危險因子的長期累積共同作用形成,其中最主要因素為衰老。根據Harman[6]提出衰老的自由基學說,認為衰老是由自由基引起的組織隨機毒害的結果,自由基損害作用的機制主要涉及DNA的損傷,尤其是mtDNA的氧化損傷,其持續產生于線粒體電子傳遞鏈的活性氧簇(ROS)是衰老過程中線粒體DNA氧化損傷產生的主要原因。由于mtDNA裸露于產生高氧自由基的線粒體內膜下,本身無組蛋白的保護又缺乏損傷修復系統,其突變率較核DNA高10~20倍[7],極易受氧自由基的侵襲發生氧化修飾和mtDNA突變,由缺陷的mtDNA編碼的呼吸酶影響電子轉運功能,增加電子泄露,不斷產生的ROS又反過來繼續氧化損害線粒體功能,導致不同組織細胞程度不一的mtDNA累積突變的蓄積,即“老化的線粒體理論[8]”。衰老過程中產生的mtDNA缺失突變型mtDNA比完整的野生型mtDNA長度小[9],復制速度快,其增殖優勢導致缺失突變型mtDNA分子比例隨年齡增長不斷累積增加。當細胞內突變mtDNA積累達到氧化磷酸化(OXPHOS)所生成的能量低于維持正常細胞功能閾值時,組織器官開始退化,出現臨床癥狀,引起老年退化性疾病,在聽覺器官表現為聽力逐漸下降。本實驗中對照組大鼠ABR平均聽閾為(19.50±3.69)dB peSPL,觀察組大鼠ABR平均聽閾(46.00±3.94)dB peSPL,兩組間差異有統計學意義(P<0.05)。
老年性聾是機體衰老過程在聽覺器官的表現。研究發現線粒體DNA突變與老年性耳聾的發生關系密切。人類mt DNA 在8483~13459bp之間存在著13bp的重復序列,在氧化應激刺激下,易發生斷裂,在修復時由于兩端重復序列,產生DNA兩條鏈下游環,該堿基環富含鳥嘌呤堿基,對自由基及其敏感,一旦受到ROS攻擊,該環降解即容易導致“誤認”發生了4977bp片段缺失[10],此機制被稱為線粒體重組滑行錯配學說。此缺失在大鼠,mtDNA8103-12937bp之間存在16bp直接重復序列,故大鼠也存在類似缺失突變(mtDNA4834bp)的常見缺失(CD)。突變不僅存在衰老組織中(心肌、肝、腦、肺、腎等),還可存在于線粒體腦肌病、進行性眼外肌麻痹和Kearns-Sayre綜合征等疾病中,且隨著年齡增加在組織中逐漸積累[11]。本實驗PCR擴增出597bp片段(提示mtDNA4834bp缺失),該缺失突變編碼包括ND3,ND4,NDS,COIII,ATPaseB,ATPase6 等[12]基 因,影響呼吸鏈有關亞單位蛋白的合成,從而形成有缺陷的呼吸鏈,ATP產生不足導致蝸內離子失平衡,內環境紊亂,最終導致內耳毛細胞非特異性損害,引起聽力下降[13]。本實驗在大鼠蝸神經核和內耳組織中均發現了mt4834bp缺失,表明該常見缺失普遍存在于衰老組織。因此以后可以通過減少和預防mtDNA缺失突變的發生,以緩解和預防老年性耳聾發生發展,從而為治療老年性耳聾提供新突破。但老年性聾不一定均有mtDNA4834bp片段缺失,本實驗中觀察組有1只未檢測到該缺失突變,表明除了基因背景以外,還有其他環境易感因素(如藥物,噪音等)在老年性聾的發病過程中起重要作用,具體機制有待于進一步的研究。
