轉多抗基因新疆大葉苜蓿光合生理特征對土壤水分變化的響應
2018-11-19 08:46:16李敏蘇國霞熊沛楓康繼月郭尚洙王智徐炳成
草業學報 2018年11期
李敏,蘇國霞,熊沛楓,康繼月,郭尚洙,王智,*,徐炳成,*
(1.西北農林科技大學黃土高原土壤侵蝕與旱地農業國家重點實驗室,陜西 楊凌 712100;2.中國科學院水利部水土保持研究所,陜西 楊凌 712100;3.韓國生命工學研究院植物系統工程研究中心,韓國 大田 305-806)
紫花苜蓿(Medicagosativa)是一種優良的多年生豆科牧草,具有營養價值高、適應能力強、產量高等特點,有“牧草之王”的美稱,在全球范圍內被廣泛種植,也是我國栽種面積最大的牧草種之一[1]。紫花苜蓿生物固氮和較高的生長速率能夠顯著改善土壤性狀和提高地表蓋度,是一種優良的水土保持植物[2]。但苜蓿屬于高耗水作物,水分是影響其分布和產量的主要因子。在我國北方旱區,降水量少且年季分布不均,培育具有較強抗旱性的苜蓿品種,對解決因水分虧缺造成的苜蓿品質差、產量低問題,以及促進苜蓿產業的持續發展具有重要意義。
在優良苜蓿品種培育過程中,采用雜交、回交、自然選擇以及接種真菌等傳統方法可以對苜蓿抗旱性進行改良[3]。相比傳統育種方法,轉基因技術具有周期短、效率高、成本低且可以定向改良遺傳性狀等特點[4]。自1986年Deak等[5]首次培育出轉pGA471基因苜蓿以來,采用轉基因技術已培育出了許多抗逆性強的苜蓿品種,如通過對編碼滲透調節物質合成基因[6]、功能性蛋白和有機分子合成酶基因[7]、信號分子與轉錄因子基因[8]、活性氧清除分子基因[9]等的克隆以及在植物體內的表達,均能夠提高苜蓿的抗逆性。
膽堿氧化酶基因(codA)的表達產物甜菜堿是一種重要的滲透調節物質,能夠有效地穩定生物大分子的結構和功能,維持酶活性以及膜的高度有序性,從而提高植物的耐逆性[10]。NDPK2基因是與核苷二磷酸激酶(NDPKs)合成密切相關的基因之一,該酶可維持細胞二磷酸核苷(NDP)和三磷酸核苷(NTP)的代謝平衡,與細胞的生長和分化、信號轉導密切相關,并參與干旱、熱脅迫等響應過程[11-12]。ABF3是編碼ABF/AREB轉錄因子的基因,ABF/AREB轉錄因子參與脫落酸(abscisicacid,ABA)或脅迫誘導的信號傳遞過程進而提高植物對環境脅迫的耐受性[13]。Orange基因(Or)能夠通過誘導無色質體分化為有色體的代謝途徑來促進類胡蘿卜素積累,類胡蘿卜素為一種光合色素和非酶促抗氧化系統的重要組成成分,在植物的抗逆生理中發揮重要作用[14]。
目前,已對上述4種基因的轉基因新疆大葉株系在干旱等脅迫下的生理生化特性進行了研究,但有關光合生理的變化特征還未涉及[15-18]。光合作用作為植物生長發育的基礎,能直接反映外界環境因子對植物生理過程的影響。植物對外界環境特征的響應可以通過光合生理特征予以展現,并可通過葉綠素熒光參數的變化來反映[19]。因此,本研究通過設置不同水分條件,模擬土壤水分降低過程中干旱加劇,對比研究了4種轉多抗基因株系及其母本新疆大葉苜蓿的光合及熒光參數特征的響應過程和差異,以期明確4種多抗基因的表達對苜蓿抗性生理狀況的影響,為苜蓿抗旱新品種的培育提供依據和參考。
1 材料與方法
1.1 試驗材料
以新疆大葉苜蓿(NT)及其為母本的4種轉基因株系[SN:轉擬南芥NDPK2基因(AtNDPK2);SC:轉球形節桿菌codA基因(codA);SOR:轉甘薯IbOr基因(IbOr);SAF:轉擬南芥ABF3基因(AtABF3)]為研究材料。轉基因植株由韓國生命工學研究院提供。試驗土壤為陜北天然草地耕層(0~30 cm)黃綿土,土壤養分含量分別為:全氮0.28 g·kg-1,全磷0.67 g·kg-1,有機質19.90 g·kg-1。土壤pH值8.21,田間最大持水量(field capacity, FC)為20%,萎蔫系數(wilting point)為4%。
