NEDDylation參與乙型肝炎病毒前基因組RNA的包裝
2018-11-14 11:34:22李程張境王勇翔
中華實驗和臨床病毒學雜志 2018年5期
關鍵詞:檢測
李程 張境 王勇翔
200032上海,復旦大學基礎醫學院病原生物學系醫學分子病毒學教育部/衛生部重點實驗室
乙型肝炎病毒(hepatitis B virus,HBV)感染是世界范圍的健康問題。據估計全球約有2.5億人罹患慢性乙型肝炎[1]。目前治療慢性乙肝的抗病毒藥物有干擾素α、PEG化干擾素α和核(苷)酸類似物等[2]。干擾素治療具有明顯的副作用;核苷酸類似物治療因停藥后出現病毒復制反彈而需長期用藥,有病毒耐藥的風險[3]。因此,迫切需要開發新的抗HBV制劑。
HBV通過結合鈉離子-牛磺膽酸共轉運蛋白(sodium taurocholate co-transporting polypeptide,NTCP)受體進入肝細胞,核衣殼被運輸到細胞核。在細胞核內,核衣殼內的松弛環狀 DNA(relaxed circular DNA,rcDNA)被修復成共價閉合環狀DNA(covalently closed circular,cccDNA)。 以cccDNA為模板,細胞RNA聚合酶II轉錄出5種HBV RNA,包括3.5 Kb編碼HBe抗原前體的precore RNA和編碼核心蛋白和聚合酶的前基因組RNA(pregenomic RNA,pgRNA)、2.4 Kb編碼 L蛋白的 mRNA、2.1 Kb編碼M和S蛋白的mRNA,0.7 Kb的編碼X蛋白的mRNA。HBV聚合酶結合pgRNA5’端莖環結構,同時招募核心蛋白二聚體包裹pgRNA成核衣殼[4]。在核衣殼內,HBV聚合酶以pgRNA為模板逆轉錄合成負鏈 DNA后,通過 RNaseH活性將pgRNA/負鏈DNA雙鏈中的pgRNA降解,再以負鏈DNA為模板合成正鏈DNA。核衣殼在多囊泡體出芽并釋放出細胞[5]。
NEDD8是與泛素類似的蛋白。NEDD8在其活化酶(NEDD8 activating enzyme, NAE/E1)、結合酶(E2),連接酶(E3)的催化下,將其羧基端第76位的甘氨酸與底物的賴氨酸共價結合,這一過程叫做NEDDylation。NEDDylation的底物主要是含 cullin的泛素連接酶(cullin-RING liagses,CRLs)[6],參與調節泛素-蛋白酶體系統(ubiquitin-proteasome system,UPS)從而參與調節細胞信號轉導、細胞周期、細胞生長及凋亡。近年來研究表明NEDDylation還與病毒復制有關。皰疹病毒、腺病毒和流感病毒通過NEDDylation激活CRLs而促進病毒復制[7-8];一些RNA病毒(如仙臺病毒、水皰性口炎病毒等)通過 CRLs相關的 UPS促進干擾素調節蛋白-3(interferon regulatory factor 3,IRF3)降解而干擾抗病毒先天性免疫反應[9-10]。近年來研究還發現存在不依賴CRLs的NEDDylation[11],提示NEDD8可能具有額外的生物學功能。目前關于細胞NEDDylation在HBV復制中的作用仍不清楚。本研究采用NAE特異性抑制劑 MLN4924抑制細胞CRLs和其他一些底物的NEDDylation,檢測HBV復制和HBV pgRNA包裝效率。
