秈米中嘉早17儲藏過程中蛋白質氧化程度及結構的變化
2018-11-13 07:28:46吳曉娟
中國糧油學報 2018年10期
關鍵詞:結構
吳 偉 吳曉娟
(中南林業科技大學食品科學與工程學院;稻谷及副產物深加工國家工程實驗室,長沙 410004)
稻谷是我國65%以上人口的口糧,稻谷的儲藏和生產直接關系到國家糧食供應體系的安全。早秈稻比中晚秈稻具有更好的耐儲藏性,是南方地區儲備糧輪換的主要品種。目前,南方地區的稻米產量已經達到中國稻米產量的50%以上[1]。然而作為“第五、七儲糧生態區”的廣大南方地區,夏季氣溫通常高達37℃以上、相對濕度高達85%以上,儲藏過程中稻米極易陳化變質[2]。
稻米在陳化過程中稻谷的物理、化學和生理性質發生一系列變化,導致稻米食用品質和使用品質下降,最終影響其在市場上的正常流通[3-4]。長期以來,稻米儲藏過程中淀粉和脂質含量和結構的變化被視為陳化導致稻米食用品質下降的重要原因,近年來相關研究表明稻米儲藏過程中大米蛋白結構和含量的變化也是導致稻米陳化的主要因素之一[3,5]。已有研究表明,隨著儲藏時間的延長,稻米中的米谷蛋白和醇溶蛋白含量變化不大,清蛋白和球蛋白含量顯著下降[6-7],但關于儲藏過程中大米蛋白結構變化的研究報道較少。此外,中國稻米品種繁多,已有研究報道不同品種稻米的蛋白質結構差異較大,崔莎莎[8]研究低水解度大米蛋白的結構時,僅發現β-折疊、無規卷曲和β-轉角結構,而缺少α-螺旋結構;易翠平等[9-10]研究堿處理和酸法脫酰胺對大米蛋白結構的影響時,僅發現β-折疊、α-螺旋和β-轉角結構,而缺少無規卷曲結構。中嘉早17是目前我國年推廣面積最大的秈稻品種[11]。因此,本研究以新收獲秈米中嘉早17為原料,采用溫度37℃、相對濕度85%進行加速陳化儲藏,研究儲藏過程中大米蛋白氧化程度及結構的變化,以期為深入探討秈米的陳化機理提供參考。
1 材料與方法
1.1 材料與試劑
新收獲秈米中嘉早17(國標一級):湖南糧食集團長沙儲備分公司;5,5'-二硫代二硝基苯甲酸鹽(DTNB)、1-苯氨基萘 -8-磺酸(ANS):美國Sigma-Aldrich公司;低分子量標準蛋白:上海生化試劑公司;甘氨酸(Gly)、三(羥甲基)氨基甲烷(Tris)、十二烷基硫酸鈉(SDS)等試劑均為分析純:國藥集團化學試劑有限公司。
1.2 儀器與設備
RXZ-128A型人工氣候箱:寧波市科技園區新江南儀器有限公司;Sorvall LYNX6000型高速冷凍離心機:美國Thermo Fisher公司;LGJ-18型冷凍干燥機:北京四環科學有限公司;722s型可見光分光光度計:上海精密科學儀器有限公司;F7000熒光分光光度計:日本日立公司;SE260型電泳儀:美國GE公司;Nano ZS型納米粒度分析儀:英國Malvern公司;IRTracer-100型傅里葉變換紅外光譜儀:日本島津公司。
1.3 方法
1.3.1 秈米加速陳化儲藏