1.2 試驗處理
采用盆栽控制試驗,規格為10 cm×56 cm(內徑×高度)底部封堵的PVC管,每桶裝4.5 kg干土,裝桶時桶底鋪碎石子,沿內壁放一根內徑為2 cm的PVC管作為灌水管。
2016年3月中旬,將培養于1/2 MS培養基的各轉基因苜蓿幼苗轉移到基質[有機質含量>20%、N-P2O5-K2O總養分>1.5%(富含多種礦物質成分)、通氣孔隙度>20%、pH 8.12]中于培養箱(光照時間6:00~20:00,溫度25 ℃)內進行擴繁,待其生長至分枝期(4月末)后剪取長勢一致的枝條在1/4 Hoagland(pH 6.5~7.0)溶液中水培生根,溫室光照時間14 h(6:00~20:00),晝溫25~28 ℃,夜溫18~20 ℃。培育2周后,當枝條新生根達到10 cm左右,挑選地上和地下部分長勢一致的枝條移栽于試驗桶中在室外防雨棚下進行土培,每桶1株。苗期充分供水(土壤含水量保持80% FC左右),并在各桶覆蓋2 cm珍珠巖,以減少土壤表面水分蒸發。
當植株處于始花期時開始水分處理(8月1日),土壤含水量由80% FC逐漸降至70% FC后維持3 d,隨機選取3株進行指標測定(8月4日),繼續降至60% FC、50% FC、40% FC后分別于各土壤含水量水平下控水3 d后(8月7日、8月10日、8月13日)測定各項指標(圖1),并保留3桶植株至試驗結束用于收取生物量。處理總數=5(株系)×18(重復)=90(桶)。土壤含水量采用稱重法控制,控水于每日18:00進行。

圖1 土壤水分處理示意圖Fig.1 Diagram of soil water treatment
1.3 測定項目與方法
1.3.1光合氣體交換參數 采用Li-6400便攜式光合儀(LI-COR Inc., Lincoln, NE, USA)測定。在測定當日9:00~11:30選取植株第3莖節充分展開葉進行測定,重復3次。測定時光強設定為1200 μmol·m-2·s-1。項目包括:凈光合速率(Pn, μmol·m-2·s-1),蒸騰速率(Tr, mmol·m-2·s-1),氣孔導度(Gs, mmol·m-2·s-1),葉片瞬時水分利用效率(WUEi, μmol·mmol-1)=Pn/Tr。
1.3.2葉綠素熒光參數 采用Imaging-PAM熒光儀(imaging PAM, WALZ, effeltrich, germany)測定。測定當日清晨,植株經暗適應30 min后,選取植株第3莖節充分展開葉測定,重復3次。項目包括:初始熒光(Fo)、最大光化學效率(Fv/Fm)、實際光化學效率(ΦPSⅡ)、光化學淬滅系數(qP)、非光化學淬滅系數(NPQ)、表觀光合量子傳遞速率(ETR)。
1.3.3生物量 試驗結束后,分別收集每桶植株地上和地下生物量。地上生物量在105 ℃殺青15 min,80 ℃烘干至恒重,最后稱其干重(g)。植株根系經水沖洗后裝入信封袋在80 ℃烘干至恒重稱其干重(g)。總生物量(g)=地上生物量+地下生物量。
1.4 數據分析
采用SPSS 21.0軟件對試驗數據進行統計分析,用平均值和標準誤表示測定結果,分別對同一水分條件下不同株系間以及同一株系不同水分處理下進行單因素方差分析,用LSD進行顯著性檢驗(P≤0.05),采用SigmaPlot 10.0進行繪圖。