1 材料與方法
1.1 材料 pCH-9/3091質粒由德國弗賴堡大學Michael Nassal教授提供,包含D基因型的HBV基因組(GenBank登記號V01460)。為檢測包裝入核衣殼的pgRNA,本研究構建了HBV聚合酶失活變異體表達質粒pCH-9/3091-YMHD:以pCH-9/3091質粒為基礎,將聚合酶催化中心YMDD基序基因突變為YMHD基因而獲得HBV聚合酶失活變異體pCH-9/3091-YMHD,將該質粒轉染Huh-7細胞后,pgRNA可被包裝入核衣殼,但不能合成負鏈DNA,pgRNA也不被降解,因此核衣殼內的HBV核酸為pgRNA。將 HBV核心蛋白基因擴增后克隆入pCDNA6載體獲得pCDNA-core質粒,將該質粒轉染Huh-7細胞后,細胞雖然產生了大量核心顆粒,但用HBV特異的探針作雜交發現這些顆粒不包裝任何HBV特異的核酸,用作對照。Huh-7細胞和四環素調控且穩定表達HBV的HepAD38細胞為本室保存。MLN4924購自 MedChemExpress公司,胎牛血清購自Gibco公司,DMEM培養液和核酸預雜交液購于ThermoFisher Scientific公司,隨機引物DNA標記試劑盒購自Roche公司,α-32P-dGTP購于Perkin-Elmer公司。實驗中所用抗體及來源如下:anti-core多克隆抗體(Dako公司),anti-core單克隆抗體由張繼明教授(復旦大學附屬華山醫院)饋贈,actin單克隆抗體和gpr78多克隆抗體(Proteintech公司),辣根過氧化物酶(horseradish peroxidase,HRP)標記的二抗(Jackson ImmunoResearch公司)。
1.2 方法
1.2.1 細胞培養:Huh-7細胞用含10%胎牛血清、100 U/ml青霉素、100 U/ml鏈霉素的DMEM培養基,于37℃、5%CO2恒溫恒濕培養箱中培養,每1~2 d進行細胞傳代。HepAD38細胞在上述培養基中額外添加200μg/ml G418抗生素,在相同培養條件下生長,不含四環素的培養液培養使pgRNA轉錄而起始HBV復制,每2~3 d傳代。
1.2.2 DNA轉染:計數3.0×105個Huh-7細胞鋪于6孔板中,18~24 h后進行轉染。取250μl opti-MEM,加入質粒 DNA 2.5μg,混勻后加入7.5μl TransIT-LT1轉染試劑,混勻后靜置15 min。將混合物均勻滴加到已更換新鮮培養液的6孔板中。