參考王凡等[1]的方法加速陳化儲藏,取適量中嘉早17大米(含水量13.48%)置于15 cm×20 cm大小的托盤中,放入人工氣候箱中加速陳化儲藏,儲藏條件為溫度37℃、相對濕度85%,每周取樣。
1.3.2 大米蛋白制備
參考吳偉等[12]的方法制備大米蛋白,將儲藏不同時間的大米磨粉過80目篩,按料液比1∶7(m/V)加入去離子水中,再用2 mol/L NaOH溶液調節pH值至9.0,在40 ℃、120 r/min下攪拌反應4 h,然后將懸浮液在4℃、8 000 r/min下離心20 min,取上清液用2 mol/L鹽酸溶液調節pH至4.0,靜置20 min,4℃、8 000 r/min離心15 min得到蛋白沉淀,用去離子水洗滌3次后,再將蛋白沉淀分散于4倍體積的去離子水中,用2 mol/L NaOH溶液調節pH 至7.0,透析72 h后冷凍干燥得到純度為82%的大米蛋白。
1.3.3 大米蛋白羰基含量測定
參考Huang等[13]的方法,采用 2,4-二硝基苯肼比色法測定大米蛋白的羰基含量。稱取100 mg左右的大米蛋白分散于5 mL 0.05 mol/L pH 8.0的Tris-HCl緩沖液中,磁力攪拌2 h后在4℃、8 000 r/min下離心30 min,用考馬斯亮藍比色法測定上清液的蛋白濃度,然后將上清液稀釋為蛋白質量濃度2.86 ~4.29 mg/mL 的溶液。用移液管量取0.35 mL該溶液置于離心管中,加入1 mL含10 mmol/L 2,4-二硝基苯肼的2 mol/L鹽酸溶液混合均勻,以不加2,4-二硝基苯肼為空白對照。將混合液在20℃下水浴2 h后,分別加入0.45 mL 40%的三氯乙酸振蕩搖勻,靜置20 min后在4℃、8 000 r/min下離心20 min,棄去上清液,使沉淀懸浮于1.5 mL乙醇-乙酸乙酯(1∶1,V/V)混合液中,在 4 ℃、8 000 r/min 下離心20 min,如此重復洗滌沉淀3次。最后將蛋白沉淀懸浮于1 mL 0.1 mol/L含6 mol/L鹽酸胍的pH 8.0的Tris-HCl緩沖液中,在37℃下水浴20 min。以空白為對照在367 nm波長校正,以22 000 L/(mol·cm)消光系數計算每毫克蛋白質中羰基衍生物的物質的量。
1.3.4 大米蛋白游離巰基及二硫鍵含量測定
參考 Beveridge等[14]的方法,采用 Ellman's試劑比色法測定大米蛋白的巰基和二硫鍵含量。稱取120 mg大米蛋白置于燒杯中,加入20 mL 8 mol/L尿素Tris-Gly溶液(pH為8.0),室溫磁力攪拌2 h后在4℃、8 000 r/min下離心30 min,用考馬斯亮藍比色法測定上清液的蛋白濃度。取4 mL上清液加入160μL Ellman's試劑(4 mg DNTB溶于1 mL pH 8.0的Tris-Gly緩沖液),以不加DNTB為對照,在412 nm波長下測定吸光度,以13 600 L/(mol·cm)消光系數計算游離巰基含量。另取4 mL上清液加入0.2% β-巰基乙醇處理2 h后,再加入8 mL 12%的三氯乙酸沉淀蛋白質1 h,在10 000 r/min下離心10 min,用三氯乙酸溶液重復洗滌沉淀3次后,將沉淀溶于6 mL Tris-Gly緩沖液中,以不加 DNTB為對照,在412 nm波長下測定吸光度,以13 600 L/(mol·cm)消光系數計算總巰基含量。總巰基與游離巰基差值的1/2即為二硫鍵含量。