2 結果與分析
2.1 轉多抗基因新疆大葉苜蓿的光合氣體交換參數
在80% FC時,SAF和SOR的Pn值顯著高于NT、SC和SN(P<0.05),SN的Pn值顯著最低(P<0.05)。當土壤水分降低至70% FC時,各轉基因株系的Pn值間無顯著差異但均顯著大于NT(P<0.05),在60% FC時,株系間Pn值無顯著差異,50% FC時,SC與SOR無顯著差異,但均顯著大于其他各株系(P<0.05)。當土壤水分為40% FC左右時,SN、SAF和SOR的Pn相對80% FC分別顯著下降了61.0%、75.6%和69.4%(P<0.05),而NT(6.06 μmol·m-2·s-1)降低幅度最大,顯著降低了84.3%(P<0.05)。此時SC的Pn值為21.5 μmol·m-2·s-1,顯著下降了43.8%(P<0.05),但仍顯著高于其余各株系(P<0.05)(圖2A)。

圖2 不同土壤含水量條件下各株系凈光合速率、蒸騰速率、水分利用效率及氣孔導度變化Fig.2 The leaf net photosynthesis rate (Pn), transpiration rate (Tr), water use efficiency (WUEi) and stomatal conductance (Gs) of all alfalfa lines under each soil water content NT:新疆大葉苜蓿Xinjiang Daye alfalfa;SN:轉AtNDPK2基因株系Transgenic AtNDPK2 lines;SC:轉codA基因株系Transgenic codA lines;SAF:轉AtABF3基因株系Transgenic AtABF3 lines;SOR:轉IbOr基因株系Transgenic IbOr lines.不同小寫字母表示同一水分處理下株系間差異顯著(P<0.05),括號內不同小寫字母表示同一株系水分處理間差異顯著(P<0.05),下同。Different small letters above the bars indicate significant differences between alfalfa lines (P<0.05); different small letters in bracket show significant differences among soil water contents for each alfalfa line (P<0.05). The same below.
在80% FC時,NT和SC的Tr值顯著高于SN、SAF和SOR(P<0.05),SN的Tr值顯著最低(P<0.05)。隨著土壤水分下降,SC和SOR的Tr值逐漸下降,于40% FC時降至最低,分別顯著下降64.2%和74.3%(P<0.05);NT和SAF的Tr值隨土壤水分降低呈先下降后上升再下降的趨勢,土壤水分降至40% FC時,NT、SN和SAF分別顯著下降82.3%、56.4%和82.8%(P<0.05),此時SC的Tr值最大且顯著大于其他株系(P<0.05),其他株系間差異不顯著(圖2B)。
在土壤水分下降過程中,各株系的葉片WUEi值總體呈升高趨勢。80% FC時,SN的WUEi顯著高于其他株系(P<0.05),SAF與SOR無顯著差異,但均顯著大于NT和SC。70% FC時,SC和SAF的WUEi顯著低于其他株系(P<0.05)。土壤水分由60%降至40% FC過程中,各株系的WUEi無顯著差異,在40% FC時以SAF的WUEi最大(圖2C)。