1.2.3 藥物處理:向HepAD38細胞培養上清中加入梯度濃度的MLN4924,并用不同濃度的HBV聚合酶抑制劑 LAM作對照,每 3 d更換1次含MLN4924或LAM的培養基,加藥后第6天用NP40裂解緩沖液裂解細胞;向轉染了pCH-9/3091-YMHD的Huh-7細胞中添加MLN4924(終濃度為0.1 μmol/L),并添加DMSO作為對照(MLN4924溶于DMSO),每3 d更換1次含MLN4924或DMSO的培養基,加藥后第6天裂解細胞。
1.2.4 細胞內病毒核衣殼檢測:利用非變性瓊脂糖凝膠電泳(native agarose gel electrophoresis,NAGE)分離HBV核衣殼:NP40裂解液裂解細胞,細胞膜、細胞器膜和HBV包膜破裂,核衣殼釋放出來。裂解液經1%的瓊脂糖凝膠分離,衣殼顆粒在電場的作用下向正極遷移。隨后用免疫印跡檢測核衣殼:通過虹吸轉膜裝置將衣殼顆粒轉移至尼龍膜上,5%脫脂奶粉封閉后與anti-core多克隆抗體孵育,TBST漂洗后與 HRP標記的二抗孵育,用化學發光液(chemiluminescence, ECL)檢測。
1.2.5 十二烷基硫酸鈉-聚丙烯酰氨凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)和免疫印跡:細胞樣品用含SDS的樣品緩沖液處理,100℃變性10 min。配制4.5%濃縮膠和12.5%分離膠進行電泳。電泳后將蛋白轉移到PVDF膜上,按照1.1.3所述方法檢測蛋白。
1.2.6 細胞內HBV DNA提取:取一定量細胞裂解上清加入DNase I 37℃消化處理30 min,再加入蛋白酶K 53℃消化釋放病毒基因組,最后經酚/氯仿抽提和DNA沉淀獲得HBV DNA。
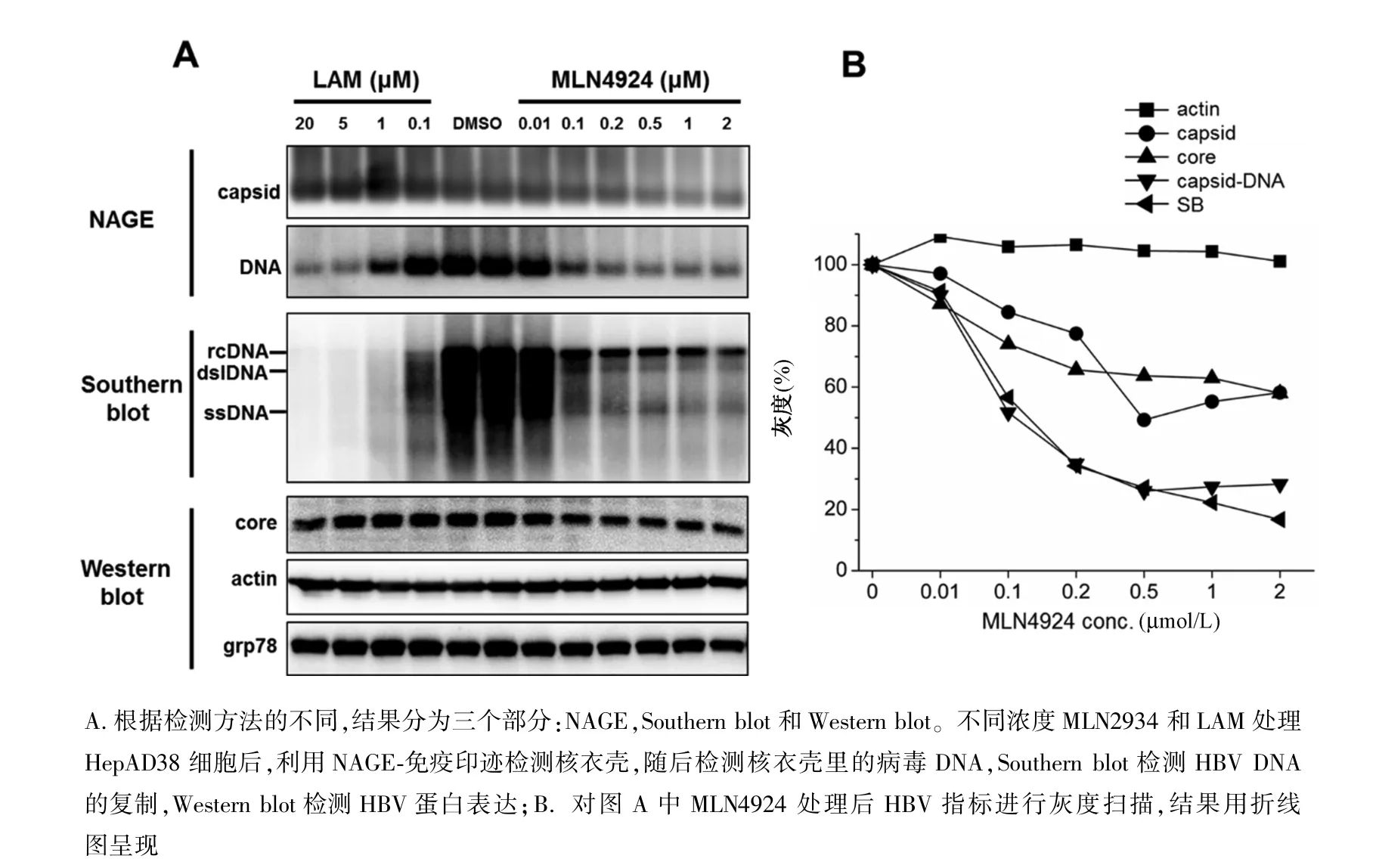
圖1 MLN4924劑量依賴地抑制HepAD38細胞HBV DNA復制A.According to the different detection methods, the results were divided into three parts:NAGE, Southern blot and Western blot.HepAD38 Cells were incubated with various concentrations of MLN4924 and lamivudine for 6 days.Intracellular capsids were analyzed by NAGE, capsid associated viral DNAs by Southern blot, and viral and cellular proteins by Western blot;B.Graphic presentation of HBV replication marker levels at the treatment of increasing concentrations of MLN4924Fig.1 MLN4924 inhibited HBV DNA replication in a dose-dependent manner in HepAD38 cells