1.3.5 大米蛋白表面疏水性測定
參考Huang等[13]方法,采用ANS熒光探針法測定大米蛋白的表面疏水性。稱取150 mg大米蛋白分散于10 mL 0.05 mol/L pH 8.0 的 Tris-HCl緩沖液中,磁力攪拌2 h后在4℃、8 000 r/min下離心30 min,用考馬斯亮藍比色法測定上清液的蛋白濃度,然后將上清液稀釋為蛋白質量濃度0.005、0.01、0.05、0.20、0.40、0.50 mg/mL 的溶液。取不同蛋白濃度溶液4 mL,分別加入50μL 8 mmol/L ANS溶液,測定熒光強度,激發波長390 nm,發射波長470 nm。以熒光強度對蛋白質濃度作圖,外推至蛋白質濃度為0,曲線初始階段的斜率即為大米蛋白的表面疏水性指數。
1.3.6 大米蛋白傅里葉變換紅外光譜的測定
參考Liu等[15]的方法,稱取1~2 mg大米蛋白和200 mg KBr混合,研磨均勻后制作透明薄片,采用傅里葉變換紅外光譜儀掃描,掃描條件:分辨率4 cm-1,掃描范圍 400 ~4 000 cm-1,每個樣品掃描32次。通過傅里葉紅外解卷積技術提高分辨率,以Peak fit 4.12軟件對圖譜進行分峰處理。
1.3.7 大米蛋白凝膠電泳
參考Huang等[13]的不連續電泳方法。分離膠質量分數12.5%,濃縮膠質量分數4%,電極緩沖液含0.05 mol/L Tris、0.384 mol/L 甘氨酸、0.1%SDS(pH 8.3),樣品溶解液含2%SDS、5% β - 巰基乙醇、10%甘油、0.02%溴酚藍、0.01 mol/L pH 8.0 Tris -HCl緩沖液。電泳樣品質量濃度1.5 mg/mL,上樣量10μL,開始電泳時電流為10 mA,待樣品進入分離膠后電流為25 mA。
1.3.8 大米蛋白粒徑測定
將大米蛋白分散于0.05 mol/L pH 8.0的Tris-HCl緩沖液中,制備1 mg/mL蛋白液,采用納米粒度分析儀測定大米蛋白粒徑分布。
1.3.9 數據處理
所有實驗平行測定3次。數據采用Microsoft excel 2003和Origin 7.5進行處理,結果用“均值±標準偏差”表示。指標比較采用最小顯著差異法,取95%置信度(P <0.05)。
2 結果與分析
2.1 大米蛋白氧化程度的變化