在80% FC時,NT和SC的Gs值顯著最高,SAF與SOR無顯著差異但顯著小于SC和NT,以SN顯著最低(P<0.05)。在土壤水分下降過程中,SAF的Gs值一直呈降低的趨勢,在40% FC時降至最低值,相對80% FC顯著下降了89.0%。SN的Gs值呈先上升后下降的趨勢,在70% FC時升至最高值后下降,在40% FC時下降至最低,相對80% FC顯著下降77.4%(P<0.05);NT和SC的Gs值呈先下降后上升再下降的趨勢,在40% FC時,NT和SC的Gs值分別顯著下降了87.5%和74.4%(P<0.05),SC的Gs值顯著高于其他株系(P<0.05),其他株系間無顯著差異(圖2D)。
2.2 轉多抗基因新疆大葉苜蓿的葉綠素熒光參數
2.2.1初始熒光(Fo) 80% FC時,各株系間的Fo值無顯著差異。與80% FC相比,在土壤水分降至40% FC時,SN的降幅最大,顯著降低了34.3%(P<0.05),SAF的降幅最小為8.1%。SC的Fo值在80%和70% FC維持較高水平,而后顯著下降。在40% FC時,以SAF、SC和SOR的Fo值最大,但相互間無顯著差異。在水分降低過程中,NT的Fo值呈先降后升趨勢,40% FC時僅比80% FC低0.3%(表1)。

表1 不同土壤含水量條件下各株系苜蓿葉片初始熒光(Fo)值Table 1 Leaf initial fluorescence (Fo) values of all alfalfa lines under each soil water content
注:同一列數字后不同小寫字母表示相同水分條件下品系間差異顯著(P<0.05),括號內不同小寫字母表示同一株系水分處理間差異顯著(P<0.05),下同。
Note: Values within a column followed by different small letters indicate significant differences between alfalfa lines under same soil water content (P<0.05). Different small letters within a row in bracket show significant differences among soil water contents for each alfalfa line (P<0.05). The same below.
2.2.2最大光化學效率(Fv/Fm) 80% FC時,SAF的Fv/Fm值顯著低于NT和SN(P<0.05),除SAF外其余4株系間無顯著差異(P<0.05)。在土壤水分下降至40% FC時,NT和SOR的Fv/Fm值降至最低值,且NT的Fv/Fm值顯著低于其他各株系(P<0.05),各轉基因株系中,以SAF最低,顯著低于SN和SC(P<0.05)(表2)。
2.2.3實際光化學效率(ΦPSⅡ) 80% FC下,NT的ΦPSⅡ值顯著最低(P<0.05),4種轉基因株系的ΦPSⅡ值間差異不顯著,以SOR最高。各株系的ΦPSⅡ值在土壤水分為70% FC時達到最高值后開始下降,在40% FC時降至最低值。在土壤水分為40% FC時,與80% FC相比,NT、SN、SC、SAF和SOR分別顯著下降85.7%、76.0%、78.8%、68.0%和62.6%(P<0.05),SOR的ΦPSⅡ值顯著高于SC、SN和NT(P<0.05),SOR和SAF間差異不顯著,NT的ΦPSⅡ值最小且顯著小于SOR和SAF(表3)。