1.2.7 Southern blot檢測病毒 DNA復制:HBV DNA樣品經1%的NAGE分離;變性液和中和液分別處理凝膠45 min后,通過虹吸轉膜裝置將病毒DNA轉移至尼龍膜上;尼龍膜80℃烤膜固定;預雜交液42℃孵育30 min;用隨機引物DNA標記試劑盒標記探針;將變性后的摻入32P標記的dGTP的DNA探針加入預雜交液中42℃雜交過夜;洗膜后磷屏曝光,最后用多功能生物分子成像儀掃描磷屏記錄的信號并成像。
1.2.8 原位檢測核衣殼中DNA:1.2.4中經NAGE分離和免疫印跡檢測的核衣殼在80℃烤箱中烘烤2 h,使衣殼顆粒固定在尼龍膜上,DNA變性液處理5 min,中和液處理5 min;再用1.2.6所述方法檢測顆粒內的病毒DNA。
1.2.9 原位檢測核衣殼中RNA:相關溶液均用DEPC水配制,檢測方法同1.2.7。
1.2.10 數據處理:利用 FUJIFILM公司的 Multi Gauge V2.2軟件對蛋白、DNA及RNA信號進行灰度掃描;利用數據處理軟件Origin 8.0進行折線圖的繪制;兩組數據均數的比較采用Student-t檢驗,P<0.05具有統計學意義。
2 結果
2.1 MLN4 924劑量依賴地抑制HBV DNA復制Western blot結果顯示0.1~20μmol/L LAM和0.01~2μmol/L MLN4924對actin和GRP78蛋白的表達無明顯影響(圖1A,Western blot),提示高達20 μmol/L拉米夫定和2μmol/L MLN4924不引起細胞毒性。Western blot檢測細胞內核心蛋白,NAGE-免疫印跡檢測細胞內核衣殼,結果顯示LAM不改變核心蛋白的表達和核衣殼的形成,而MLN4924能輕度劑量依賴地減少核心蛋白和核衣殼(圖1A,Western blot和NAGE)。Southern blot和原位病毒DNA檢測結果顯示LAM和MLN4924都能顯著地且劑量依賴地減少核衣殼相關病毒DNA(圖1,Southern blot和NAGE)。灰度掃描結果顯示,隨著MLN4924濃度增高,actin的量無明顯變化,核心蛋白和核衣殼變化趨勢一致,Southern blot和NAGE原位檢測病毒DNA結果一致(圖1B,Southern blot和NAGE),且隨著MLN4924濃度增加,核衣殼中病毒DNA下降幅度顯著高于核衣殼(圖1A,capsid和DNA;圖1B),這提示HBV pgRNA包裝或下游DNA合成可能被抑制。
2.2 MLN4924抑制HBV pgRNA包裝 pCDNA-core轉染Huh-7細胞后,細胞產生了大量核心顆粒(圖2A,capsid),但HBV特異的探針作雜交結果顯示顆粒中不包裝任何HBV特異的核酸(圖2A,pgRNA)。pCH-9/3091-YMHD轉染Huh-7細胞后,細胞也產生了大量的核心顆粒(圖2A,capsid),而且這些顆粒包裝了pgRNA(圖2A,pgRNA)。當用核心顆粒的量作校正后,發現0.1μmol/L MLN4924使核衣殼包裝的pgRNA減少至DMSO對照組的53.4%(圖2B),而相同濃度的MLN4924也使衣殼相關病毒 DNA減少至對照組的56.6%(圖1B)。因此MLN4924可能通過抑制pgRNA包裝而減弱HBV復制。