羰基含量的變化是目前最常用的衡量蛋白質氧化程度的指標。如表1所示,秈米中嘉早17加速陳化過程中,大米蛋白羰基含量呈逐漸上升趨勢。第1~6周增加比較緩慢,由初始4.02 nmol/mg上升到4.72 nmol/mg;第 6 ~12 周則顯著增加,由 4.72 nmol/mg上升到8.34 nmol/mg。這可能是由于加速陳化過程中稻米中存在的脂肪氧化酶的同工酶,脂肪氧化酶同工酶可催化亞油酸等脂質產生自由基和羰基化合物等活性氧化產物[16]。肉類和大豆蛋白質氧化的大量研究表明,脂質自由基可通過奪氫、加氧、偶合以及裂解等反應,使得蛋白質主肽鏈斷裂、側鏈基團氧化以及形成共價交聯物;脂質活性氧化產物可與多肽鏈的側鏈基團發生反應,造成多肽鏈交聯[17-18]。因而,秈米中嘉早17加速陳化過程中的蛋白質氧化程度變化趨勢與油脂氧化自催化反應類似,遵循開始緩慢而逐漸加快的變化規律。
巰基是最容易被自由基攻擊的蛋白質氨基酸殘基側鏈基團[19]。依據氧化環境和氧化強度的改變,蛋白質巰基可形成二硫鍵和次磺酸等可逆氧化狀態,也可形成亞磺酸和磺酸等不可逆氧化狀態[20]。如表1所示,秈米中嘉早17加速陳化過程中,大米蛋白游離巰基含量呈逐漸下降趨勢,而二硫鍵含量無顯著變化。這表明,大米儲藏過程中,其蛋白質游離巰基主要發生的是不可逆氧化反應,生成了更多的非二硫鍵共價化合物。
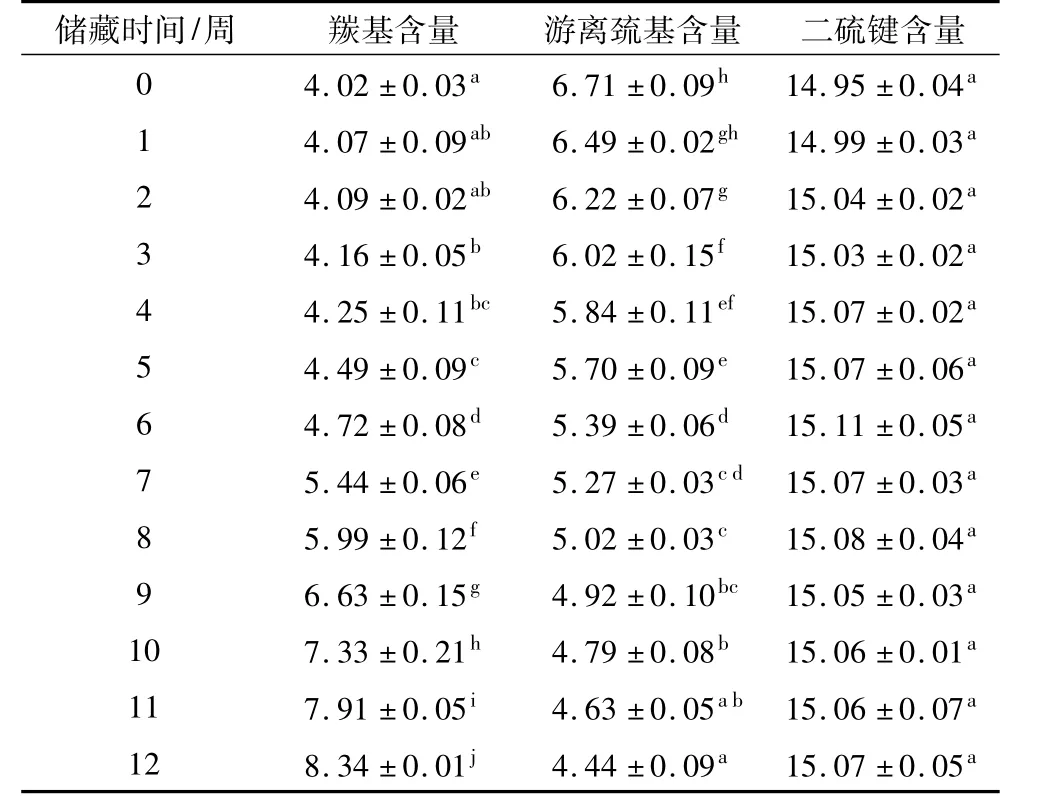
表1 秈米中嘉早17儲藏過程中大米蛋白羰基、游離巰基和二硫鍵含量/nmol/mg
2.2 大米蛋白表面疏水性的變化
表面疏水性反映蛋白質中疏水性氨基酸殘基的相對暴露程度,也可以用它來衡量蛋白質的結構變化。如圖1所示,第1~10周隨著儲藏時間延長,中嘉早17大米蛋白的表面疏水性逐漸升高;第10~11周,其表面疏水性無顯著變化;第12周,其表面疏水性反而略微下降。前期儲藏過程中表面疏水性升高可能是由于在脂質氧化形成的自由基氧化體系中,大米蛋白發生了去折疊反應,使埋藏在蛋白質天然構象內部的脂肪族與芳香族氨基酸側鏈基團等疏水性氨基酸殘基暴露。儲藏后期,大米蛋白表面疏水性不升反降,可能是由于之前暴露的疏水性基團進一步氧化轉變為親水基團,或者是暴露的疏水基團通過疏水相互作用形成了聚集體[21]。
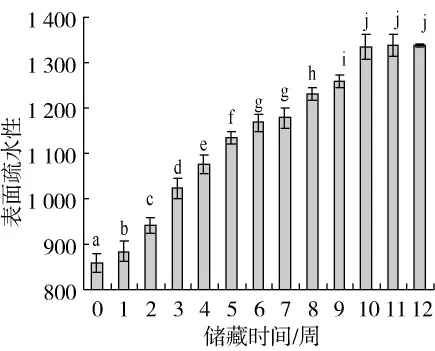
圖1 秈米中嘉早17儲藏過程中大米蛋白表面疏水性的變化
2.3 大米蛋白二級結構的變化
參考Liu等[15]的方法對大米蛋白的去卷積酰胺Ⅰ帶譜峰(如圖2所示)進行鑒定,β-折疊結構:1 618、1 624、1 630、1 636 和1 684 cm-1;無規卷曲結構:1 645 cm-1;α-螺旋結構:1 655和1 660 cm-1;β-轉角結構:1 670和 1 676 cm-1。采用 Peak fit 4.12對酰胺Ⅰ帶圖譜進行分峰擬合處理,得到大米蛋白的二級結構組成(如圖3所示)。隨著中嘉早17儲藏時間的延長,大米蛋白β-折疊結構相對含量顯著降低,無規卷曲和β-轉角結構相對含量逐漸增加,α-螺旋結構相對含量無顯著變化。有序的β-折疊逐漸向無序的無規卷曲和β-轉角結構轉變,這表明秈米中嘉早17加速陳化儲藏導致大米蛋白二級結構穩定性降低。
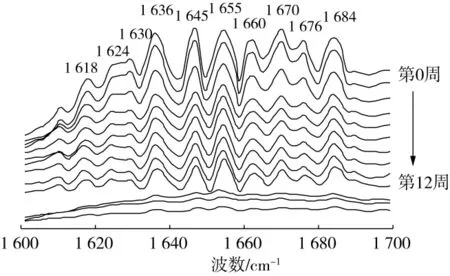