2.2.4光化學淬滅(qP)和非光化學淬滅(NPQ) 80% FC下,各株系的qP值間無顯著差異,以NT最低。各株系qP值在土壤水分為70% FC時上升至最高后開始下降,在40% FC時降到最低,與80% FC相比,NT、SN、SC、SAF和SOR的qP值分別顯著下降78.0%、65.9%、69.2%、55.8%和51.1%(P<0.05),SAF的qP值顯著高于SC和NT,與SOR間無顯著差異,且SC和NT二者差異不顯著,以NT的qP值最低(表4)。

表2 不同土壤含水量條件下各株系苜蓿葉片最大光化學效率(Fv/Fm)值Table 2 Leaf maximum photochemical efficiency values of PSⅡ (Fv/Fm) values of all alfalfa lines under each soil water content

表3 不同土壤含水量條件下各株系苜蓿葉片實際光化學效率(ΦPSⅡ)值Table 3 Leaf active photochemical efficiency values of PSⅡ (ΦPSⅡ) values of all alfalfa lines under each soil water content

表4 不同土壤含水量條件下各株系苜蓿葉片光化學淬滅(qP)值Table 4 Leaf photochemical quenching coefficient of PSⅡ (qP) values of all alfalfa lines under each soil water content
80% FC時,各株系的NPQ值無顯著差異。土壤水分下降過程中,SAF和SOR的NPQ值呈逐漸上升趨勢,在土壤含水量為40% FC時升至最高值。其他各株系的NPQ值隨土壤水分下降總體呈先升后降趨勢,NT和SN于50% FC時升至最高值后下降,SC的則為60% FC時升至最高值后下降。40% FC時,NT、SN、SC、SAF和SOR的NPQ值相比80% FC分別顯著升高489.4%、606.5%、647.9%、491.8%和307.0%(P<0.05),SN的NPQ值顯著高于NT、SOR和SAF(P<0.05)(表5)。
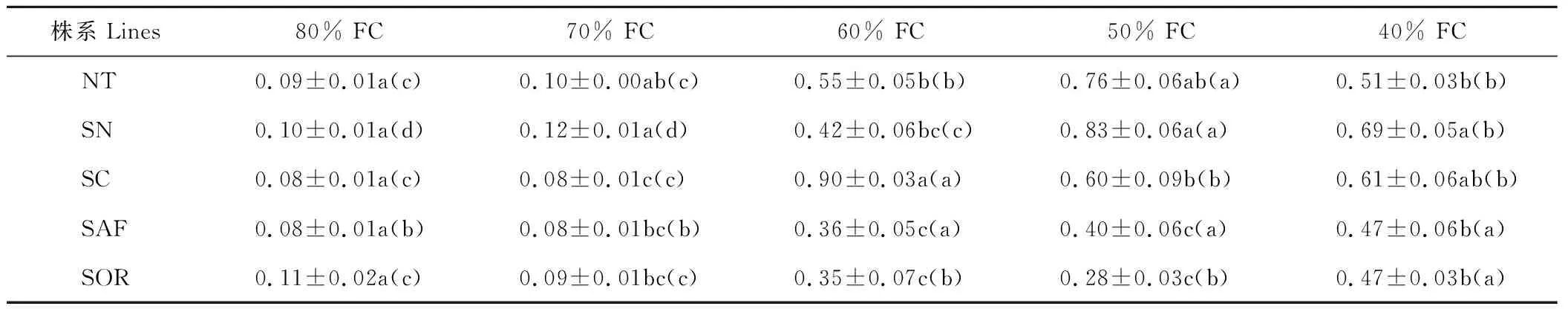
表5 不同土壤含水量條件下各株系苜蓿葉片非光化學淬滅(NPQ)值Table 5 Leaf non-photochemical quenching coefficient of PSⅡ (NPQ) values of all alfalfa lines under each soil water content
2.2.5各株系表觀光合量子傳遞效率(ETR) 80% FC時,SOR的ETR值顯著高于NT(P<0.05),且4種轉基因苜蓿間差異不顯著。各株系的ETR值在土壤水分減少時呈先上升后下降趨勢。SC和SOR在土壤含水量為50% FC達到最大值,其他株系在60% FC時均升至最大而后下降。在40% FC時,NT的ETR值提高了121.6%,而各轉基因株系SN、SC、SAF和SOR的ETR值分別顯著升高271.0%、229.0%、395.5%和479.1%(P<0.05)(表6)。

表6 不同土壤含水量條件下各株系苜蓿葉片表觀光合量子傳遞效率(ETR)值Table 6 Apparent electron transport rate values of PSⅡ (ETR) values of all alfalfa lines under each soil water content