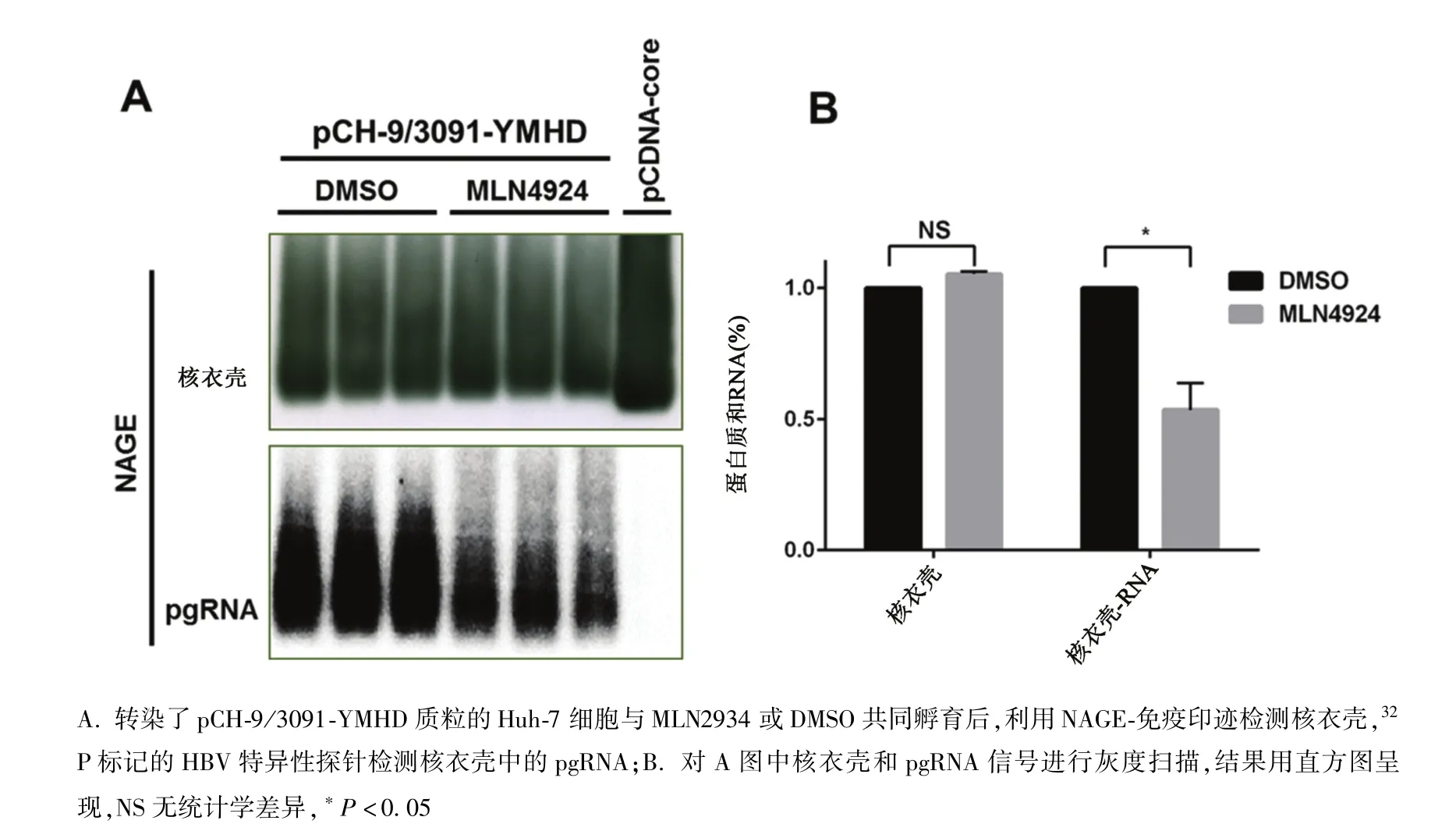
圖2 MLN4924抑制HBV pgRNA的包裝A.Huh-7 cells were respectively transfected with pCH-9/3091-YMHD or pCDNA-core plasmids,then incubated with MLN4924 or vehicle-DMSO.Cytoplasmic capsids and capsid-associated RNA were sequentially detected with NAGE-immunoblotting and hybridization with 32 P labeled HBV specific DNA probes;B.Graphic presentation shows that relative pgRNA encapsidation efficiency by normalizing the amount of packaged pgRNA with that of capsids.NS:Not significant, ?P <0.05 Fig.2 MLN4924 inhibited pgRNA encapsidation
3 討論
本研究采用NAE抑制劑-MLN4924干擾細胞內蛋白NEDDylation修飾反應。MLN4924輕度劑量依賴地抑制核心蛋白的表達。有研究發現NEDDylation修飾HBV X蛋白(HBx)后可阻止HBx通過泛素化途徑降解[12],從而促進cccDNA轉錄激活[13]。NAE抑制劑MLN4924抑制HBx NEDDy lation,HBx降解增多,其促進cccDNA轉錄作用減弱,從而導致編碼核心蛋白的pgRNA減少。
MLN4 924劑量依賴地減弱pgRNA包裝和HBV DNA復制,而且兩者下降程度相近。HBV DNA復制是 pgRNA包裝入核衣殼的后續步驟,因此MLN4924通過抑制pgRNA包裝而減弱HBV DNA復制。pgRNA包裝是個復雜的過程。在宿主因子如HSP90等的幫助下,HBV聚合酶識別并結合pgRNA,再招募核心蛋白二聚體組裝成核衣殼,如果NEDDylation修飾上述病毒或宿主蛋白,或修飾其他未知宿主因子而調節pgRNA包裝,則像MLN4924這樣的NEDDylation抑制劑將抑制pgRNA包裝。因此,本研究還將檢驗 HBV聚合酶、核心蛋白和HSP90是否為NEDDylation的底物,這將有助于進一步了解HBV復制機理。
利益沖突 無
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