圖2 秈米中嘉早17儲藏過程中大米蛋白傅里葉變換紅外光譜酰胺Ⅰ帶
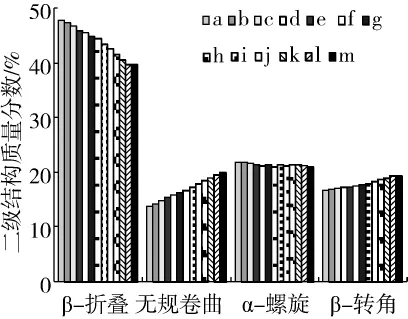
圖3 秈米中嘉早17儲藏過程中大米蛋白二級結構組成的變化
2.4 大米蛋白亞基結構的變化
大米蛋白的主要成分為谷蛋白和醇溶蛋白,其含量占大米蛋白總量的80%以上。醇溶蛋白亞基由單肽鏈通過分子內二硫鍵連接而成,其相對分子質量在7~12.6 ku之間[22]。相對分子質量21 ~22 ku為谷蛋白堿性亞基,34~37 ku為谷蛋白酸性亞基,51 ku為谷蛋白低分子聚肽[23]。如圖4所示,隨著儲藏時間的延長,大米蛋白凝膠電泳圖譜沒有出現新的譜帶,但儲藏10周以后,谷蛋白堿性亞基、谷蛋白酸性亞基帶、谷蛋白低分子聚肽的顏色明顯變淺,而分離膠頂部的顏色逐漸加深。這可能是由于大米蛋白中巰基含量較高,巰基在脂質自由基和脂質過氧化產物的氧化下發生分子內或分子間交聯,使谷蛋白形成分子質量超過100 ku的聚集體。儲藏12周時大米蛋白氧化聚集體的大量產生,可能正是導致其表面疏水性不升反降的重要原因。而醇溶蛋白亞基帶的顏色則先變淺后變深,這可能是由于較低濃度的脂質過氧化產物會誘導醇溶蛋白亞基聚集,但較高濃度的脂質過氧化產物也可能導致蛋白質組分部分降解[24]。
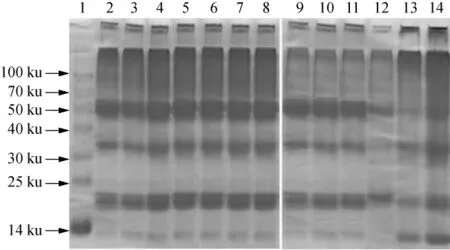
圖4 秈米中嘉早17儲藏過程中大米蛋白亞基結構的變化
2.5 大米蛋白粒徑分布的變化
中嘉早17儲藏過程中大米蛋白粒徑分布的變化如圖5所示,隨著儲藏時間的延長,大米蛋白粒徑逐漸增加,這表明儲藏過程中大米蛋白形成了可溶性聚集體。吳偉等[25]在儲藏時間對米糠蛋白結構影響的研究中,也發現隨著儲藏時間的延長米糠球蛋白形成了水溶性聚集體。如前所述,這可能是由于秈米中特有的脂肪氧化酶同工酶催化殘余脂質形成了活性脂質氧化產物,進一步誘導大米蛋白氧化形成了聚集體,從而使得粒徑不斷增加。
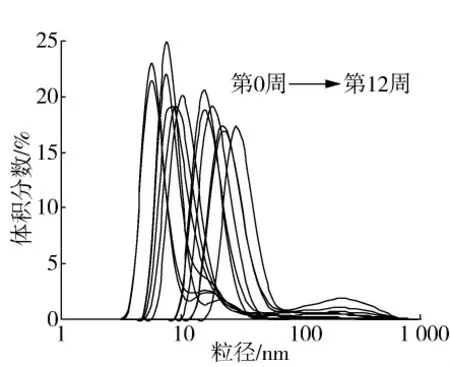
圖5 秈米中嘉早17儲藏過程中大米蛋白粒徑分布的變化
3 結論
秈米中嘉早17經過為期12周的加速陳化儲藏,制備的大米蛋白發生了一定程度的氧化和結構變化。儲藏過程中,大米蛋白氧化程度的變化趨勢與油脂氧化自催化反應類似,遵循開始緩慢而逐漸加快的變化規律,主要表現為蛋白質羰基含量不斷增加,游離巰基含量不斷減少,二硫鍵含量無顯著變化。游離巰基主要發生的是不可逆氧化反應,生成了更多的非二硫鍵共價化合物。同時,大米蛋白表面疏水性則呈現先升高后緩慢降低的趨勢。傅里葉變換紅外光譜分析表明,大米蛋白二級結構中有序的β-折疊逐漸向無序的無規卷曲和β-轉角結構轉變,蛋白質氧化導致大米蛋白二級結構穩定性降低。凝膠電泳和粒徑分析結果均表明,秈米中嘉早17儲藏過程中,蛋白質氧化導致大米蛋白形成了氧化聚集體。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