2.3 轉多抗基因新疆大葉苜蓿的生物量
試驗結束時,SN的地上生物量顯著大于NT,轉基因株系SC和SOR與NT間無顯著差異,而SAF則顯著小于NT。各株系間地下生物量均無顯著差異;總生物量以SN最大,且顯著大于NT(P<0.05),4種轉基因苜蓿株系的總生物量無顯著差異。SC、SN、SOR和SAF苜蓿的總生物量較NT分別提高了12.3%、36.5%、29.4%和13.8%(圖3)。

圖3 各株系地上生物量、地下生物量和總生物量Fig.3 The shoot biomass, root biomass and total biomass production of each alfalfa line
3 討論
水分是影響植物光合生理特性的重要因子[20]。水分脅迫會引起植物氣孔關閉,降低葉綠體活性,影響電子傳遞、光合磷酸化及暗反應等一系列過程,使植物的光合速率降低[21]。光合作用是受水分脅迫影響最為明顯的生理過程之一,葉綠素熒光參數更具有反映氣體交換“內在性”特點,所以常用于評價環境脅迫對植物葉片光合器官結構與功能的影響[22]。葉綠素熒光參數中的初始熒光(Fo)值是PSⅡ反應中心處于完全開放時的熒光產量,其減小表明天線色素熱耗散增加,增大表明PSⅡ反應中心遭受嚴重損傷,而最大光化學效率(Fv/Fm)的高低與植物光合速率的高低密切相關[23]。本研究中,當土壤水分從80% FC降至70% FC時,非轉基因、轉AtABF3和轉IbOr基因株系的Pn、Gs值顯著下降,但三者的Fv/Fm并未發生顯著變化,說明此階段光系統并未受到損傷,且Pn下降可能是由于Gs下降造成胞內CO2供應不足即氣孔限制引起的[24]。當土壤水分從60% FC降至40% FC時,各株系的Pn和Gs值均顯著下降,但轉基因株系的Fv/Fm值均未發生顯著變化,僅非轉基因株系的Fv/Fm值顯著下降,且從50% FC降至40% FC時,其Fo值顯著上升,說明此階段NT的Pn值下降不僅受氣孔限制影響,其光系統也受到嚴重損傷,而各轉基因株系Pn值下降的原因仍然是氣孔限制[25]。
在土壤水分下降過程中,以轉codA基因株系Pn值最高(圖2),尤其是土壤水分為50% FC和40% FC時,除與轉IbOr基因株系無差異外,均顯著大于其他各株系,這可能與codA基因參與滲透物質甜菜堿的合成而使植株維持較高的滲透壓有關。研究表明,轉codA基因新疆大葉株系的甜菜堿、脯氨酸和葉片相對含水量在干旱脅迫下均能維持較高水平且顯著大于非轉基因植株[16],Sakamoto等[26]對轉codA基因水稻(Oryzasativa)的研究指出,葉肉細胞中甜菜堿含量的增加對轉基因植株的PSⅡ反應中心可提供更有效的保護。植物PSⅡ反應中心的活性可用表觀光合量子傳遞效率(ETR)來反映,它是表征光合能力大小的有效參數[27]。本研究中轉codA基因株系ETR值在50% FC降至40% FC時顯著下降,而其他株系的ETR值出現顯著下降時的土壤含水量均要高于此(表6),說明轉codA基因株系PSⅡ反應中心活性受損程度較其他株系要小[28]。因此轉codA基因株系具有高光合速率的原因可能是codA基因過表達使植物體內甜菜堿含量增多,從而維持細胞在干旱脅迫下的滲透壓、提高PSⅡ反應中心抗性,保持其轉基因株系在干旱脅迫下的高光合效率。
植物的實際光化學效率(ΦPSⅡ)是評價光合電子傳遞速率快慢的指標,也可反映植物PSⅡ反應中心活性[29]。本研究中,各株系葉片ΦPSⅡ和ETR值隨土壤水分下降均呈先升高后降低的趨勢,但二者發生顯著降低時所對應的土壤水分存在差異(表3和表6)。ΦPSⅡ值在土壤水分從70% FC降至60% FC各株系均顯著下降,而ETR值在60% FC降至50% FC以及50% FC到40% FC才顯著下降,說明ETR值變化要滯后于ΦPSⅡ值,這可能是因為植物為了有效適應干旱脅迫,將光反應中心大量電子分配到光化學反應中[30]。當土壤水分為50% FC和40% FC時,轉基因株系的ΦPSⅡ和ETR值均顯著高于非轉基因,說明轉基因株系的光系統對干旱脅迫具有更強的耐受性[31]。在各轉基因株系中,以轉IbOr基因的ΦPSⅡ值和ETR值顯著最高,說明轉IbOr基因株系能在干旱脅迫下PSⅡ反應中心維持較高的電子傳遞效率[27],這是其具有較高Pn值的重要原因,同時轉IbOr基因株系的非光化學淬滅系數(NPQ)的變化也可對此做出進一步說明。NPQ是衡量過剩激發能耗散的重要指標,植物可通過增大NPQ將過剩光能耗散以保護光合器官不受損傷來適應外界環境變化。在土壤水分下降過程中,轉IbOr基因株系的NPQ值變化幅度最小,且在40% FC時仍能顯著上升(表5),說明其能通過增加熱耗散對光系統起到了很好的保護作用[32],這是其體內類胡蘿卜素含量上升所導致的。類胡蘿卜素在淬滅過剩光能使光系統在脅迫下免遭嚴重損傷中起重要作用[33]。IbOr基因能夠通過誘導無色質體分化為有色體的代謝途徑來促進類胡蘿卜素積累[34],對轉IbOr基因新疆大葉株系的研究表明,其總類胡蘿卜素及其組分葉黃素、紫黃質的含量較非轉基因株系均顯著升高,這是轉IbOr基因株系具有較高光合速率的物質基礎[14]。
在干旱環境下,植物會通過調節碳同化和水分消耗之間的關系來提高葉片水分利用效率(WUEi)[35],本研究中,各株系Pn下降的同時Tr下降程度更大,說明植株可通過減小蒸騰速率來提高水分利用效率,這是植物的重要抗旱策略之一[36]。在土壤水分為40% FC時,以轉AtABF3株系的WUEi最高(圖2),這可能是由于AtABF3基因與脫落酸(ABA)的信號傳遞過程相關。ABA對氣孔開啟具有明顯的抑制作用,它在植物受到脅迫時可迅速合成并傳遞至地上部分引起氣孔關閉降低蒸騰速率而提高水分利用效率,以更好地應對干旱脅迫[37]。但ABA也可促進子葉脫落、抑制植物細胞生長[38],轉AtABF3基因新疆大葉株系的研究表明,轉基因株系的葉面積較非轉基因顯著減小了30%以上[14],這是轉AtABF3株系的地上生物量顯著最低的重要原因之一。地上生物量和總生物量僅有轉AtNDPK2株系的顯著大于非轉基因株系(圖3),這與AtNDPK2基因的功能密不可分。Choi等[39]對擬南芥(Arabidopsisthaliana)ndpk2缺失突變體研究生長發育與NDPK2的關系,發現NDPK2參與生長素相關的子葉發育、生長素極性運輸等,以促進植物生長發育;也有研究表明AtNDPK2基因的過表達會促使吲哚乙酸分泌,可促進細胞分裂分化和促使植物生長,這是轉AtNDPK2株系地上生物量和總生物量顯著大于非轉基因的原因之一[40]。
綜上所述,在土壤水分下降過程中,轉基因株系在光合生理中表現出了優于非轉基因的特性,其中轉codA基因株系可維持較高的凈光合速率,轉AtABF3基因株系能夠高效利用水分,轉IbOr基因株系能夠維持較高的光能利用能力,轉AtNDPK2基因株系的地上生物量顯著提升。本研究主要是在始花期開展的短期初步探索,轉基因能否對苜蓿整個生育期的光合及最終產量產生影響,還需進一步就各株系在不同生育期的光合生理及其根冠生長進行更深入系統的研究,為高產優質耐旱苜蓿新品種的培育奠定基礎。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
